
Abstract
Keywords
A
Despite its widespread tissue distribution, the physiological role of SSAO is far from understood. Together with its amine oxidase activity, SSAO might participate in the regulation of glucose disposal in several insulin-sensitive tissues, including cardiac and skeletal muscle and adipocytes, where it is associated with the glucose transporter GLUT4 (Enrique–Tarancón et al. 1998). A vascular adhesion protein, VAP-1, with identical sequence to the tissue-bound SSAO has recently been described (Smith et al. 1998; Bono et al. 1998). Because VAP-1 is involved in lymphocyte migration to lymphoid organs, a new physiological role for SSAO in cellular trafficking has been suggested (Smith et al. 1998; Jaakkola et al. 1999).
Changes in plasma SSAO activity have been related to several pathological situations. Plasma SSAO activity is reduced in patients suffering from burns (Lewinsohn 1984), fibrotic liver disease (Lewinsohn 1984), and diabetes mellitus (Boomsma et al. 1995). A decrease in tissue-bound SSAO activity accompanied by malignancy has also been reported in chemically induced rat breast tumors (Lizcano et al. 1991). Finally, the formaldehyde generated from methylamine metabolism by SSAO in vascular tissues has been proposed as a risk factor in stress-related angiopathy (Yu et al. 1997).
To better understand the physiological role of SSAO, the activity and cellular distribution of SSAO were determined in human tissues, including adrenal gland, duodenum, heart, kidney, lung, liver, pancreas, spleen, thyroid gland, and blood vessels, by enzymatic and immunohistochemical techniques using a specific antibody against tissue-bound SSAO (Castillo et al. 1998; Lizcano et al. 1998). Because MAO-A and -B may contribute to amine oxidation, their activities were also determined in these tissues. Because the role of SSAO has been related to a dual cellular distribution in some cells (Lizcano et al. 1991; Smith et al. 1998), its localization was also studied in the labeled cell types from each of these peripheral tissues. Finally, our results are compared with the MAO-A and -B distribution previously reported. (Saura et al. 1997; Rodríguez et al. 2000).
Materials and Methods
Human Tissues
Human tissues were generously provided by Dr. J. A. Bombí (Facultat de Medicina, Universitat de Barcelona, Spain) in accordance with local institutional ethical guidelines. Samples of adrenal gland, duodenum, heart, kidney, lung, liver, pancreas, spleen, thyroid gland, and vessels were obtained from nine routine autopsy cases (five men and four women). Tissues affected by the pathological state of the patient were discarded. The age range was 50–84 years (mean ± SEM, 72.2 ± 3.7) and the postmortem delay ranged from 4 to 22 hr (12.6 ± 1.9).
Immediately after autopsy, tissues were divided into two groups. One group was cut into 1–2-cm-thick blocks for the immunohistochemical studies and the other was stored in liquid nitrogen for enzymatic activity determination.
Enzyme Assays
Samples kept in liquid nitrogen were powdered with a mechanical hammer. They were then homogenized 1:10 (w/v) in 50 mM Na+, K+ phosphate buffer, pH 7.6, and filtered through a double gauze. Aliquots were taken and stored at –80C.
SSAO and MAO-B activities were determined radio-chemically (Fowler and Tipton 1981) with 100 μM [7-14C]-benzylamine (55 mCi/mmol; Amersham, Madrid, Spain) as substrate. MAO-A activity was measured towards 100 μM [14C]-5-HT (55 mCi/mmol; Amersham) as specific substrate. Because SSAO and MAO-B are both active towards benzylamine, MAO-B was previously inhibited with 1 mM deprenyl for SSAO determinations and SSAO was inhibited with 1 mM semicarbazide for MAO-B assays. After preincubation with the corresponding inhibitor for 30 min, each enzymatic reaction (150–220 μg of protein) was carried out at 37C for 1 hr in a shaking water bath in a final volume of 225 μl 50 mM potassium phosphate buffer, pH 7.6, stopped by 100 μl 2 M citric acid, and the radioactivity directly counted after a 4-ml addition of scintillation liquid (Lizcano et al. 1998). MAO-A activity was determined in similar conditions using an incubation time of 30 min.
Time-course assays were used to ensure that initial rates of the reaction were determined, and proportionality to enzyme concentration was established in each case. Protein was measured by the Bradford method (Bradford 1976), using γ-globulin as standard. All nonradioactive products were purchased from Sigma (St Louis, MO).
Immunoblotting Analysis
Human tissues homogenates or purified porcine kidney diamine oxidase (Sigma) were fractionated by electrophoresis on 8% SDS-polyacrylamide gels and transferred to nitrocellulose membranes. Blots were blocked for 1 hr at room temperature with 5% (w/v) non-fat dried milk in 20 mM Tris-HCl, pH 7.5, containing 150 mM NaCl and 0.05% Tween-20, then incubated with primary anti-SSAO antibody (1:500 dilution) overnight at 4C, followed by incubation for 1 hr with secondary antibody. Finally, the blots were developed with the avidin–biotin–peroxidase technique, following the manufacturer's instructions (Vector Laboratories; Bretton, UK). After color development, the membrane was washed with distilled water, air-dried, and photographed. Prestained molecular weight standards used were myosin (250 kD), bovine serum albumin (98 kD), glutamate dehydrogenase (64 kD), alcohol dehydrogenase (50 kD), carbonic anhydrase (36 kD), and myoglobin (30 kD) from Novex (San Diego, CA).
Immunohistochemical Assays
After autopsy, 1–2-cm tissue blocks were immediately fixed by immersion in paraformaldehyde in 0.1 M phosphate buffer, pH 7.4, at 4C. Different fixation times (2, 4, 7, or 24 hr) and fixative concentrations [0, 0.1, 1, 2, 4 % (w/v)] were tested. After fixation, tissues were immersed for 72 hr in 15% (w/v) sucrose in 0.1 M phosphate buffer, pH 7.4, for cryoprotection, frozen with dry ice, and stored at –30C. Sections of fresh frozen tissue were also postfixed with acetone or paraformaldehyde 4% (w/v) for 2 or 5 min.
Cryostat sections (12 μm) were cut from the fixed tissue blocks and mounted on gelatinized slides. The immunohistochemical avidin–biotin method was used. Endogenous peroxidase activity was inhibited by a 30-min preincubation in 30% (v/v) H2O2 in 10 mM Tris-buffered saline (TBS), pH 7.6, followed by a 15-min wash in TBS. Then sections were incubated in 20% (v/v) normal goat serum (NGS) in TBS for 20 min. A rabbit polyclonal anti-bovine SSAO antibody (Castillo et al. 1998; Lizcano et al. 1998) was used diluted in TBS-1% NGS as primary antibody for detection of SSAO protein. Incubation was performed overnight at 4C and antibody dilutions of 1:200, 1:300, 1:400, and 1:500 were tested. The antibody 9H7 (an anti-γ-gliadin, which has been typed as subclass IgG1) was diluted 1:400 and used to define the nonspecific staining. After washing, sections were incubated for 30 min at room temperature in biotinylated goat anti-rabbit IgG (1:300 in TBS–1% NGS), rinsed, and washed in TBS, and then incubated for 2 hr at room temperature in ExtraAvidin–horseradish peroxidase (1:250 in TBS–1% NGS). After washing, sections were developed for 15 min in a 50 mM Tris-HCl solution, pH 7.6, containing 0.03% (w/v) diaminobenzidine and 0.006% (v/v) H2O2. Some sections were counterstained with Mayer's hematoxylin for morphology. Secondary antibodies and reagents were purchased from Sigma.
Results
Specific Activities
Specific activities of MAO-A, MAO-B, and SSAO were detected in most of the tissues assayed, with marked differences (see Table 1). The highest specific activity of MAO-A was observed in liver, duodenum, and kidney (1003–1158 pmol·min−1·mg prot−1). Heart, lung, thyroid and adrenal glands presented about half the value found in the former tissues, and aorta, spleen, muscle, and vena cava showed the lowest activity. Low activity was detected in pancreas. For MAO-B, the highest activity was found in heart, liver, duodenum, and kidney (209–322 pmol·min−1·mg prot−1). Half this amount was found in adrenal gland, and negligible activity was observed in pancreas and spleen.
SSAO specific activity was found in all the tissues assayed. Aorta showed the highest activity (338 pmol·min−1·mg prot−1); lung, duodenum, and venous cava showed lower activity (86–133 pmol·min−1·mg prot−1) and the remaining tissues much lower (<54 pmol·min−1·mg prot−1). Comparison of MAO-B and SSAO contribution to benzylamine oxidative deamination indicates that SSAO is the main oxidase in aorta, pancreas, lung, and vena cava, whereas MAO-B is the main oxidase in heart, liver, duodenum, kidney, and adrenal gland.
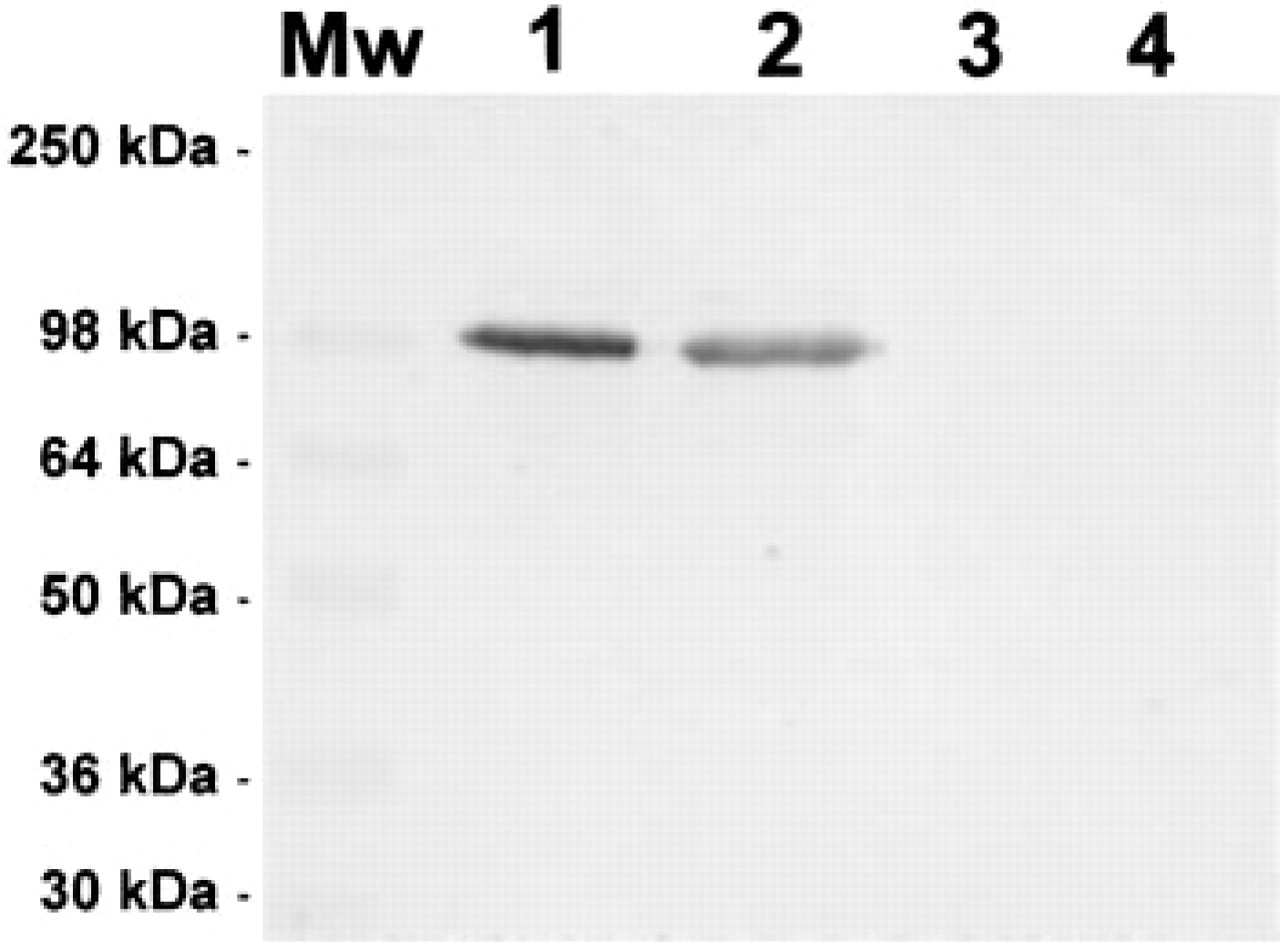
Specificity of the anti-SSAO antibody. Immunoblotting analysis was performed using a rabbit antibody directed against purified bovine lung SSAO. Each sample was electroblotted after 8% SDS-PAGE and stained as described in the text. Lane 1, 10 μg of human heart homogenate; Lane 2, 10 μg of human adrenal gland homogenate; Lane 3, 1 μg of purified porcine kidney diamine oxidase; Lane 4, 10 μg of purified porcine kidney diamine oxidase. Mw, prestained molecular weight standards.
Distribution and Cellular Localization of SSAO Immunoreactivity
To test the specificity of the polyclonal antibody anti-SSAO used in this study, immunoblotting analysis were performed using different human tissue homogenates. Figure 1 (Lanes 1 and 2) shows that human heart and adrenal gland homogenates rendered a single immunoreactive band of 100 kD, similar to the apparent molecular weight of SSAO protein reported in bovine (Lizcano et al. 1998) and rat (Enrique–Tarancón et al. 1998). Furthermore, the possibility that the antibody used might also recognize a related Cu-dependent amine oxidase was discarded. No immunoreactivity was observed when porcine kidney diamine oxidase of 93 kD molecular weight (Schwelberger and Bodner 1997) was assayed (Figure 1, Lanes 3 and 4), and no signal was detected at 84 kD, the molecular weight of the human form (Morel et al. 1992) not commercially available.
MAO-A, MAO-B, and SSAO specific activities in human peripheral tissues (pmolmin−1·mg prot−1) a
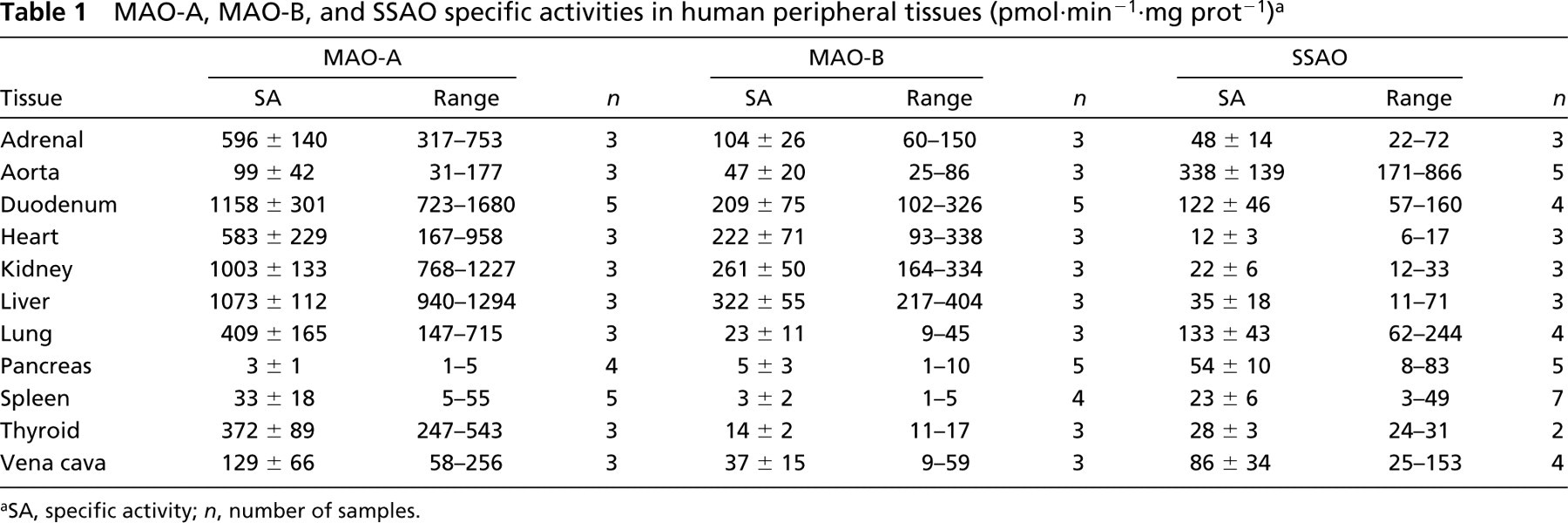
SA, specific activity; n, number of samples.
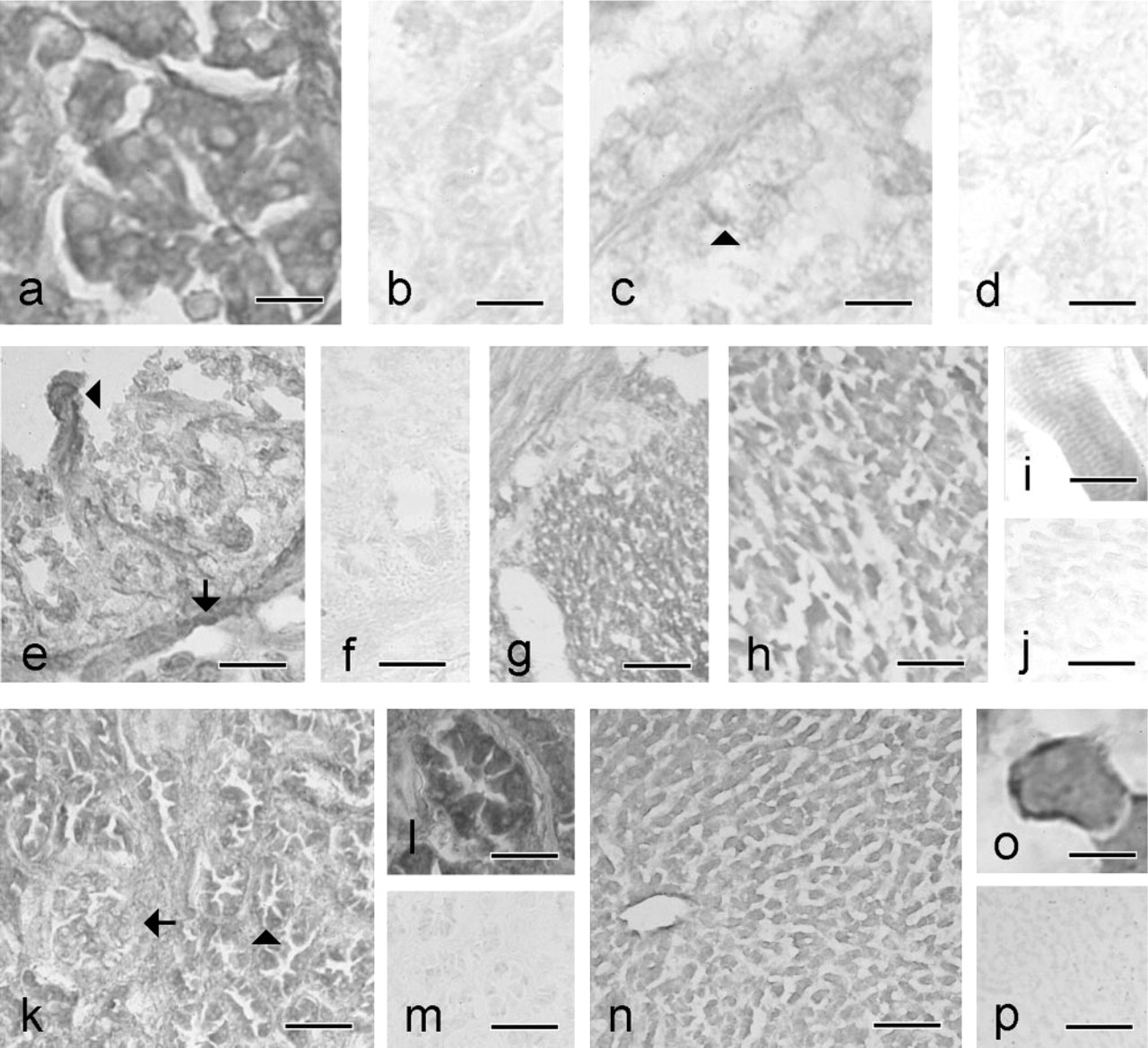
SSAO and 9H7 staining in adjacent sections of adrenal gland, duodenum, heart, kidney, and liver. (
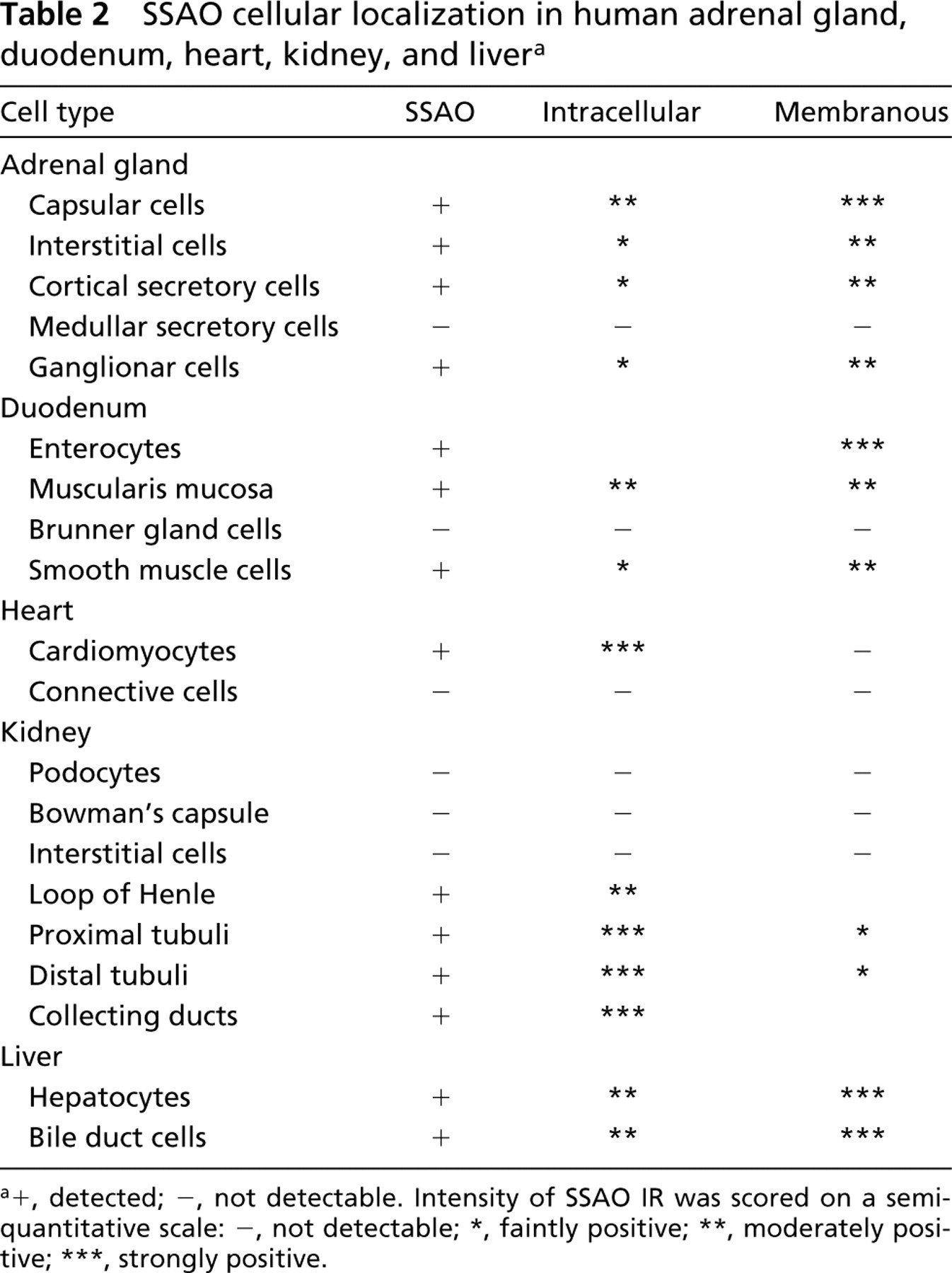
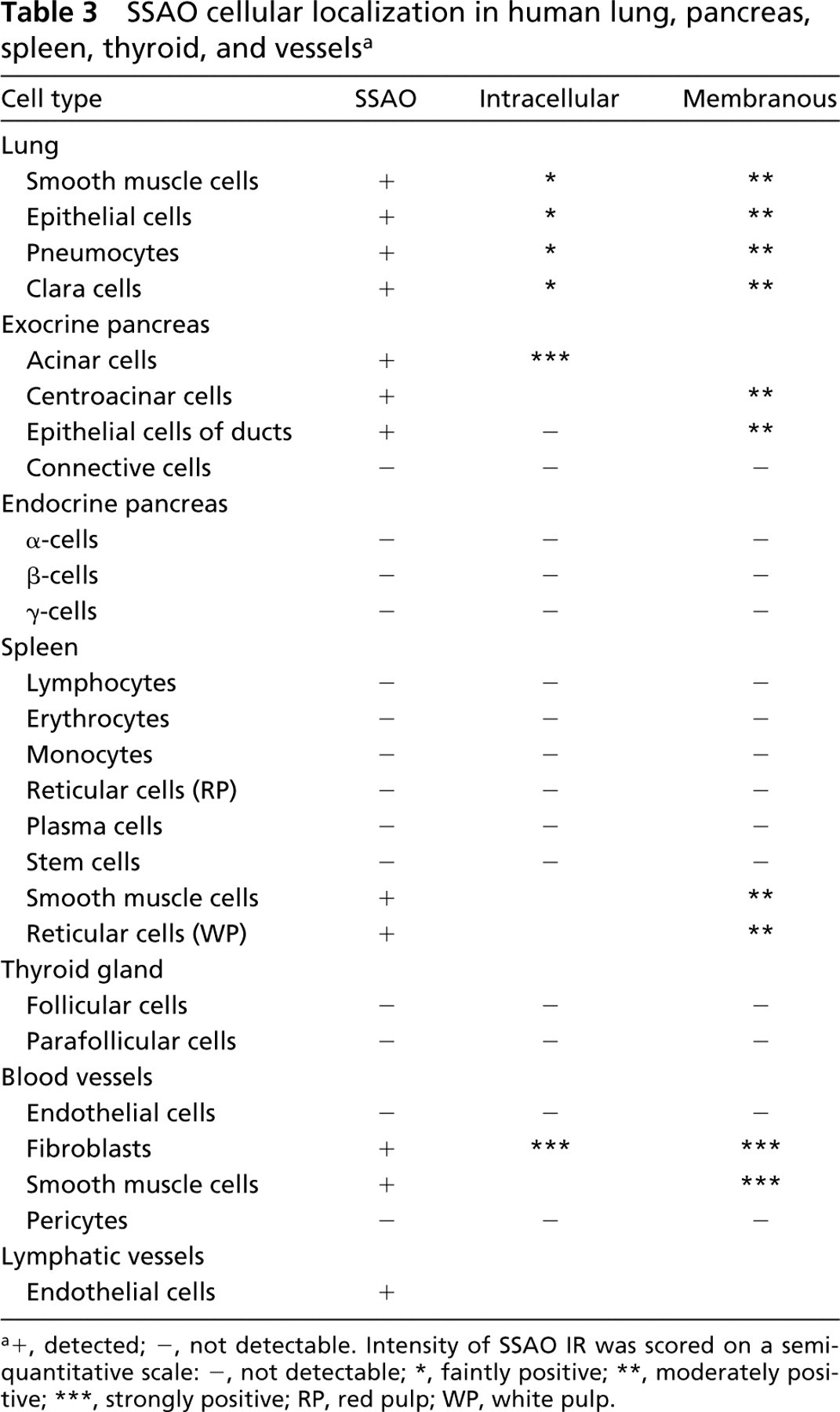
Widespread SSAO distribution was observed in all tissues except the thyroid gland and endocrine pancreas, which were devoid of specific SSAO immunoreactivity (IR) (Figures 2 and 3; Tables 2 and 3). In most tissues, SSAO IR appeared to be bound to the cytoplasmic membrane of the labeled cells. Only a weak and diffuse stain was detected in the presence of the 9H7 antibody. The optimal condition for fixation was found to be 7 hr of immersion in 2% paraformaldehyde. Longer fixation times and higher paraformaldehyde concentrations reduced the intensity of staining. Postfixation with acetone was discarded because of poor tissue preservation. A primary antibody dilution of 1:400 was considered an optimal compromise because higher dilution reduced the signal in several tissues.
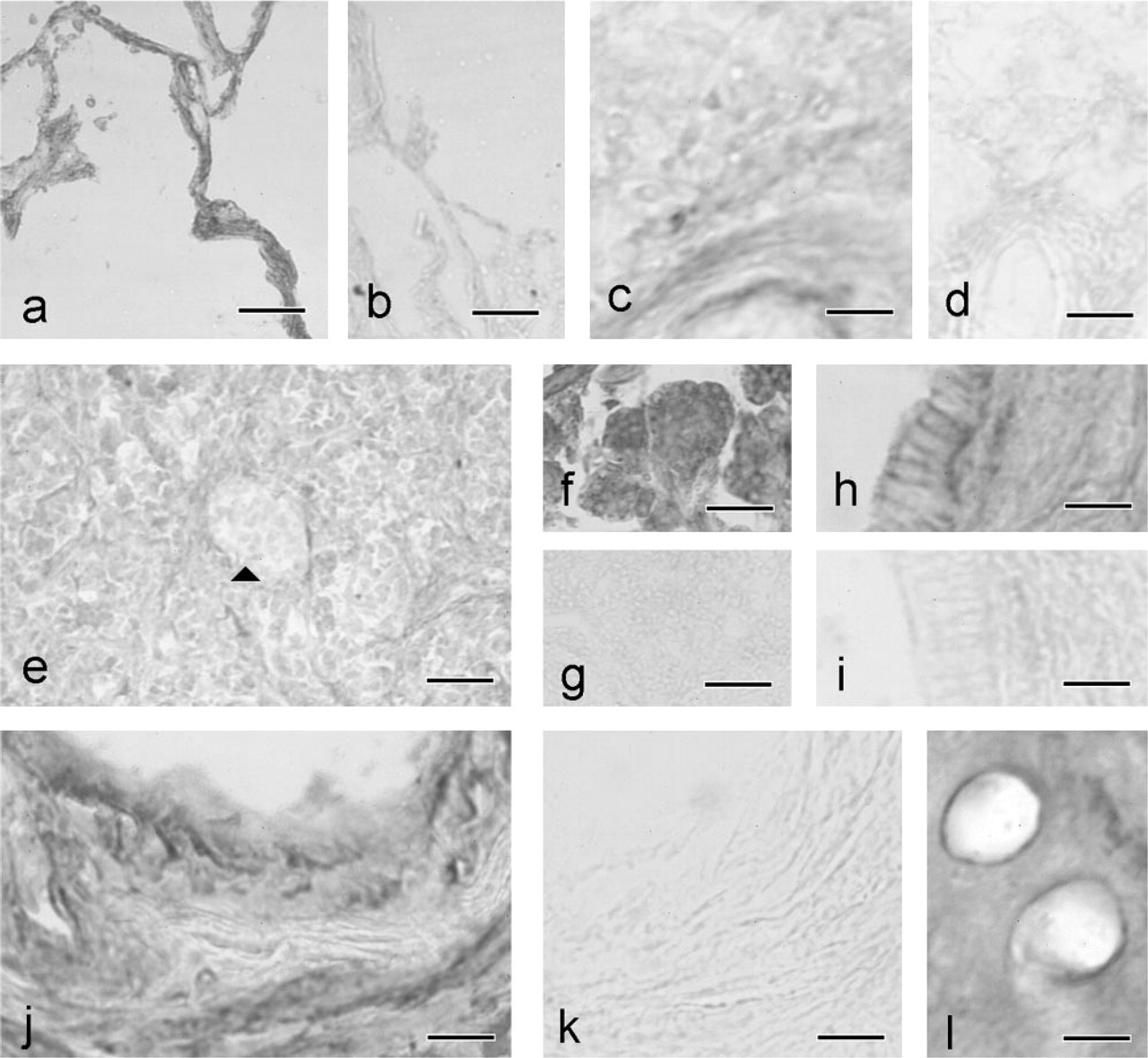
SSAO and 9H7 staining in adjacent sections of lung, spleen, pancreas, and vessels. (
In the adrenal gland, strong SSAO IR was observed in the cytoplasmic membrane, and also intracellularly in the capsule cells. Intracellular positive staining was also seen in both secretory and interstitial cells of the cortex (Figures 2a and 2b). A weak SSAO stain was seen in the medulla, where most cells were negative, and the only SSAO IR was observed in the wall of the central vein and in some scattered cells (Figures 2c and 2d).
In duodenum, SSAO IR was present mostly in the membrane of the enterocytes of villi and Lieberkühn crypts. However, the cytoplasm of the muscularis mucosa and both outer and inner muscular layers showed strong SSAO IR (Figures 2e–2e). No signal was found in the Brunner glands.
SSAO cellular localization in human adrenal gland, duodenum, heart, kidney, and liver a
detected
detected
not detectable. Intensity of SSAO IR was scored on a semiquantitative scale: −, not detectable
faintly positive
moderately positive
strongly positive.
In heart, a strong intracellular SSAO IR was found in the cardiomyocytes but not in the surrounding connective tissue (Figures 2h–2h). In kidney, intracellular SSAO IR was observed in the epithelial cells of renal tubuli, whereas stronger staining was seen in the collecting ducts. Glomeruli were devoid of SSAO IR because neither Bowman's capsule nor podocytes showed specific staining (Figures 2k–2k). Interstitial tissue was also devoid of SSAO staining.
In liver, strong SSAO IR was observed intracellularly and in the membranes of hepatocytes (Figures 2n–2n). A similar pattern was observed in the bile duct and central vein. No portal-to-central vein IR gradient was detected.
In lung, strong staining was detected in bronchiolar cells, both ciliated and Clara cells, and in all pneumocytes of alveoli (Figures 3a and 3b). In this tissue, SSAO IR was stronger in the membrane than in the cytoplasm. In spleen, SSAO staining was observed in the membrane of the capsule and trabecular cells, and in some cells in the white pulp (Figures 3c and 3d). In contrast, no signal was detected in the red pulp, and spleen lymphocytes were negative for SSAO. In the thyroid gland, all the follicular and parafollicular cells were SSAO IR-negative (not shown).
SSAO cellular localization in human lung, pancreas, spleen, thyroid, and vessels a
detected
detected
not detectable. Intensity of SSAO IR was scored on a semiquantitative scale: −, not detectable
faintly positive
moderately positive
strongly positive; RP, red pulp; WP, white pulp.
In the exocrine pancreas, intracellular SSAO IR was seen in all acinar cells (Figures 3e–3e). Centroacinar cells, intercalated ducts, and epithelial cells of pancreatic ducts showed specific IR in the cell membrane (Figures 3h and 3i). The walls of pancreatic ducts and the connective tissue showed no SSAO-specific staining. The endocrine pancreas was devoid of SSAO IR.
In all the organs studied, endothelial cells of arteries and veins were devoid of anti-SSAO staining. However, fibroblasts surrounding the internal elastic lamina of arteries observed at a higher magnification showed strong membrane and intracellular SSAO IR (Figures 3j and 3k). A moderate level of IR was seen in the membrane of vascular muscle-layer cells. In contrast to blood vessels, endothelial cells of lymphatic vessels gave a very strong SSAO IR (Figure 3l). Because these cells are very narrow, we were unable to localize more precisely their SSAO label.
Discussion
We have studied the human tissue and cellular distribution of SSAO using a polyclonal SSAO antibody (Castillo et al. 1998; Lizcano et al. 1998). MAO-A-, MAO-B-, and SSAO-specific activities have also been determined in these tissues to help understand the physiological role of these oxidases. In studying human postmortem samples, factors such as age, postmortem delay, and cause of death are a source of variability. The present study should be regarded as a description of the SSAO activity and localization in tissues of aged humans, which may not be applicable to younger individuals. We excluded tissues that might have been affected by the pathological condition of the donor, and because the pattern of distribution for a given tissue was the same in all the cases included, we are confident that the results reflect the non-pathological condition.
Our results confirm the presence of MAO-A, MAO-B, and SSAO in most human peripheral tissues, with a higher MAO-A specific activity. In agreement with previous reports (Lewinsohn 1984; Lyles 1996), SSAO specific activity was high in aorta and lung, whereas heart, spleen, and thyroid gland presented low activity. Values obtained in aorta, skin, and liver were similar to those previously described (Lewinsohn 1984; Yu et al. 1994). Human heart SSAO activity was much lower than that described for rat (Lyles and Callingham 1975). These species differences, also reported for MAO-A and -B (Saura et al. 1994, 1996) and for SSAO (Morgan 1985; Lizcano et al. 1991), should be kept in mind when experimental data are extrapolated from rodent studies to humans.
SSAO specific activities found in this study are mainly in accordance with their tissue distribution revealed by immunohistochemistry. The high specific activity and strong immunostaining found in highly vascularized tissues such as duodenum, liver, lung, and blood vessels, but not heart, could reflect the metabolic deamination of the circulating amines of endogenous or exogenous origin.
The presence of SSAO in adrenal gland, kidney, exocrine pancreas, and lymphatic vessels in cells lacking MAO-B (Saura et al. 1997; Rodríguez et al. 2000) suggests its participation in amine oxidation, as proposed for vascular smooth muscle (Lewinsohn 1984; Lyles 1996). The adrenal gland is the main producer of hormonal catecholamines. Therefore, it is interesting to note that we found no SSAO IR in chromaffin cells. However, we found SSAO IR in the central vein, ganglionar, capsular, and interstitial cells. The absence of both SSAO and MAOs in human chromaffin cells (Rodríguez et al. 2000) suggests that they lack uptake mechanisms for amines that will be inactivated in the target organs. It is important to note the presence of SSAO IR in the capsule and interstitial cells because they were devoid of MAO-B (Rodríguez et al. 2000). In the adrenal cortex, SSAO IR was present in the membrane and cytoplasm of all secretory cells, where it might reinforce the MAO-B autocrine regulation (Rodríguez et al. 2000). We found that SSAO is localized in all renal tubuli. Because dopamine may be degraded by MAO-B and SSAO (Lizcano et al. 1991), both enzymes may be involved in the degradation of dopamine synthesized in the kidney (Lee 1982).
In the production of pancreatic juice, there is a contribution from two cell types from the exocrine pancreas: acinar cells, producing digestive enzymes and co-factors, and centroacinar/duct cells, producing bicarbonate and water. We found SSAO in both cell populations where MAO-A is also present, whereas MAO-B appears only in the centroacinar/duct cells (Rodríguez et al. 2000). Sympathetic norepinephrine is unlikely to be a MAO substrate in these cells because the sympathetic innervation of the exocrine pancreas is mainly, if not entirely, vascular (Magee 1989). Circulating monoamines are the most likely SSAO and MAO substrates in these cells, but monoamine uptake mechanisms have not been described. Another candidate is dopamine, because it modulates the pancreatic flow rate in rat (Mori et al. 1979b) and accumulates in rat acinar cells after nonspecific MAO inhibition (Mori et al. 1979a). Moreover, rat acinar cells synthesize dopamine (Mori et al. 1979b), which inhibits amylase and bicarbonate secretion in humans (Caldara et al. 1978).
In addition, SSAO IR was observed only in the membrane of the smooth muscle of the blood vessels, probably related to the sympathetic regulation of blood pressure (Lyles 1996). Strong IR for SSAO was found in vascular fibroblasts, intracellularly and in the cytoplasmic membrane, which is consistent with the known presence of SSAO in blood vessels, predominantly associated with the smooth muscle cells (Castillo et al. 1998), and may be related to the reported secreted soluble form of SSAO from cultured vascular smooth muscle cells (Hysmith and Boor 1987). However, SSAO was absent in endothelial cells of blood vessels but present in endothelial cells of lymphatic vessels. VAP-1, an adhesion protein with the same primary structure as tissue-bound SSAO, has been located in the endothelial wall of the high endothelial venules (Bono et al. 1999). Expressed in inflammatory conditions, it binds to lymphocytes and helps them to migrate between endothelial cells into lymphoid organs (Jaakkola et al. 1999). The presence of SSAO IR in lymphatic vessels and brain microvessels (Castillo et al. 1998), but not in endothelial cells of large blood vessels, might reflect its duality as an enzyme and an adhesion protein. Because the aorta and other tissues used in this work to determine SSAO activity presented no inflammation, SSAO was not expressed in the blood vessels, whereas SSAO expression would have been induced in human brain microvessels, which are narrower and more vulnerable.
In cardiomyocytes and acinar cells, SSAO localization was mainly intracellular, but it was mainly membranous in adrenal gland, duodenum, liver, lung, and exocrine pancreas. Both liver and lung contribute to the degradation of physiological amines. The membrane and intracellular SSAO IR localization observed in these tissues suggests that it might also modulate hormonal action. Thus, as shown in adipocytes (Marti et al. 1998), in addition to its role as a scavenger of circulating amines, human SSAO might influence some physiological processes through the generation of H2O2 (Lyles 1996; Enrique–Tarancón et al. 1998). In spleen, SSAO in the capsule and trabeculae of the membrane of the smooth muscles may also have a role other than amine oxidation.
We found only two tissues devoid of SSAO IR: the thyroid gland, rich in MAO-A, and the endocrine pancreas, with MAO-A localized mainly in β-cells and MAO-B restricted to α-cells (Rodríguez et al. 2000). The absence of SSAO IR in these two glands might indicate that MAO hydrogen peroxide generation may modulate hormone secretion without the participation of SSAO, as proposed by other authors (Pizzinat et al. 1999).
In conclusion, these results are the first attempt to quantify and map SSAO localization in human peripheral tissues. As suggested by its cell localization, its role may not be restricted to amine oxidation. Further experiments are necessary to provide such evidence and understand the physiological importance of SSAO in humans.
Footnotes
Acknowledgments
Supported by SGR 99-224 from the Generalitat de Catalunya and the DGES grant SAF99-0093 from the Ministerio de Educación y Cultura.
We wish to thank Dr J. A. Bombí (Departament de Biologia Cel·lular i Anatomia Patològica, Facultat de Medicina, Universitat de Barcelona, Spain) for the gift of the human tissue samples, and Dr Ellen E. Billett (Department of Life Sciences, Faculty of Science and Mathematics, The Nottingham Trent University, UK) for the gift of the 9H7 antibody.