
Abstract
C
Calcineurin has been thought to be important in various aspects of cell functions, including those of the immune system and nervous system. This is exemplified by the effect of the immunosuppressive drugs (O'Keefe et al. 1992) FK506 and cyclosporin A and the inhibition of T-cell activity through dephosphorylation of a transcription factor, nuclear factor of activated T-cell (NF-AT) (Muramatsu and Kincaid 1996). Another notable function of calcineurin is the regulation of Ca++-induced apoptosis, which is suppressed by the anti-apototic protein Bcl-2 (Shibasaki et al. 1997). Evidence of its functions in the brain is accumulating, including long-term depression (LTD) (Mulkey et al. 1994) and long-term potentiation (LTP) (Mansuy et al. 1998; Sprengel et al. 1998). The function of calcineurin in brain tissue is under complex regulation: calcium channels in the endoplasmic reticulum, neural nitric oxide synthase (nNOS) activity, and neurotransmitter release through the regulation of voltage-gated calcium channels on presynaptic membranes (for review see Snyder et al. 1998).
Although the presence of calcineurin has been reported in chick retina (Cooper et al. 1985) and the lateral eyes of a crab (Ellis and Edwards 1994), little is known about its presence in the mammalian retina and its roles in visual function and the development of the mammalian retina. In this study, retinal tissues of three kinds of mammalian were examined by immunoblotting analysis and immunohistochemistry. The presence of calcineurin in these retinas and in developing rat retinas was demonstrated.
Materials and Methods
Antibodies
Three kinds of polyclonal antibodies against peptides of the deduced amino acid sequences of the N-terminal regions of the human and murine calcineurin catalytic subunit A, Aα, and Aβ isoforms were prepared (Giri et al. 1992; Usuda et al. 1996). Rabbits were immunized with each peptide conjugated with keyhole limpet hemocyanin through a cysteine added to the N-terminus. The IgG fractions of the antisera were affinity-purified against each peptide. These antibodies to the peptides of the amino acid sequence and that common to both isoforms (KAVFPPSHRLT) and to the respective isoforms of Aα (DPKLSTTDRVV: amino acids 8–18) and Aβ (PPPPGADRVVK: amino acids 18–28) were designated Amino X antibody, ACN-2, and ACN-3. The latter two have been employed in a previous immunohistochemical study (Usuda et al, 1996).
Processing of the Tissues
Retinal tissues of Wistar rats at various developmental ages [embryonic Day 18 (E18), postnatal Days 1 (P1), 4 (P4), 7 (P7), and 14 (P14)], retinal tissues of male Wistar rats at 4 and 12 weeks, and mature bovine retinal tissues obtained from a local slaughterhouse were used. The rats were sacrificed under deep anesthesia with diethylether. Tissues were fixed in 4% paraformaldehyde/100 mM sodium phosphate, pH 7.4, for 24 hr at 4C by immersion, after washing for 3 hr with 100 mM lysine/100 mM sodium phosphate, (pH 7.4)/150 mM sodium chloride to quench free aldehyde groups. Human retinal tissues were obtained at autopsy from three patients with no clinical evidence of neurological disease. These cases had acute respiratory failure (n = 1, age 85, male) and liver cirrhosis (n =2, ages 53 and 63, male). These human specimens were fixed in 10% neural buffered formalin for 24–48 hr. After dehydration in ethanol and xylene, they were embedded in paraffin.
Light Microscopic Immunohistochemistry
The tissues were dehydrated in an ethanol and xylene series, embedded in paraffin, and sectioned at 5 μm. After deparaffinization in xylene/ethanol series and rehydration, sections were treated in a microwave oven (MR-M201; Hitachi, Tokyo, Japan) for antigen retrieval in 10 mM sodium citrate, pH 6.0, for 10 min (Shi et al. 1993). Staining was performed according to the ABC technique (Hsu et al. 1981) at room temperature. The staining sequence was as follows: 2% goat serum for 30 min; primary antibodies of Amino X, ACN-2, ACN-3 (2 μg/ml of IgG); biotinylated anti-rabbit IgG for 1 hr; avidin–biotinylated peroxidase complex for 1 hr. Solutions for antibody dilution were made with 20 mM sodium phosphate, (pH 7.4)/150 mM sodium chloride containing 2% normal goat serum. Sera for the blocking step, biotinylated antibodies, and avidin–biotinylated peroxidase complex were purchased from Vector Laboratories (Burlingame, CA). The reaction products were developed by incubation in 0.05% 3,3′-diaminobendizine tetrachloride and 0.01% H2O2 in 50 mM Tris-HCl, pH 7.5, for 10 min. They were observed with a Nikon FXA microscope after counterstaining with 1% methyl green solution. Negative controls for ACN-2 and ACN-3 antibodies were done as reported previously (Usuda et al. 1996). The negative controls for Amino X antibody were made by adding the peptide of Amino X or recombinant proteins of the calcineurin Aα or Aβ isoform to the primary antibody, and by omitting the primary antibody.
Immunoblotting Analysis
Tissues were frozen at –80C until analysis. Rat, bovine, and human retinal tissues and whole rat brain tissue were homogenized in 10 volumes of 10 mM potassium phosphate, (pH 7.5)/150 mM sodium chloride containing the protease inhibitors pepstatin (4 μg/ml), PMSF (3.5 μg/ml), leupeptin (4 μg/ml), and E-64 (4 μg/ml). These homogenates were dissolved in a sample buffer solution containing 1% sodium dodecyl sulfate (SDS) and 200 mM 2-mercaptoethanol, subjected to SDS-polyacrylamide slab gel electrophoresis, transferred to nitrocellulose membranes, and stained with each of the primary antibodies for calcineurin (Amino X, ACN-2, and ACN-3). Goat anti-rabbit IgG antibody conjugated with alkaline phosphatase and bromochloro-indolyl phosphate/nitroblue tetrazolium (BCIP/NBT) were sequentially applied to each membrane for visualization. For analysis of retinal development, whole eyeballs were subjected to electrophoresis, because the retina could not be isolated from the eyes of embryos or newborn animals. This electrophoresis was done with samples of the same amount of protein or of the amount of protein corrected by the volume ratio of retina:total eyeball measured on light microscopic specimens. The negative controls for Amino X antibody were made by adding the peptide of Amino X or recombinant proteins of the calcineurin Aα or Aβ isoform to the primary antibody. The amount of protein was measured by protein assay CBB solution (Nakarai; Kyoto, Japan) with bovine serum albumin as a standard.
Results
Immunoblotting Analysis
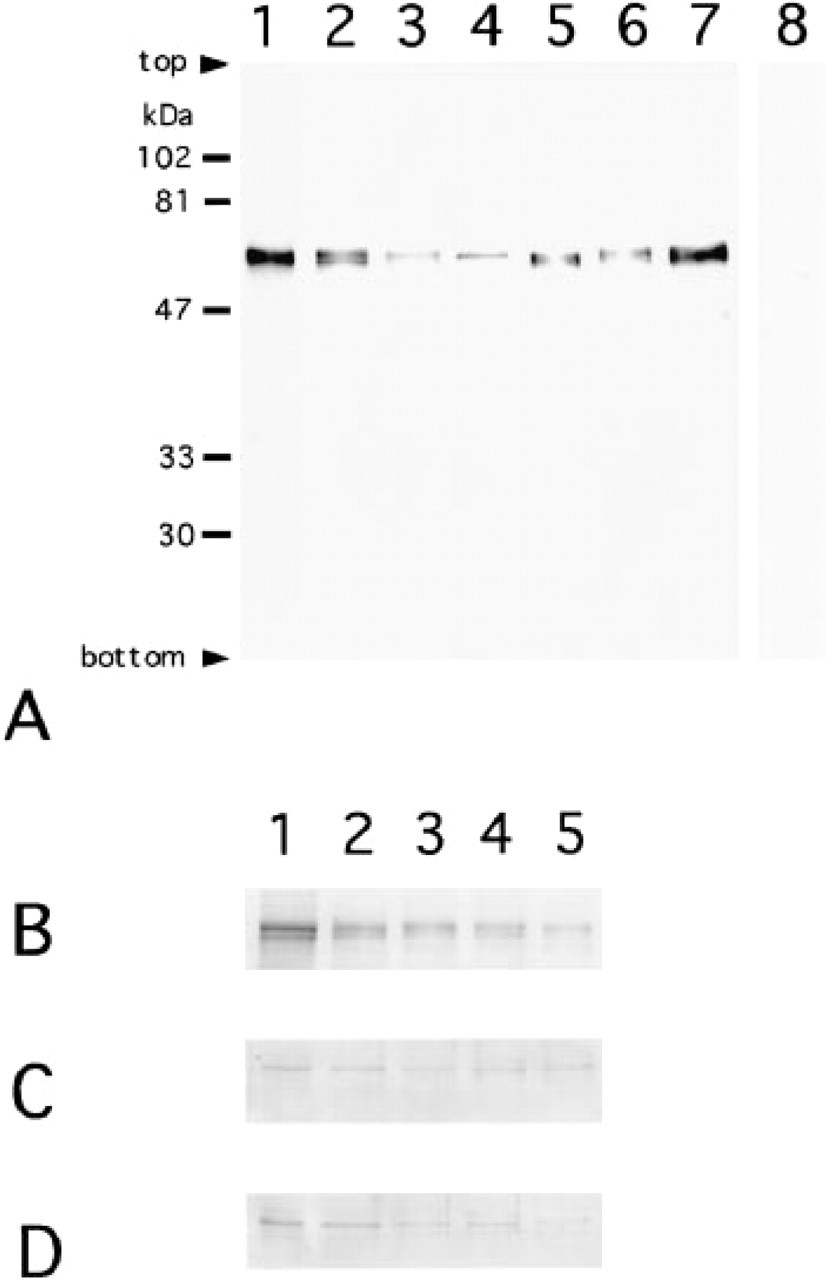
Amino X antibody detected signals in all tissues examined (Figure 1A). The mobility of each signal for rat brain, rat, bovine, and human retinas was the same as that of the corresponding recombinant proteins of calcineurin A subunit, both Aα and Aβ isoforms. The mobilities of the two isoforms were very close and their mixture showed a broad signal. In the staining controls, the addition of recombinant proteins or synthetic polypeptides to the Amino X antibody rendered the staining weak or negative. The presence of the calcineurin A subunit, both Aα and Aβ isoforms in E18, P4, P7, P14, and mature rat eyeballs was detected with Amino X, ACN-2, and ACN-3 antibodies (Figures 1B–1B). The signals for Amino X antibody were weaker in the samples from postnatal animals than in embryonic samples in both cases when the same amounts of protein were analyzed (Figure 1B) or the amounts of protein corrected by the volume ratios of retina:total eyeballs were analyzed (data not shown). ACN-2 and ACN-3 antibodies detected signals for Aα and Aβ isoforms in all samples of developmental rat eyes (Figures 1C and 1D). The signals for ACN-3 were more easily detectable than ACN-2 when signals were developed (data not shown). These data indicated the presence of two isoforms in the retina of the three mammalians and showed that the two are present in rat eyes through all developmental stages, in amounts that do not increase with increasing development.
Immunohistochemistry of the Mature Retina
The localization of the calcineurin A subunit in the mature retina is shown in Figure 2. The ganglion cell layer, inner plexiform layer, and inner nuclear layer of the mature rat retina were strongly positive for the reaction of Amino X antibody (Figure 2A). The ganglion cells and amacrine cells in the inner nuclear layer were the most strongly immunoreactive. Other kinds of cells in the inner nuclear layer, bipolar and horizontal cells, and the outer plexiform layer were also immunopositive. In the rat inner plexiform layer, laminae 1, 2, and 3 were visible as well-delineated bands, and the inner segment of photoreceptor cells was weakly positive. Similar to the immunoreaction of the mature rat retina, the ganglion cell, inner plexiform, and inner nuclear layers in the bovine and human retinas reacted strongly to the Amino X antibody (Figures 2B and 2C). The ganglion cells and both the amacrine and bipolar cells of the inner nuclear layer were strongly immunoreactive, with the horizontal cells somewhat less so. The inner plexiform layer showed diffuse positive reactivity without well-delineated bands. The inner segment of the photoreceptor cells was weakly positive. In the staining controls, recombinant proteins or synthetic polypeptides reduced the staining of Amino X antibody to weak or negative (Figure 2D).
The reaction products for Aα and Aβ isoforms detected by ACN-2 and ACN-3 were observed in ganglion cells and the inner nuclear layer. The staining of ganglion cells was distinctive, in that subcellular localization in these cells differed between ACN-2 and ACN-3. Aα and Aβ isoforms were localized in the nucleus for ACN-2 (Figures 3A, 3C, and 3E) and in the cytoplasm for ACN-3 (Figures 3B, 3D, and 3F) in rat, bovine, and human retinas. The staining in the inner nuclear layer was weak for ACN-2 antibody (data not shown). The control for the staining employing recombinant proteins or synthetic polypeptides made the staining weak or negative (data not shown).
(
Immunohistochemistry of Calcineurin in the Developing Rat Retina
Developmental changes in the location of the calcineurin A subunit were examined in rat retinas using the Amino X antibody. At E18 the reaction appeared rather diffuse overall, preferentially localized in the inner half of the retina (Figure 4A), the region separated from the neuroblastic layer by the inner plexiform layer. The reaction products become concentrated in specific kinds of cells thereafter. With P4, the reaction products observed in the ganglion cell and the inner plexiform layers were strongly positive, and the cells in the neuroblastic layer above the inner plexiform layer (which appeared to be amacrine cells) were positive (Figure 4B). With P7, the ganglion cell, inner plexiform, inner nuclear, and outer plexiform layers all become positive. Ganglion cells and the amacrine and horizontal cells in the inner nuclear layer were strongly positive, and their immunopositive nerve cells possessed processes extending into the inner plexiform layer. Two intensely immunoreactive bands were observed in the inner plexiform layer (Figure 4C). With P14, the ganglion cell, inner plexiform, inner nuclear, and outer plexiform layers were immunopositive. Ganglion cells and the amacrine cells in the inner nuclear layer were strongly positive, and the bipolar cells were also positive. The horizontal cells were weakly labeled, although the labeling may have been masked by the intense labeling of other kinds of cells in the inner nuclear layer. Three immunoreactive bands were observed in the inner plexiform layer. At this stage, both inner and outer segments are present in the retina and were weakly immunopositive (Figure 4D). The reaction product for the calcineurin A subunit was observed in the ganglion cell layer through these developmental stages from the embryonic stage. Other kinds of cells became positive later.
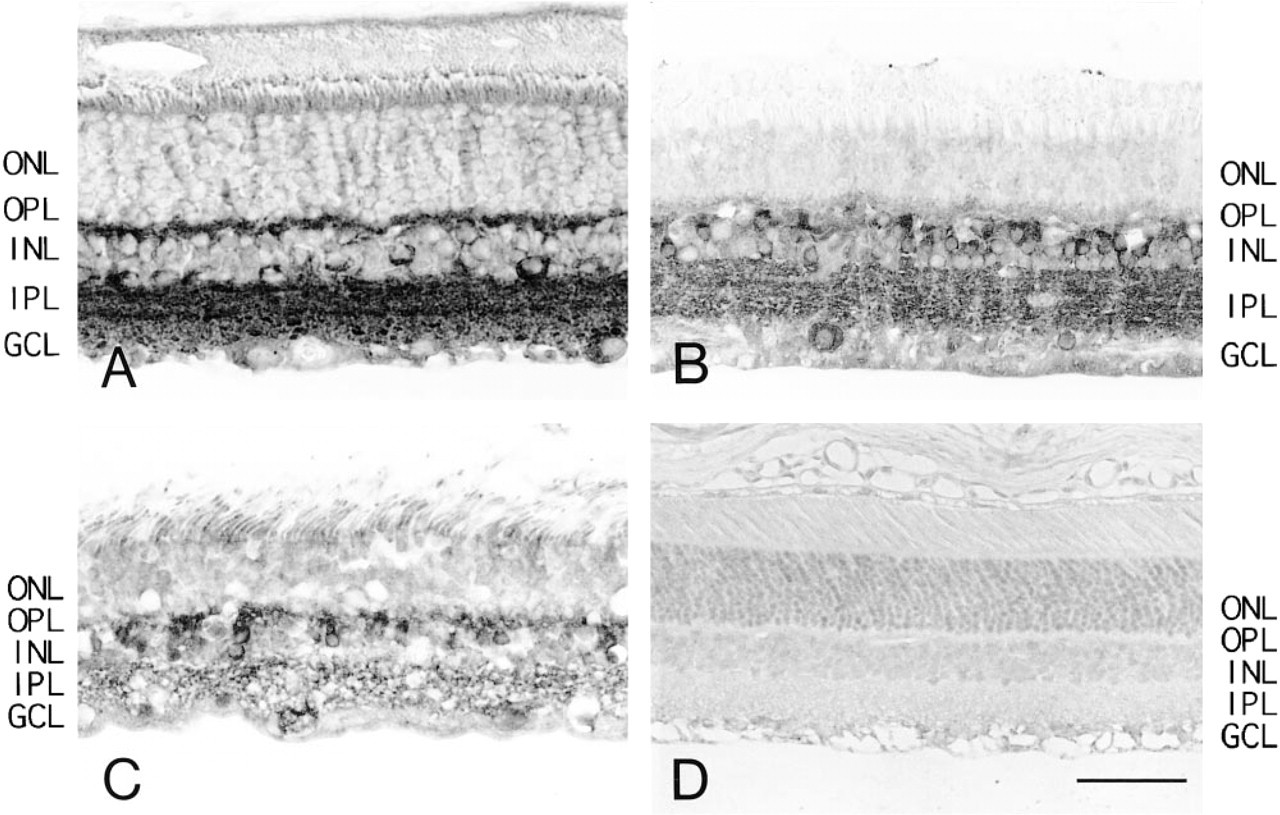
Immunohistochemistry of mature retina by Amino X antibody for calcineurin A subunit: rat (
Discussion
In the preliminary experiment for localizing calcineurin in the retina, the antibody was raised against purified protein of its A subunit. This antibody was not suitable for the immunostaining because the reaction was not inhibited by adding the recombinant proteins of the A subunit. Amino X antibody was made for the peptide of the common sequence near the N-termini of Aα and Aβ isoforms of mouse calcineurin (Parsons et al. 1994). This amino acid sequence is common to the Aα isoform of rat, bovine, human, and mouse enzymes (Ito et al. 1989; Kincaid et al. 1990; Muramatsu and Kincaid 1993; Griffith et al. 1995). It is also present in the amino acid sequence of the Aβ isoform of rat, human, and mouse enzymes (Kuno et al. 1989; Guerini and Klee 1989; Giri et al. 1991). The amino acid sequence of Aβ isoform of bovine enzyme has not been reported. As shown in the present study, Amino X antibody was immunoreactive for recombinant proteins of Aα and Aβ isoforms of human calcineurin. It detected signals in rat brain and in rat, bovine, and human retinal tissues by immunoblotting analysis. The Amino X antibody, in addition to the subtype-specific antibodies ACN-2 and ACN-3 (Giri et al. 1992; Usuda et al. 1996), was employed in the present immunohistochemical study. The immunohistochemical reaction using the Amino X antibody was inhibited by each recombinant protein of Aα and Aβ isoforms and by the synthetic antigen polypeptide. This Amino X antibody was shown to be useful in detecting calcineurin or its A subunit of both isoforms by immunoblotting analysis and immunohistochemistry. The Amino X antibody was also employed on chick retina by immunoblotting analysis and immunohistochemistry (data not shown) as preliminary experiments. Strong immunoreactivity was observed in ganglion cells, amacrine cells, and the inner plexiform layer. The outer plexiform layer and inner segment of photoreceptor cells were weakly positive. These data agreed with the previous report by Cooper et al. (1985) showing the localization of calcineurin in the nerve terminals of photoreceptor cells and bipolar cells of chick retina employing a polyclonal antibody against purified bovine brain calcineurin.
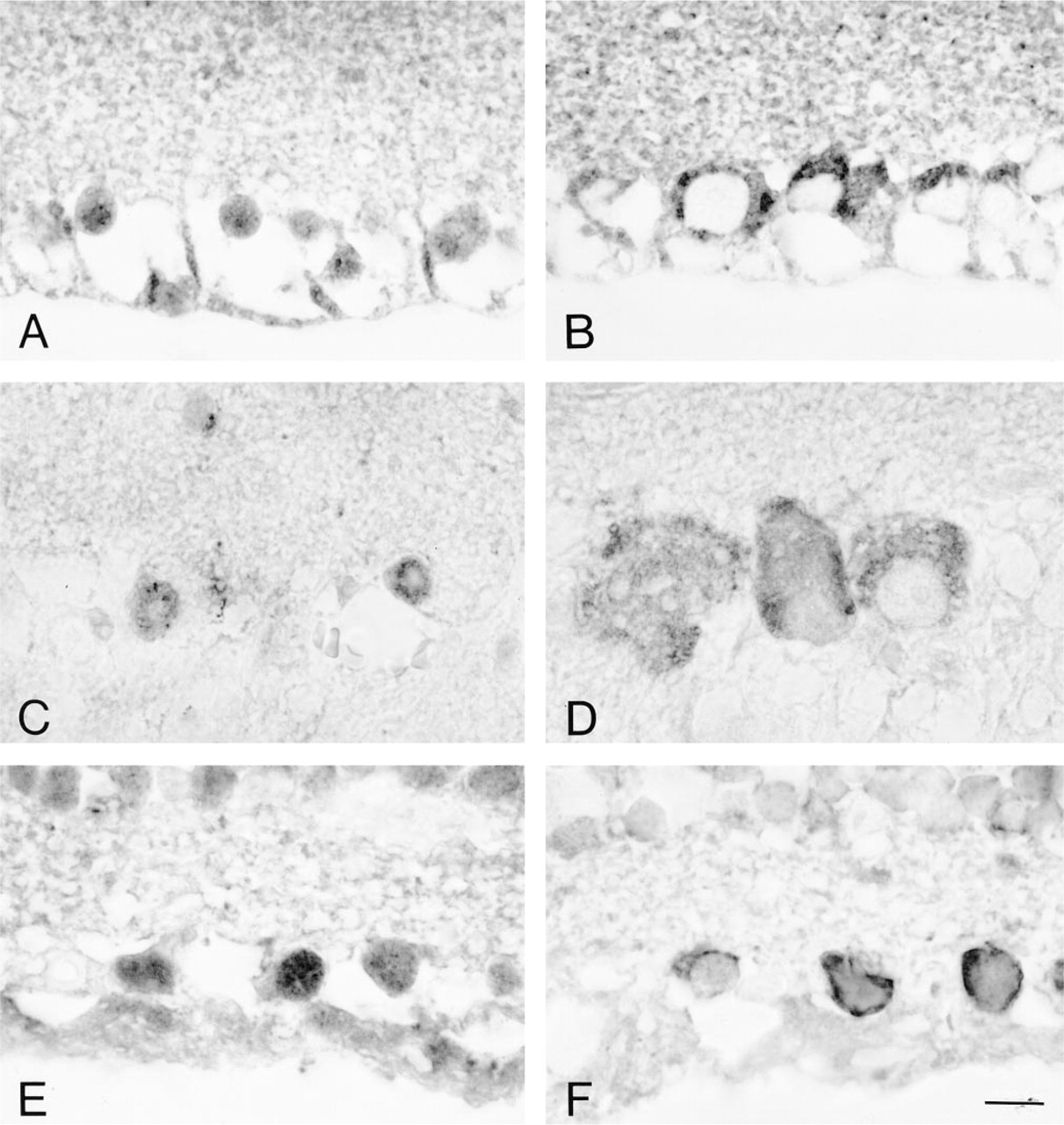
Immunohistochemistry of mature retina to detect calcineurin A subunit Aα and Aβ isoforms: rat (
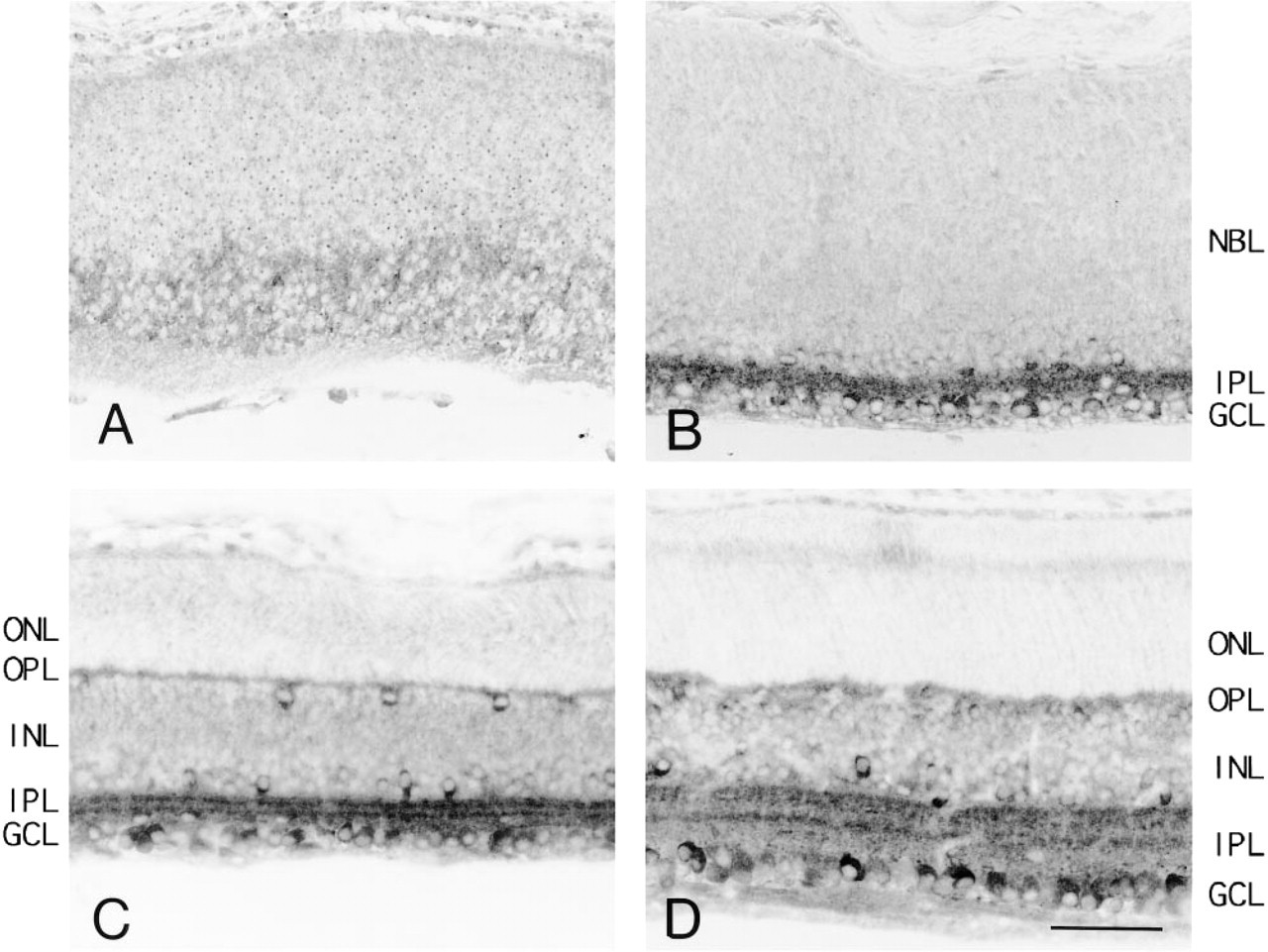
Immunohistochemistry of the developing rat retina by Amino X antibody for calcineurin A subunit. Embryonic Day 18 (
In the tissues examined, the presence of calcineurin was confined to retinal neurons and was not detected in Müller cells. This preferential localization in the neuron was similar to that in the brain, as shown by in situ hybridization and immunocytochemistry (Takaishi et al. 1991; Steiner et al. 1992; Usuda et al. 1996). The site-specific localization of Aα and Aβ isoforms in the nucleus and in the cytoplasm of the ganglion cells also reflects the differential distribution of isoforms observed in the nerve cells of the brain (Usuda et al. 1996). The Amino X antibody detected the localization of calcineurin mainly in the cytoplasm. The reaction of the ACN-2 antibody was weaker than that of ACN-3 when it was applied to immunoblotting analysis and immunohistochemistry of the retina. The immunoreaction was rather easily detectable with Amino X and ACN-3 antibodies in various kinds of nerve cells in the retina. The immunoreaction in the nucleus of ganglion cells with ACN-2 antibody was consistently observed, but that in other kinds of nerve cells, including amacrine cells whose immunoreaction was easily detected by Amino X and ACN-3 antibodies, was very weakly positive with ACN-2 antibody. Previous immunohistochemical analysis reported stronger immunoreactivity for the Aα isoform than for the Aβ isoform in the brain (Goto et al. 1986). In the retinal tissues examined in the present study, the Aβ isoform was more easily detectable than Aα using ACN-2 and ACN-3 antibodies, both by immunoblotting analysis and immunohistochemistry. The relative abundance of the Aβ isoform compared with Aα in the retina might explain the stronger staining of the cytoplasm by Amino X and ACN-3 antibodies than by ACN-2 antibody.
All three antibodies to calcineurin A subunit detected signals in the developing eye tissues of the rat by immunoblotting analysis. The intensity of the signals decreased with development. However, by immunohistochemistry, the localization of calcineurin changed during development, from a rather diffuse pattern limited to the inner half of the retina at E18, before neural layers are distinguishable, to concentration in the ganglion cells and amacrine cells in the inner nuclear layer after birth. In the blotting analysis there was a tendency for antibody labeling to decrease, whereas in the sections there may be an opposite trend. These results from two different procedures are apparent but contradictory. The reason for this is not clear, but the different pattern of localization (a diffuse pattern in the immature retina and specifically localized in particular areas in the mature retina) may explain this contradiction. Summarizing these observations: (a) calcineurin is present in the retina of three mammalian species, including humans; (b) calcineurin is most abundant in the ganglion cells and cells in the inner nuclear layer; (c) Aα and Aβ isoforms are differentially located in the nucleus and in the cytoplasm of ganglion cells; and (d) both isoforms, Aα and Aβ, are already present in the eye of the developing rat, but their rather diffuse presence in the immature retina becomes confined to ganglion cells and amacrine cells after the developmental period.
In our study, calcineurin was shown to be present in the retinal neurons. The functions of Ca++ in the retina have been studied extensively in the photoreceptor cells (for review see Yarfitz and Hurley 1994), and Ca++-binding proteins and CaM kinases have been reported to be present mainly in retinal neurons other than photoreceptor cells. The preferential localization of calcineurin in specific kinds of neurons, ganglion cells and amacrine cells should be compared with that of Ca++- or CaM-dependent kinases. The Ca++- and/or CaM-dependent protein kinases have been considered in general to function coordinately with the Ca++/CaM-dependent phosphatase calcineurin (Klee et al. 1998). Protein kinase C has been localized in ganglion, amacrine, and bipolar cells but in neither photoreceptor nor in Müller cells in mature rabbit retina (Usuda et al. 1991). Similarly, CaM kinase II, the most abundant Ca++/CaM-dependent protein kinase in the brain, and CaM kinase IV have been localized in the ganglion cells, the inner plexiform layer, and the amacrine cells, but not in the photoreceptor or Müller cells in mature rat retina (Ochiishi et al. 1994; Sakagami and Kondo 1996). Co-localization of Ca++- binding proteins, Ca++/CaM kinases, and calcineurin in the same kinds of cells in the retina appears to be rational. CaM kinase II and CaM kinase IV are known to phosphorylate calcineurin in vitro (Hashimoto et al. 1988; Kasahara et al. 1999). The anchoring protein of protein kinase A (AKAP79) has binding sites both for protein kinase C and calcineurin (Klauck et al. 1996). Functional evidence suggesting that calcineurin, and protein kinase C or CaM kinases act in regulating Ca++/CaM-dependent functions concerning neurotransmitter release and long-term memory in nervous tissue has also been reported (For review see Snyder et al. 1998). Regarding the function of Ca++-binding proteins and CaM kinases in the retina outside the photoreceptor cells, nothing has been clarified except for the possibility that CaM kinases may phosphorylate CREB in cells in the ganglion cell layer and inner nuclear layer (Yoshida et al. 1996). Information on the role of calcineurin in the retina is also limited. The existence of FK506-binding protein, the binding protein of the inhibitor of the function of calcineurin, has been demonstrated in the retina by RT-PCR (Kitagawa et al. 1996). GABA-mediated currents in ganglion cells were shown to be mediated by calcineurin in the retina (Akopian et al. 1998). Calcineurin was presumed to mediate glutamate-induced neurotoxicity in cultured rat retinal cells, against which they can be protected by administering FK506 (Kikuchi et al. 1998). However, there is no functional evidence for the coincidental localization of calcineurin and either CaM kinase II or protein kinase C in the retina.
In the present study, the calcineurin isoforms Aα and Aβ were localized mainly in the nucleus and the cytoplasm, respectively, the same as neurons of the central nervous system (Usuda et al. 1996). Evidence showing that the presence of calcineurin is important both in the nucleus and the cytoplasm is accumulating. The most representative functions of calcineurin are related to apoptosis, LTP, and LTD, as described in the Introduction. Their molecular mechanisms have been partly clarified, as a modification of the phosphorylation state of intranuclear transcription factors. The immune response related to calcineurin, i.e., the mechanism of T-cell activation, is regulated by dephosphorylation of NF-AT. Dephosphorylation of NFAT4 by calcineurin suppresses its import into the nucleus (Chow et al. 1997). LTP is also regulated by calcineurin. The activity of CREB, an important transcription factor that regulates memory (Yin et al. 1995; for review see Snyder et al. 1998), is regulated by its phosphorylation state. Calcineurin is a negative regulator for CREB-dependent transcription in gene expression in LTP (Bito et al. 1996). The difference in the function of the calcineurin A isoforms, Aα and Aβ has not been clarified yet.
To understand the presence of calcineurin in the retina, it is also important to consider its localization in relation to the presence and the function of other Ca++-binding proteins in the retina. Calcineurin comprises two subunits, of which the catalytic subunit (A) has serine/threonine activity, while the regulatory subunit (B) binds the A subunit in a Ca++-dependent manner involving CaM. Therefore, calcineurin not only binds calcium but is regulated by a most representative Ca++-binding protein, CaM. Among the Ca++-binding proteins whose presence has been shown by immunohistochemistry in various retinal cells (ganglion and amacrine but, specifically, neither photoreceptor nor Müller cells) are CaM (Seto-Ohshima et al. 1987; Pochet et al. 1991), calbindin D-28k (Uesugi et al. 1992), parvalbumin (Endo et al. 1985), calretinin (Bastianelli et al. 1995), and neurocalcin (Nakano et al. 1992). Another Ca++-binding protein, recoverin, is exceptional because it is located mainly in photoreceptor and bipolar cells (Korf et al. 1992; Günhan-Agar et al. 2000). The location of most Ca++-binding proteins in ganglion cells and amacrine cells, which coincides with our localization of calcineurin, may indicate that ganglion cells and amacrine cells, but not Müller cells, are the main site of the function of calcium-binding activity in the retina.
The developmental changes of these proteins in the retina have been examined for CaM, calbindin D-28K, parvalbumin, protein kinase C, and CaM kinase IV. CaM appears in the embryonic mouse retina before the formation of neural layers, and is confined to the ganglion cells and cells in the inner nuclear layer during development (Seto-Ohshima et al. 1987). Calbindin D-28K appears in the neuroblastic layers of the embryonic rat retina, where its presence is confined to ganglion cells, amacrine cells, and horizontal cells during development (Uesugi et al. 1992). Parvalbumin is not present in the retina of rat embryo, rather appearing in weanling animals once the neural layers have formed (Oguni et al. 1998). Protein kinase C appears in rat retina at the latest 10 days after birth (Zhang and Yeh 1991), and CaM kinase IV appears in the ganglion cells of rat retina at the latest 3 days after birth (Sakagami and Kondo 1996), while bipolar cells also become positive for both protein kinase C and CaM kinase IV during development. The developmental changes of Ca++-binding proteins are very similar to those of calcineurin but quite different from those of protein kinase C and CaM kinase IV. The pattern of the appearance of Ca++-binding proteins and kinases regulated by Ca++ and/or CaM and the time course and kind of cells have similarities. Among these, the change in the localization of calcineurin that occurs with development was most similar to that of CaM, which regulates calcineurin.
The present study demonstrated the presence of calcineurin in mammalian retinas, especially in limited kinds of retinal cells. Despite the presence of calcineurin in the retina throughout development and the presence of calcineurin, together with other Ca++-binding proteins and Ca++/CaM-dependent kinases, in similar kinds of cells of the retina, information on this Ca++/CaM-dependent protein phosphorylation and dephosphorylation system in the retina is very limited. The physiological role of this major Ca++/CaM-dependent protein phosphatase in the retina, in morphogenesis, and in visual function needs to be elucidated.
Footnotes
Acknowledgments
We thank Dr Randall L. Kincaid (Department of Pharmacology, Pennsylvania State University College of Medicine) for kindly providing the calcineurin antibodies and Dr Yasuo Watanabe (Department of Pharmacology, Nagoya University) for helpful discussion.