
Abstract
Vesicle-associated membrane proteins (VAMPs) are important to the trafficking of vesicles between membrane-bound intracytoplasmic organelles, in the facilitation of neurosecretion, and in constitutive and regulated secretion in non-neuronal cells. We used a pre-embedding ultrastructural immunonanogold method to localize VAMPs to subcellular sites in human cells of five lineages known to have cytoplasmic vesicles that may function in vesicular transport. We found VAMPs localized to caveolae in pericytes, vascular smooth muscle cells, and endothelial cells of venules, to the vesiculo–vacuolar organelle, recently defined in venular endothelial cells, to the vesicle-rich intergranular cytoplasm and secretory granule membranes of neutrophils, and to perigranular cytoplasmic secretory vesicles and secretory granule membranes in eosinophils. These specific localizations in five human vascular and granulocyte lineages support the notion that VAMPs have vesicle-associated functions in these cells.
Keywords
H
VAMP-2 has been reported in a large number of different cell types, including parotid acinar cells (Fujita–Yoshigaki et al. 1996, 1999), kidney collecting duct cells (Nielsen et al. 1995a,b; Mandon et al. 1996; Rothman and Wieland 1996), adipocytes (Cain et al. 1992; Martin et al. 1996; Tamori et al. 1998), neutrophils (Brumell et al. 1995; Mott et al. 1999), mast cells (Guo et al. 1998), pancreatic islet cells (Braun et al. 1994; Regazzi et al. 1995), kidney glomerular cells (Rossetto et al. 1996; Rothman and Wieland 1996), brain (Baumert et al. 1989; Elferink et al. 1989; Trimble et al. 1990; Chilcote et al. 1995), skeletal muscle (Volchuk et al. 1994), pancreatic acinar cells (Braun et al. 1994; Gaisano et al. 1994), adrenal gland (Chilcote et al. 1995; Desnos et al. 1995; Grote et al. 1995; Grote and Kelly 1996; Rothman and Wieland 1996), liver hepatocytes (Rothman and Wieland 1996), thyroid gland (Rothman and Wieland 1996), heart (Rothman and Wieland 1996), smooth muscle cells of aorta (Rossetto et al. 1996), gastric enterochromaffin cells (Höhne–Zell et al. 1997), gastric parietal cells (Peng et al. 1997), adrenal chromaffin cells (Höhne–Zell et al. 1994), and lung endothelial cells (Schnitzer et al. 1995b). Subcellular localization of VAMP-2 has been determined primarily by biochemical measurements performed on fractions of cell extracts and by light microscopic immunohistochemical methods. Ultrastructural immunocytochemical methods have also been applied to localize VAMP-2 to subcellular fractions (Nielsen et al. 1995b; Martin et al. 1996). For example, intracellular vesicles isolated from adipocytes revealed co-localization of the glucose transporter GLUT-4 and VAMP-2 by immunoelectron microscopy (Martin et al. 1996). Similarly, isolated synaptic vesicle preparations labeled for VAMP-2, and isolated vesicles from kidney collecting duct cells co-labeled for VAMP-2 and aquaporin-2 water channels (Nielsen et al. 1995b). At least one in situ ultrastructural immunocytochemical study to localize VAMP-2 has appeared (Brumell et al. 1995). In this study of human neutrophils, immunoelectron microscopy of frozen thin sections revealed VAMP-2 associated with the small tertiary gelatinase-containing granule population (but not with primary or secondary granules), as well as with small secretory vesicles in the cytoplasm (Brumell et al. 1995).
Definition of the tissue and cellular distribution and subcellular organellar localization of specific proteins or classes of proteins is important for understanding their function(s). To this end, we applied a pre-embedding immunonanogold protocol for ultrastructural detection of VAMPs in human skin tissues and granulocyte preparations. The samples included five cell types known to contain small cytoplasmic vesicles which, in many instances, are believed to have a role in intracytoplasmic trafficking (Palade 1953; Simionescu et all. 1975; Dvorak et al. 1991, 1992; Anderson 1993a,b; Chang et al. 1994; Brumell et al. 1995; Predescu et al. 1997; Feng et al. 1999). In several cases, either biochemical or morphological studies have implicated VAMP-2 as the v-SNARE in these cells (Brumell et al. 1995; Schnitzer et al. 1995a; Rothman and Wieland 1996; Mott et al. 1999). Our immunonanogold studies localized VAMPs to caveolae in pericytes, endothelial cells, and vascular smooth muscle cells, to vesiculo-vacuolar organelles (Kohn et al. 1992; Dvorak et al. 1996; Feng et al. 1996, 1999) in venular endothelial cells, and to secretory vesicle-rich cytoplasm in eosinophils and neutrophils.
Materials and Methods
Tissue and Cells
Two foreskins were immersed in freshly prepared 4% paraformaldehyde in a 0.02 M PBS, pH 7.4, immediately after resection. Tissues were fixed for 4 hr at room temperature (RT), immersed in 30% sucrose in the 0.02 M PBS, pH 7.4, overnight at 4C, embedded in OCT compound (Miles; Elkhart, IN), and stored in −176C liquid nitrogen for subsequent use. Frozen 10-μm sections were cut with a standard cryostat and collected on pre-cleaned glass slides which were air-dried for 20 min before staining.
Citrate anticoagulated blood from normal donors was depleted of erythrocytes and centrifuged on Ficoll-Paque gradients (Amersham Pharmacia; Uppsala, Sweden) as described (Bandeira–Melo et al. 2000). Granulocyte mixed suspensions of about 8% eosinophils plus 92% neutrophils were then fixed in 4% paraformaldehyde in 0.02 M PBS, pH 7.4, for 1 hr at RT, washed in 0.02 M PBS, pH 7.4, and transferred to microtubes and centrifuged at 1500 × g for 1 min. They were then resuspended in molten 2% agar and quickly recentrifuged. The resultant agar pellets containing the cells were washed in PBS before immersion in 30% sucrose in 0.02 M PBS, pH 7.4, overnight. Ten μm frozen sections of OCT compound-embedded agar pellets were prepared as for foreskin.
Antibody to VAMP-2
Rabbit VAMP-2 polyclonal antibody (StressGen Biotechnologies; Victoria, BC, Canada) was produced using a 21-residue synthetic peptide based on residues 36–56 of rat VAMP-2 (Pevsner et al. 1994) coupled to KLH. This peptide sequence is conserved among VAMP family members. For example, VAMP-1 and VAMP-3 are 95% homologous to VAMP-2 within the amino acid region on which the synthetic peptide is based. By using an antibody directed at a conserved region of VAMP family isoforms, a wider spectrum of VAMPs can be searched for in ultrastructural samples than by labeling with an antibody that recognizes a single VAMP isoform. Thus, positive label implicates the VAMP family in the function of labeled subcellular organelles. One of us (RF) has previously used this same antibody in immunoblotting of brain and platelet lysates and has found that the antibody recognizes a band of the appropriate molecular weight for the VAMP family and does not recognize other bands in these lysates (Flaumenhaft et al. 1999).
Immunonanogold-Silver Staining Protocol and Processing for Electron Microscopy
The following steps were performed at RT on cryostat sections mounted on glass slides: (a) one wash in 0.02 M PBS, pH 7.6, 5 min; (b) immersion in 50 mM glycine in 0.02 M PBS, pH 7.4, 10 min; (c) one wash in 0.02 M PBS, pH 7.4, 5 min; (d) immersion in 5% normal goat serum (NGS) (Vector Laboratories; Burlingame, CA), 20 min; (e) incubation in the primary antibody, an affinity-purified rabbit polyclonal antibody against VAMP-2 (StressGen Biotechnologies) (12.5 μg protein/ml, diluted 1:20, 1:100, or 1:140 in 0.02 M PBS), 60 min; (f) three washes in 0.02 M PBS, pH 7.4, 5 min each; (g) incubation in the secondary antibody (affinity-purified 1.4 nm nanogold-conjugated goat anti-rabbit Fab′ (Nanoprobes; Stony Brook, NY), 1:50 in 0.02 M PBS, pH 7.4, 30 min; (h) three washes in 0.02 M PBS, pH 7.4, 5 min each; (i) postfixation in 1% glutaraldehyde in 0.02 M PBS, pH 7.4, 5 min; (j) three washes in distilled water, 5 min each; (k) development with HQ silver enhancement solution (Nanoprobes) for 6–11 min in the darkroom; (l) two washes in distilled water, 2 min each; (m) fixation in 5% sodium thiosulfate, 1 minute; (n) two washes in distilled water, 2 min each; (o) postfixation in 1% osmium tetroxide in Sym-Collidine buffer, pH 7.4, 10 min, RT; (p) one wash in 0.05 M sodium maleate buffer, pH 5.2, 2 min; (q) staining with 2% uranyl acetate in 0.05 M sodium maleate buffer, pH 6.0, 5 min, RT; (r) one wash in distilled water, 2 min; (s) dehydration in graded ethanols and infiltration with a propylene oxide–eponate (Eponate 12 Resin; Ted Pella, Redding, CA) sequence; (t) embedment by inverting eponate-filled plastic capsules over the slide-attached tissue sections; (u) polymerization at 60C for 16 hr; (v) separation of eponate blocks from glass slides by brief immersion in liquid nitrogen; (w) cutting of thin sections with a diamond knife on an ultratome (Reichert; Vienna, Austria) and collection of sections on uncoated 200-mesh copper grids (Ted Pella); (x) viewing of unstained grids with a transmission electron microscope (CM 10; Philips, Eindhoven, The Netherlands).
Controls for Immunostaining
Four controls were performed on foreskin samples to ensure the specificity of immunostaining: (a) primary antibody was replaced by an irrelevant rabbit IgG; (b) omission of specific primary antibody; (c) omission of the secondary antibody; (d) omission of the HQ silver enhancement solution. Controls a and b were performed on leukocyte preparations.
Results
Human Foreskin
Circumcision samples from newborn infants were prepared for pre-embedding immunonanogold staining. Cells associated with dermal venules in these samples were positive for VAMP. The most consistently labeled cells were pericytes and vascular smooth muscle cells; endothelial cells also were intermittently labeled.
Pericytes. Venular pericytes (Figure 1) beneath venular endothelium were elongated bipolar cells that sometimes formed contacts with overlying endothelial cells. Their cytoplasmic constituents included actin filaments, intermediate filaments, and focal rows of single caveolae attached primarily to their abluminal plasma membranes. In contrast, the luminally oriented plasma membrane was generally devoid of caveolae in most pericytes (Figures 1A, 1C, 1D, and 1E). Basal lamina was associated primarily with the abluminal pericyte surface. Mitochondria, strands of rough endoplasmic reticulum, and rare coated vesicles were also present. Pericyte caveolae were extensively labeled with antibody to VAMP. Label was associated primarily with the cytoplasmic face of abluminal plasma membrane-attached caveolae (Figures 1A–1C). Clusters of silver-enhanced nanogold particles sometimes obscured underlying caveolae. Rarely, VAMP was also localized to pericyte abluminal plasma membranes near caveolar attachments (Figure 1C). Coated vesicles were not labeled for VAMP. Controls for the immunonanogold method (omission of the primary antibody or substitution of an irrelevant rabbit IgG for the primary antibody) revealed no label (Figures 1D and 1E).
Endothelial Cells. Venular endothelial cells (Figures 1A, 1D, and 2) contained abundant caveolae, associated with both luminal and abluminal plasma membranes. In addition, focal clusters of vesicles and vacuoles, vesiculo-vacuolar organelles (VVOs) (Kohn et al. 1992; Dvorak et al. 1996; Feng et al. 1996, 1999), spanned the endothelial cells (Figure 2); rarely, coated pits and vesicles were noted. Of these structures, a small number (~5%) of VVOs were labeled for VAMP (Figure 2B). As was the case for pericyte caveolae, the VAMP labeling of VVO vesicles and vacuoles was primarily associated with their cytoplasmic faces. One vessel that showed morphological criteria of injury displayed heavily labeled endothelial cells. Controls, omission of the primary antibody or substitution for the primary antibody with irrelevant rabbit IgG (Figures 1D and 2C), were negative.
Smooth Muscle Cells. Processes extended from the surfaces of smooth muscle cells (Figure 3) to form characteristic “ball and socket-like” contacts with adjacent smooth muscle cells, thus creating a distinctive smooth muscle cell syncytium. Smooth muscle cells were encased by basal lamina and displayed many cytoplasmic thick and thin filaments. These actin and myosin filaments were aggregated as fusiform dense bodies in cytoplasmic filaments, and subplasmalemmal, electron-dense attachment areas for these filaments were prominent. The blunt, rounded tips of cell processes were typically packed with caveolae, several mitochondria, and collections of lipid and glycogen (Figures 3A–D and 3F). Coated pits and vesicles were infrequently present in smooth muscle cells (Figure 3H). Nanogold particles labeling VAMP in smooth muscle cells were attached to caveolar clusters (Figures 3A, 3C, 3E, and 3F), which encompassed all surfaces of these cells, in contradistinction to pericytes, in which labeled caveolae were generally not clustered and were largely confined to the abluminal surface. Plasma membranes (Figure 3A, 3E and 3F), coated pits (Figure 3H), and coated vesicles were not labeled; caveolae were labeled on their cytoplasmic surfaces (Figure 3G). Specificity controls were negative (Figure 3B and 3D).
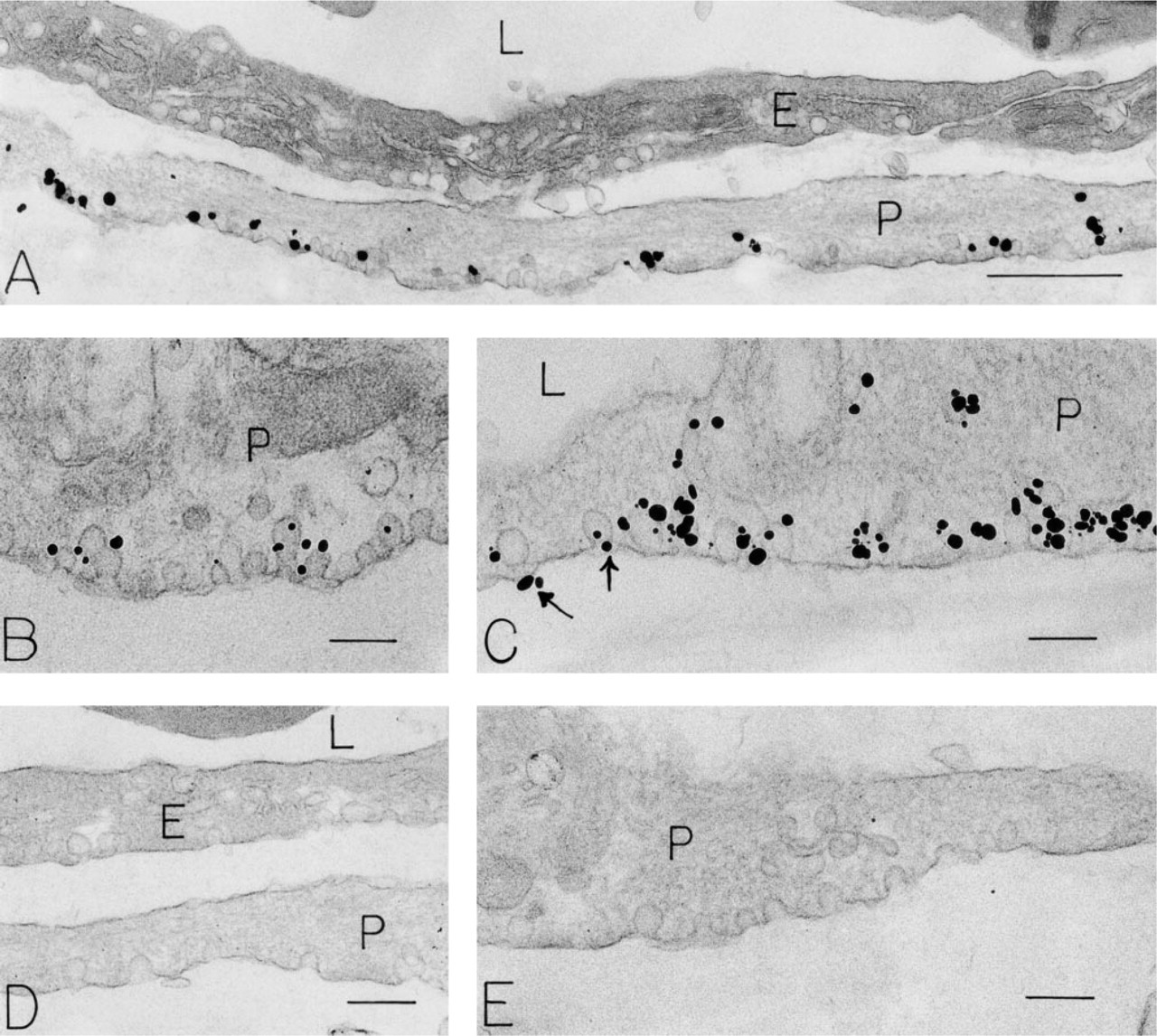
Immunonanogold localization of VAMP in pericytes of human foreskin venules. (
Human Peripheral Blood
Human peripheral blood samples from normal donors were partially purified to enrich granulocytes (Bandeira–Melo et al. 2000) before preparation of pre-embedding immunonanogold samples to detect VAMP. These preparations contained both eosinophils and neutrophils in sufficient numbers for the immunonanogold study.
Eosinophils. Eosinophils (Figure 4), granulocytes with polylobed nuclei, and typical crystalloid-containing cytoplasmic secretory granules displayed many cytoplasmic vesicles adjacent to granules and beneath the plasma membrane in a relatively granule-free zone. VAMP label was associated with the granule-poor subplasmalemmal, and inter-granule cytoplasm in eosinophils (Figure 4A and 4B). At higher magnifications and in cells showing evidence of secretion (Dvorak et al. 1991, 1992) from their granule matrix compartments, VAMP was attached to the cytoplasmic membrane surfaces of perigranular vesicles (Figure 4B). Some granule membrane (Figure 4A) and plasma membrane label for VAMP was also noted. Substitution of irrelevant rabbit IgG (Figure 4C) for the VAMP primary antibody yielded negative results; omission controls were also negative.
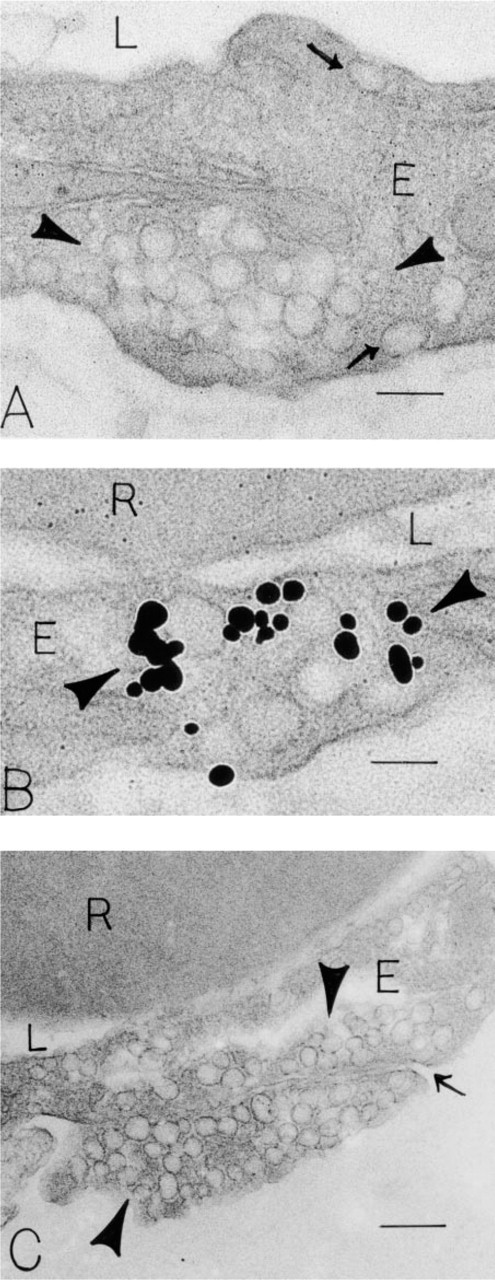
Immunonanogold localization of VAMP in endothelial (E) cells of human foreskin venules. (
Neutrophils. Neutrophils (Figure 5) with polylobed nuclei and a full component of secretory granules were labeled for VAMP (Figure 5A). Although some gold particles were associated with secretory granule membranes on their cytoplasmic surfaces (Figure 5A) and some were found at the interface between condensed and non-condensed nuclear chromatin, in many cells VAMP label was present in a granule-poor, circumferential cytoplasmic band just beneath the plasma membrane. Substitution of irrelevant rabbit IgG for the VAMP-specific antibody gave no labeling (Figure 5B).
Discussion
We used a pre-embedding immunonanogold ultrastructural method (Feng et al. 2000a) to define the subcellular distribution of a v-SNARE, VAMP, in three cell lineages (endothelial cell, pericyte, smooth muscle cell) in human skin venules and in two granulocyte lineages. In all five cell types, VAMP was associated with individual and/or clustered cytoplasmic vesicles and in endothelial cells with caveolae and the interconnected vesicles and vacuoles of VVOs (Kohn et al. 1992; Qu-Hong et al. 1995; Dvorak et al. 1996; Feng et al. 1996, 1997, 1999, 2000a,b; Vasile et al. 1999).
Caveolae, also called plasmalemmal vesicles, were originally described in capillary endothelial cells, which contain rows of these flask-shaped vesicles individually attached to endothelial cell luminal and abluminal surfaces (Palade 1953). Since this original description, caveolae have been described in many cell lineages, including pericytes and smooth muscle cells (Chang et al. 1994). Caveolae have been thought to have at least three functions that are not necessarily mutually exclusive. One such proposed function is that individual caveolae separate from their plasma membrane attachments and shuttle back and forth across capillary endothelial cells, discharging plasma “cargo” into the extracellular space (Predescu et al. 1994; Schnitzer et al. 1995a; Predescu et al. 1997; Stan et al. 1997). They are also believed to remain attached to the plasma membranes but to periodically open and close there, delivering extracellular molecules to the cell interior, a process that has been termed potocytosis (Anderson 1993a,b). In addition, ultrastructural studies with electron-dense tracers have shown that chains of two or three capillary endothelial cell caveolae may join together to form transcellular channels (Simionescu et al. 1975).
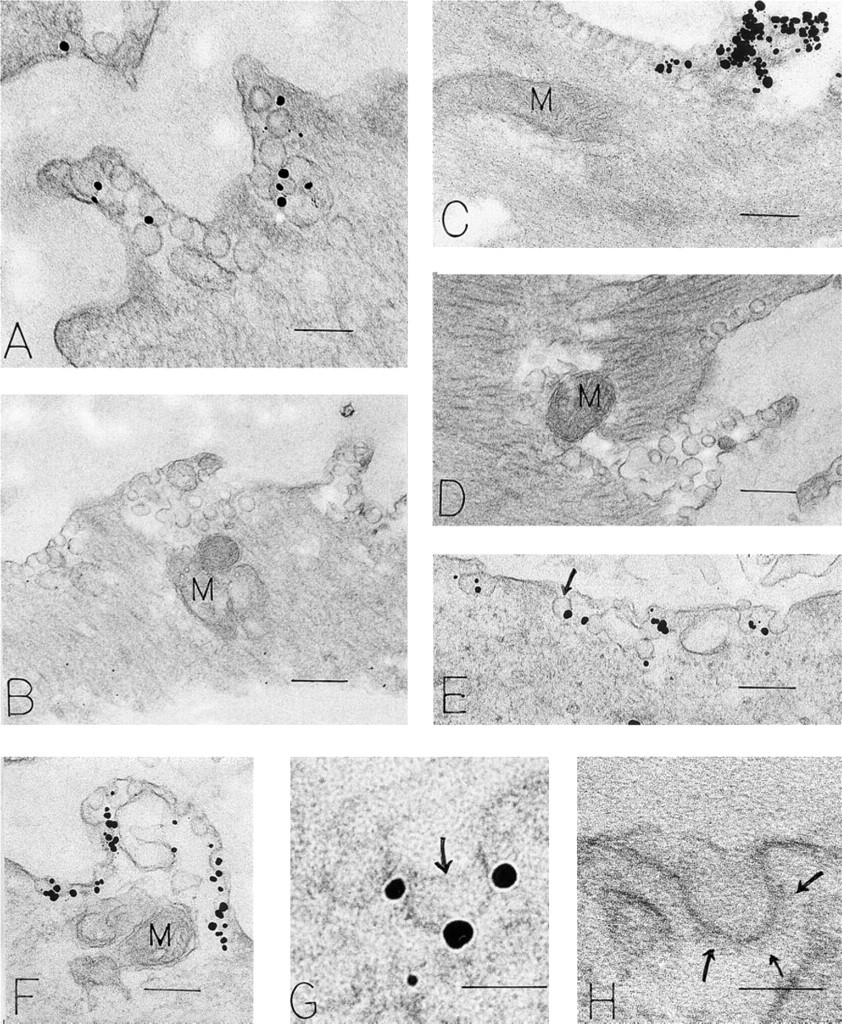
Immunonanogold localization of VAMP in vascular smooth muscle cells of human foreskin venules. (
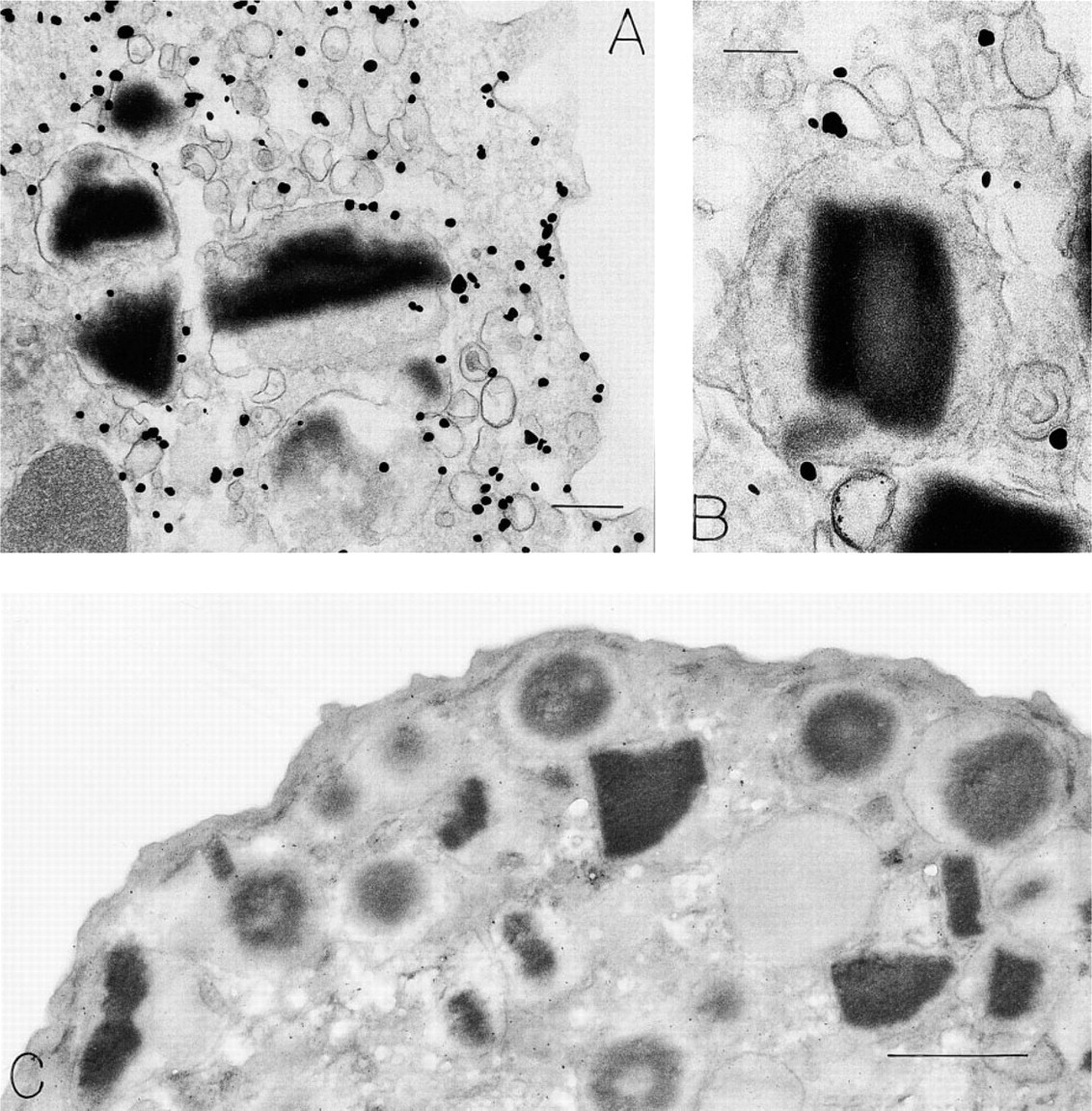
Immunonanogold localization of VAMP in human peripheral blood eosinophils. (
Purified rat lung endothelial cell caveolae fractions have been prepared (Schnitzer et al. 1995c,d), and biochemical analyses have shown that these caveolin-rich samples contain VAMP-2 (Schnitzer et al. 1995b). We found VAMP on the cytoplasmic surfaces of human foreskin venular endothelial cell caveolae attached to their plasma membranes with the immunonanogold label. However, many endothelial cell caveolae were not labeled. Whether this finding means that VAMP is truly absent from some of these structures in this site, or whether activation of endothelial cells is necessary to expose VAMP epitopes, was not determined. There is some biochemical evidence to support this idea (Flaumenhaft et al. 1999). For example, VAMP family proteins form very tight (e.g., heat- and SDS-resistant) associations with other SNARE proteins in vivo. The VAMP antibody that we used is directed against a portion of the VAMP molecule that directly associates with other SNARE proteins. The epitope in VAMP recognized by this antibody is therefore concealed when VAMP is in the bound state. Therefore, it is likely that we are labeling only free VAMP. It is possible that free VAMP represents a small portion of total VAMP in some of the specimens. The ratio of free to bound VAMP may depend on whether or not the tissue/cell has been activated in vivo. Other VAMPs (McMahon et al. 1993; Galli et al. 1994; Volchuk et al. 1994; Calakos and Scheller 1996; Martin et al. 1996; Rossetto et al. 1996; Linial 1997) may fulfill similar functions in the proposed transport, docking, and fusion of vesicles in endothelial cells. The antibody we used would most likely recognize other VAMPs, in addition to VAMP-2, that may function in endothelial cells. Therefore, precise specification of which VAMPs are involved requires further study. The strategy we used was designed to localize the VAMP family to subcellular organelles and did so with considerable precision in five cellular lineages.
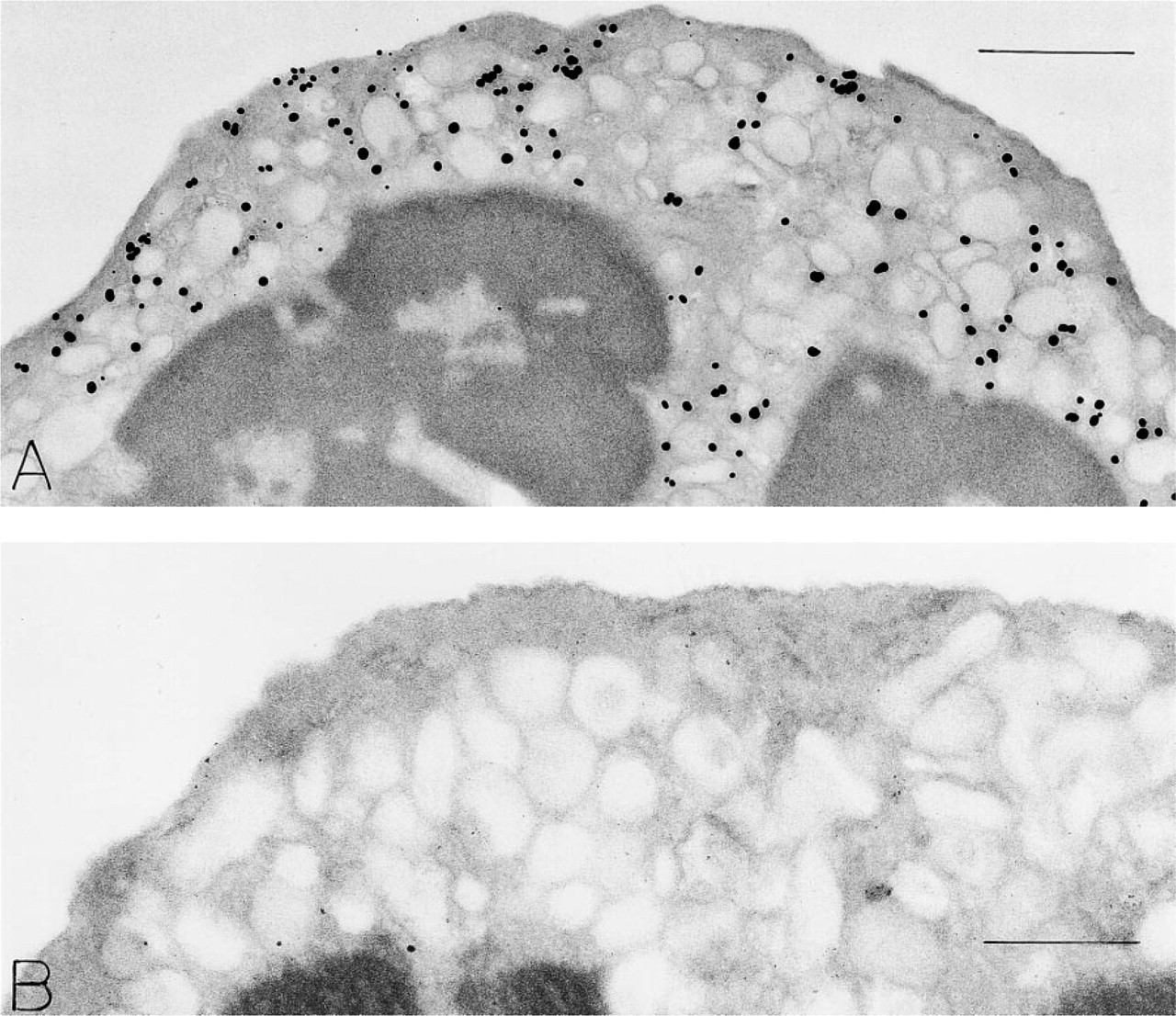
Immunonanogold localization of VAMP in human peripheral blood neutrophil (
Although capillary endothelial cell caveolae are believed by many to be capable of detachment from the plasma membrane and movement across cells (motile caveolae), cells also display single caveolae and clusters of surface-attached caveolae [capillary endothelial cells (Bundgaard et al. 1979; Frøkjaer–Jensen 1980, 1991; Bundgaard 1983; Frøkjaer–Jensen et al. 1988; Anderson 1993a), pericytes (Dvorak et al. 1976; Dvorak 1998a), smooth muscle cells (Dvorak et al. 1980; Chang et al. 1994) that are believed to be incapable of movement (sessile caveolae). Pericytes display caveolae that usually are single, attached to the abluminal surfaces, and believed not to be motile. Almost all of the purportedly sessile caveolae in venular pericytes were labeled with VAMP. This indicates that the methodology to demonstrate VAMP is effective in our preparations, that human pericyte caveolae have an abundance of this v-SNARE family, and, because these structures are single and sessile, that VAMP may subserve additional functions in novel membrane-mediated events in pericyte biology than those generally proposed in the SNARE hypothesis (Rothman and Warren 1994).
Smooth muscle cells also displayed surface-attached caveolae but, unlike pericytes, these were distributed on all surfaces (Dvorak et al. 1980). These caveolae were not found unattached to the cell–plasma membrane. Many were single flask-shaped structures, but clusters of interconnected caveolae were also present (Dvorak et al. 1980; Chang et al. 1994). These clusters were typically concentrated at smooth muscle cell contacts with adjacent smooth muscle cells, ball and socket-like connections that provide the basis for the anatomic and functional syncytium, unique to this cell lineage (Dvorak et al. 1980). VAMP-2 has been reported in aortic smooth muscle by in situ hybridization (Rossetto et al. 1996). The presence of VAMP in sessile clusters of caveolae at intercellular contact points suggests that VAMP may facilitate the opening/closing of these structures for intercellular communications, i.e., analogous to the proposed potocytosis of single sessile caveolae in capillary endothelial cells (Anderson 1993a,b).
Vesiculo–vacuolar organelles (VVOs) were first identified in tumor-associated microvessels and in the endothelia of normal venules of mice and guinea pigs (Kohn et al. 1992; Dvorak et al. 1996; Feng et al. 1996). They are bunches of grape-like clusters of vesicles and vacuoles attached to endothelial cell luminal, lateral, and basilar plasma membranes that interconnect with their individual units at multiple levels and with all three endothelial cell surfaces. Electron-dense tracers demonstrated that they provide a trans-endothelial pathway for the passage of macromolecules in response to vasoactive mediators such as vascular permeability factor/vascular endothelial growth factor (VPF/VEGF), histamine, or serotonin (Kohn et al. 1992; Dvorak et al. 1996; Feng et al. 1999). In addition, large series of ultrathin serial sections (often >100) and computer-assisted three-dimensional reconstructions showed that virtually 99% of these endothelial cell vesicles and vacuoles were connected by diaphagms identical to those of caveolae (Feng et al. 1996) (e.g., docked) and/or patent in conditions with increased permeability; <1% of such vesicles existed free in endothelial cytoplasm (Feng et al. 1996, 1999). We have proposed that these large sessile collections of vesicles and vacuoles are permeabilized when their interconnecting diaphragms are opened and that the entire structure is formed from fusion of unit-sized caveolae (Feng et al. 1999). In support of this proposal are a morphometric analysis of the volumes of individual vesicles and vacuoles, showing that vacuole growth expands in predictable multiples of the unit-sized caveolar volume (Feng et al. 1999), the presence of caveolin, a caveola-specific protein in VVOs (Vasile et al. 1999), and ultrastructural immunocytochemical studies that have localized VPF/VEGF (Qu-Hong et al. 1995) and one of its high-affinity receptors, VPF/VEGF receptor-2 (KDR) (Feng et al. 2000a), to VVOs. Morphologically, sessile VVOs in endothelial cells bear some similarities to collections of sessile caveolae in smooth muscle cells (Dvorak et al. 1980; Chang et al. 1994) and to tubulo–vesicular collections in gastric parietal cells (Peng et al. 1997). In this study, we demonstrated VAMP in vascular smooth muscle cell caveolar collections. Other investigators have shown that the gastric parietal cell cytoplasmic tubulo-vesicular organelle is associated with VAMP-2 (Peng et al. 1997).
Small secretory vesicles have been described in all three granulocyte lineages (Dvorak et al. 1991, 1992; Brumell et al. 1995; Dvorak 1998b, c). In neutrophils, these vesicular structures are involved with stimulated secretion and reside in a granule-free subplasma membrane cytoplasmic domain, as well as in the cytoplasm adjacent to granules (Brumell et al. 1995; Dvorak 1995). Whether they function to aid in release of granule products from the three granule populations or function independently of these granules is not known. However, we have localized VAMP primarily to these vesicle-rich cytoplasmic areas, in support of a previous report on VAMP-2 localization using a different ultrastructural approach (Brumell et al. 1995).
Eosinophils (and basophils) contain small cytoplasmic vesicles in similar locations that have a transport function, shuttling between their major granule populations and the plasma membrane in a secretory response termed piecemeal degranulation (Dvorak et al. 1991, 1992; Dvorak 1998b, c). These motile vesicles traverse the cytoplasm, fuse to granule and plasma membranes, and deliver granule products (eosinophil peroxidase, histamine, Charcot–Leyden crystal protein) to the extracellular milieu and extracellular electron-dense tracers to their secretory granules (Dvorak et al. 1991, 1992; Dvorak 1998b, c). In the present study we showed that human eosinophils have VAMP in these vesicle-rich perigranular and subplasma membrane cytoplasmic areas. Moreover, the identification of VAMP attached to granule membranes may indicate that at least some of the VAMP-containing perigranular vesicles are either fused with or derived from eosinophil granule membranes. Therefore, this v-SNARE (important in vesicular transport, docking, and fusion) may serve to function, as might be expected from the SNARE hypothesis (Rothman and Warren 1994), in piecemeal degranulation of eosinophils (Dvorak et al. 1991, 1992).
In summary, we have localized VAMP, an important family of v-SNAREs, to caveolae in pericytes, smooth muscle cells, and endothelial cells of human venules, to VVOs in venular endothelial cells, and to vesicle-rich cytoplasmic areas of human neutrophils and eosinophils. This distribution of label supports a role(s) for VAMP in vesicle transport mechanisms involved in macromolecular extravasation across venular endothelia, through sessile closed VVOs (which open), or in caveolae, which either do or do not (potocytosis) move. Single sessile caveolae in pericytes and clusters of sessile caveolae in vascular smooth muscle also labeled for VAMP, suggesting a role for this family of v-SNAREs in vesicular containers that may exert their function while remaining attached to the plasma membrane, much like VVOs. Circulating granulocytes also displayed label for VAMP in cytoplasmic vesicle-rich sites, lending support for a role(s) for this family of v-SNAREs in the vesicular transport of piecemeal degranulation, a secretory mode demonstrated in eosinophils (Dvorak et al. 1991, 1992) and basophils (Dvorak 1998b, c).
Footnotes
Acknowledgements
Supported by NIH grants AI-44066 and AI-33372 to A.M. Dvorak, HL-46563 and AI-20241 to Peter F. Weller, and HL-63250 to R. Flaumenhaft. R. Flaumenhaft is a Burroughs Wellcome Fund Career Awardee and is a participant in the Clinical Investigator Training Program, Beth Israel Deaconess Medical Center–Harvard/MIT Health Sciences and Technology, in collaboration with Pfizer, Inc.
We thank Peter K. Gardner for editorial assistance in the preparation of the manuscript, Patricia Fox for photographic assistance, and Tracey Sciuto for technical assistance.