
Abstract
Studies were undertaken to provide information regarding cell-specific expression of mucin genes in stomach and their relation to developmental and neoplastic patterns of epithelial cytodifferentiation. In situ hybridization was used to study mRNA expression of eight mucin genes (MUC1–4, MUC5AC, MUC5B, MUC6, MUC7) in stomach of 13 human embryos and fetuses (8–27 weeks' gestation), comparing these with normal, meta-plastic, and neoplastic adult tissues. These investigations have demonstrated that MUC1, MUC4, MUC5AC, MUC5B, and MUC6 are already expressed in the embryonic stomach at 8 weeks of gestation. MUC3 mRNA expression can be observed from 10.5 weeks of gestation. MUC2 is expressed at later stages, concomitant with mucous gland cytodifferentiation. Normal adult stomach is characterized by strong expression of MUC1, MUC5AC, and MUC6, less prominent MUC2, and sporadic MUC3 and MUC4, without MUC5B and MUC7. Intestinal metaplasia is characterized by an intestinal-type pattern with MUC2 and MUC3 mRNA expression. Gastric carcinomas exhibit altered mucin gene expression patterns with disappearance of MUC5AC and MUC6 mRNAs in some tumor glands, abnormal expression of MUC2, and reappearance of MUC5B mRNAs. In conclusion, we have observed that patterns of mucin gene expression in embryonic and fetal stomach could show similarities with some gastric carcinomas in adults. Differences in mucin gene expression in developmental, metaplastic, and neoplastic stomach compared to normal adult stomach suggest a possible regulatory role for their products in gastric epithelial cell proliferation and differentiation.
T
To date, at least eight mucin genes (MUC1–MUC4, MUC5AC, MUC5B, and MUC6–MUC7) have been identified and characterized from various normal or carcinoma tissues and epithelial cell-derived cell lines (Gendler and Spicer 1995; Van Klinken et al. 1995). The expression of these genes, as studied in adult, is tissue- and cell-specific according to a complex pattern, several genes being expressed in the same organ and sometimes in the same cell (Lesuffleur et al. 1994). Moreover, we have shown that the expression of the different mucin genes begins early in human development in the intestine (jejunum, ileum, colon) and respiratory tract, according to complex expression patterns differing from those previously observed in adults (Buisine et al. 1998, 1999). With the aim of providing further information regarding the tissue- and cell-specific expression of the mucin genes and their deregulation in epithelial cancers, we studied the spatio-temporal expression of the different mucin genes during embryonic and fetal development of the gastrointestinal tract and accessory digestive glands.
In this study, we examined the expression of all known mucin genes (MUC1–MUC4, MUC5AC, MUC5B, and MUC6–7) in the stomach of 13 human embryos and fetuses (aged 8–27 weeks of gestation) and correlated this with gene expression in normal, metaplastic, and neoplastic tissues in adults. Our results demonstrated that patterns of mucin gene expression in embryonic and fetal stomach may show similarities with some gastric carcinomas in adults.
Materials and Methods
Tissues
Stomach (cardia, fundus, and antrum) was obtained from four human embryos and nine human fetuses after spontaneous or therapeutic abortion with the approval of the local ethical committees. The specimens ranged in age from 8 to 27 weeks of gestation, dated from the last menstruation (data obtained from clinical records and confirmed by foot and crown–rump length). There was no evidence of congenital anomalies in the digestive tract of any of the specimens.
Samples of normal mucosa consisted of biopsy specimens taken from five patients undergoing endoscopy for epigastralgia. Biopsies were histologically normal and were used as controls. Additional specimens of macroscopically normal and tumor mucosae were obtained from five patients undergoing gastrectomy for gastric carcinoma.
Samples were obtained within 30 min of removal to avoid mRNA degradation and tissue lysis. They were immediately immersed in either fresh 4% paraformaldehyde or 10% phosphate-buffered formalin for in situ hybridization and further embedded in paraffin. Three-μm-thick sections were cut, mounted on gelatin-covered slides, and stored at 4C until used. Serial sections were systematically stained with hematoxylin–eosin–safran and astra blue for histological analysis.
Probes
In situ hybridization was performed using eight 35S-labeled oligonucleotide probes corresponding to each tandem repeat domain of MUC1, MUC2, MUC3, MUC4, MUC5AC, MUC5B, MUC6, and MUC7 as described in earlier studies (Buisine et al. 1998, 1999).
In Situ Hybridization
The hybridization steps were as described previously (Audié et al. 1993), with minor modifications. Briefly, tissue sections were deparaffinized, rehydrated, incubated with 2 μg/ml proteinase K (Boehringer Mannheim; Meylan, France) for 15 min, and fixed again in 4% paraformaldehyde in PBS for 15 min. Sections were then immersed in 0.1 M triethanolamine (Sigma; L'Isle d'Abeau Chesnes, France) containing 0.25% acetic anhydride for 10 min. Sections were prehybridized in 4 × SSPE, 1 × Denhardt's buffer for 45 min, and hybridized overnight at 42C in 20–100 μl of a 4 × SSPE solution containing 50% formamide (v/v), 0.1% N-lauroylsarcosine (w/v), 1.2 M sodium phosphate (pH 7.2), 1 × Denhardt's buffer, 3 mg/ml yeast tRNA, 20 mM dithiothreitol, and 7.5 × 103 dpm/μl of 35S-labeled oligonucleotide. After post-hybridization washes, slides were dipped in LM-1 emulsion (Amersham, Les Ulis, France), developed 2–3 weeks after exposure, and finally counterstained with methyl green pyronin (Sigma).
The following controls were performed: (a) tissue sections treated with 50 μg/ml ribonuclease A (Boehringer Mannheim); (b) tissue sections treated with a large excess of unlabeled oligonucleotide identical to or distinct from the 35S-labeled-probe; and (c) adult and fetal tissue sections were tested in parallel under the same conditions.
The intensity of the hybridization signal was scored semiquantitatively by two independent observers (MPB, LD) as: –, absent; +, weak (visible at magnification ×200); + +, moderate (visible at magnification ×100); + + +, strong (visible at magnification ×40).
Results
Hybridization data are summarized in Table 1.
Embryos and Fetuses
Mucin gene mRNAs were detected in all human embryonic and fetal specimens analyzed, from 8 to 27 weeks after gestation.
MUC5AC MUC5AC mRNAs were detected in all gastric specimens analyzed from 8 weeks of gestation. The labeling was largely restricted to the region of stomach destined to become the antrum. In the pseudostratified epithelium, the labeling was of moderate intensity, with a marked predominance in the upper third (Figure 1A and 1B). At 10.5 weeks, the signal remained stronger in epithelium of antrum, but epithelium of fundus was also labeled (Figure 1C). After 10.5 weeks of gestation, the labeling was homogeneous in gastric epithelium, irrespective of the region. Signal of moderate intensity was observed in all epithelial cells of the surface and pits (Figure 1D). After 18 weeks, developing glands could be easily distinguished from the crypts but remained unlabeled with the MUC5AC probe.
MUC6. MUC6 mRNAs were detected from 8 weeks of gestation in the embryonic stomach. The labeling was weak and homogeneous throughout the epithelium, irrespective of the region, until 10.5 weeks of gestation (Figure 1E). At 12 weeks, labeling was restricted to epithelial folds. After this time, MUC6 mRNAs were observed only in developing cardial and antral glands (Figure 1F).
MUC1. MUC1 mRNAs were detected from 8 weeks of gestation. The labeling was weak but homogeneously distributed throughout the cytoplasm of all epithelial cells of the surface and pits (Figure 1G). A weak signal was also observed in developing cardial and antral glands, whereas fundic glands remained unlabeled. The hybridization pattern remained identical from 8 weeks to 27 weeks.
MUC5B. MUC5B mRNAs were first detected at 8 weeks of gestation in the embryonic stomach, where the labeling was weak and homogeneous throughout the epithelium (Figure 1H and 1I). After this time, MUC5B mRNAs were inconsistently detected in stomach. When present, MUC5B expression was observed in the surface epithelium and epithelial folds (Figure 1J and 1K). At 27 weeks the signal in fundus was distributed in epithelial cells of the surface and pits, whereas it was restricted to epithelial folds and mucous glands in antrum.
MUC4. MUC4 mRNAs were detected from 8 weeks of gestation in the embryonic stomach, where the labeling was weak or moderate and heterogeneous throughout the epithelium (Figure 1L). After Week 12, MUC4 mRNAs were inconsistently found in stomach. When present, the labeling was homogeneous or heterogeneous, depending on the specimen and irrespective of the region.
MUC3. MUC3 mRNAs were first detected at 10.5 weeks of gestation in the stomach of one of the two fetal specimens of this age. The labeling was weak and was distributed throughout the epithelium, with a stronger signal in the region of stomach destined to become the antrum. After this time, MUC3 mRNAs were occasionally detected in stomach, where the labeling was limited to most of the surface epithelial cells.
MUC2. MUC2 mRNAs were detected only from Week 26 of gestation onward in fetal stomach, where the labeling was weak and limited to antral glands. MUC2 mRNAs were not detected in surface epithelium of stomach and fundic glands at any gestational age.
MUC7. MUC7 was not detected in embryonic or fetal stomach by in situ hybridization.
Adults
Normal Controls. Strong labeling was detected with the MUC5AC probe in epithelial cells of the surface and pits and in mucous neck cells, irrespective of the region (cardia, fundus, antrum) (Figure 2A). Strong labeling was also detected with the MUC6 probe, but the signal was restricted to neck cells and to cardial and antral glands (Figure 2B). A weak but homoge- nous signal was also observed with the MUC1 probe in epithelial cells of the surface and pits and in mucous glands. MUC3 and MUC4 mRNAs were inconsistently found in stomach, where the labeling was limited to the majority of surface epithelial cells. A weak signal was detected with the MUC2 probe in cardial and antral glands. Fundic glands remained unlabeled, whatever the probe. MUC5B and MUC7 mRNAs were never detected in normal adult stomach.
Intestinal Metaplasia. Intestinal metaplasia was observed in stomach of three out of the five patients with gastric carcinoma, in macroscopically normal mucosa. It was classified on the basis of histological criteria as complete intestinal metaplasia (small intestine phenotype), with non-mucin-secreting columnar cells and goblet cells. The MUC2 probe hybridized strongly to the perinuclear region of goblet cells (Figure 2C). The MUC3 probe labeled all epithelial cells homogeneously. The signal was present in both goblet and columnar cells. MUC4 mRNAs were detected in one case, in goblet and columnar cells. MUC1, MUC5AC, MUC5B, MUC6, and MUC7 were not expressed in intestinal metaplasia.
Gastric Carcinomas. Mucin gene expression was analyzed in five gastric adenocarcinomas, four of which were well-differentiated. The hybridization pattern varied greatly among the specimens and within tumor glands of a given specimen (Table 1). MUC1 and MUC4 mRNAs were detected in all specimens analyzed (Figure 2D). MUC3 mRNAs were detected in most of the specimens (4/5). MUC5AC mRNAs were detected in association with MUC6 mRNAs in 4/5 gastric adenocarcinomas, and MUC2 mRNAs in 3/5 gastric adenocarcinomas (Figure 2E). MUC5B mRNAs were also detected in 3/5 specimens (Figure 2F). In 2/3 cases, MUC5B mRNA expression was associated with MUC5AC in the same tumor glands. In contrast, within a given specimen, there was not any overlap between cellular localization of MUC2 mRNA expression and MUC5AC or MUC5B mRNA expression.
Discussion
Gastric mucins are widely assumed to play important cytoprotective roles. They protect the epithelium against acid and pepsin in the gastric juice and against exogenous damaging agents (e.g., pathogens, drugs), and against mechanical damage (Bell et al. 1985; Allen et al. 1986).
Major changes in the normal pattern of mucin expression, quantitatively and/or qualitatively, have been described in various preneoplastic and neoplastic tissues. There is evidence that mucins play diverse roles in addition to protective functions. More particularly, mucins may be implicated in progression of human carcinomas and in promotion of tumor cell metastasis (Hilkens et al. 1995; Ho and Kim 1995). Mucins may also be implicated in cellular adhesion during organogenesis (Braga et al. 1992).
Mucin gene expression in embryonic, fetal, and adult stomach a
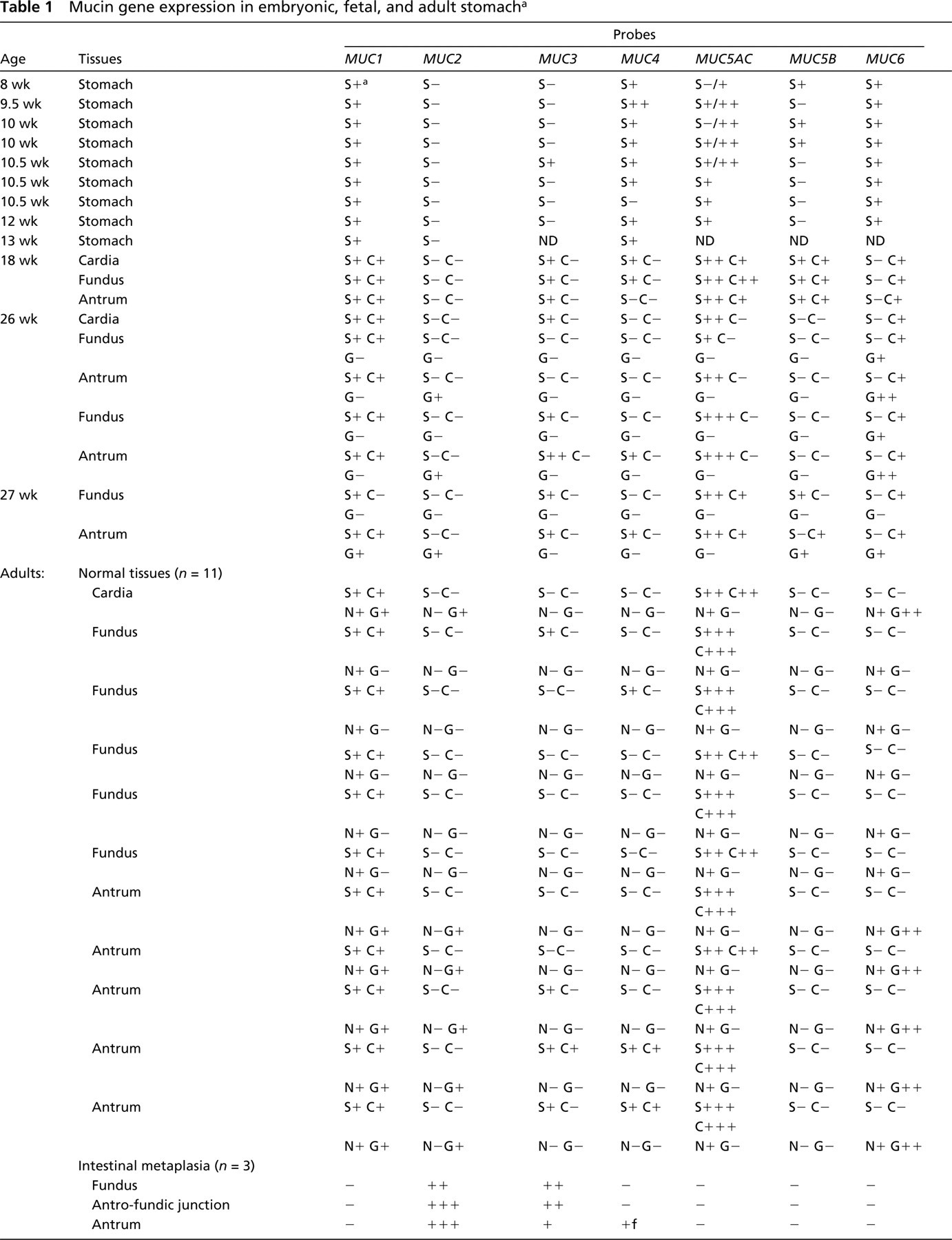
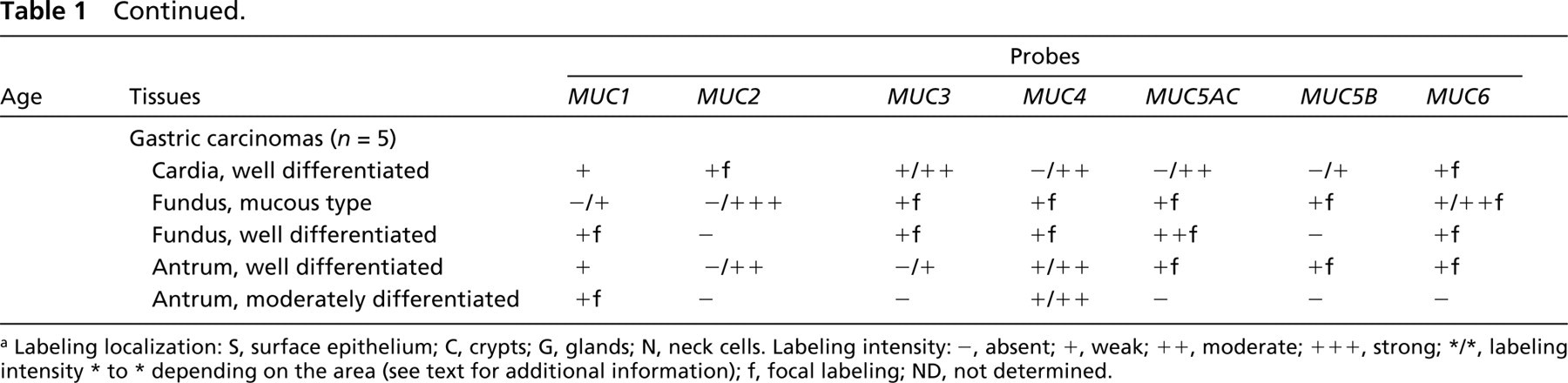
a Labeling localization: S, surface epithelium; C, crypts; G, glands; N, neck cells. Labeling intensity: –, absent; +, weak; ++, moderate; +++, strong; */*, labeling intensity * to * depending on the area (see text for additional information); f, focal labeling; ND, not determined.
Transformed cells frequently transcribe genes that are expressed during embryonic and fetal stages. A number of mucin carbohydrate antigens, including blood group antigens Tn, T, and sialyl-Tn antigens, have been demonstrated to be “oncofetal antigens” in stomach (Lesuffleur et al. 1994; Kim et al. 1996). Less is known concerning apomucins.
In this study, we have characterized mucin gene expression during human fetal development of the stomach and compared this with gene expression in normal, metaplastic, and neoplastic tissues in adults. Using in situ hybridization, we have examined the expression of all presently characterized mucin genes (MUC1–MUC4, MUC5AC, MUC5B, MUC6–7) in 13 human embryos and fetuses (aged 8–27 weeks of gestation). To our knowledge, the only studies describing developmental expression of gastric mucin genes were performed by Reid and Harris (1998), who analyzed mucin gene expression in one 23-week-old fetus, and Bartman et al. (1998), who analyzed the expression of MUC6 in nine fetuses aged 16–36 weeks.
Synthesis of mucus begins late in fetal stomach by 12 weeks of gestation, some 2 weeks after intestinal mucosae (Stauffer et al. 1990). Our results show that mucin gene expression occurs earlier, with MUC1, MUC4, MUC5AC, MUC5B, and MUC6 being already expressed at 8 weeks of gestation, which corresponds to the youngest specimen analyzed. At this stage, the gastric epithelium is poorly differentiated. The rudiments of gastric pits develop by 9–10 weeks of gestation in the fundus and later in development in the antral area and cardia around 12 weeks. Mucous glands appear at the gestational age of 13–14 weeks (Deren 1971; Grand et al. 1976). Thus, MUC1, MUC4, MUC5AC, MUC5B, and MUC6 are expressed in fetal stomach before epithelial cytodifferentiation occurs. However, MUC6 apomucin can be detected only late in development in stomach by immunohistochemistry, from 20 weeks of gestation (Bartman et al. 1998), suggesting that epitopes may be masked in embryos and younger fetuses. Likewise, we have reported previously that MUC1, MUC2, and MUC4 are expressed in embryonic lung before epithelial cytodifferentiation into ciliated or secretory cells, with MUC4 expression as early as 6.5 weeks of gestation in the foregut and MUC1 and MUC2 expression from 9–10 weeks of gestation in trachea and developing bronchi (Buisine et al. 1999). In the primitive intestine, MUC4 is, along with MUC3, the first mucin gene to be expressed at 6.5 weeks of gestation, MUC5AC being expressed from 8 weeks and MUC2 from 9 weeks of gestation (Buisine et al. 1998). Thus, each mucin gene exhibits a complex spatio–temporal expression pattern that is quite different in developing stomach, airways, and intestine. Taken together, our results demonstrate distinct regulation of the activation of the different mucin genes during morphogenesis and epithelial cytodifferentiation of the stomach compared to the developing respiratory tract and intestine, suggesting specific roles for them in each epithelial organ and the existence of complex mechanisms of tissue-specific regulation during ontogenesis. Moreover, we have shown that undifferentiated embryonic gastric epithelial cells have the multipotential capability to express five or six mucin genes (MUC1, MUC4, MUC5AC, MUC5B, MUC6 and, in some cases, MUC3), when later in development and in adults there appears to be lineage restriction, such that epithelial cells of the surface epithelium express only two to four genes (MUC1, MUC5AC and, in some cases, MUC3 and MUC4) and epithelial cells of the mucous gland only three genes (MUC1, MUC2, and MUC6). This observation shows that a concerted regulation between the mucin genes probably occurs during ontogenesis according to very complex mechanisms that lead to the co-expression of a wide variety of mucin genes in the different organs.
In this study, we have confirmed that MUC5AC is, along with MUC6, the major mucin gene expressed in adult stomach, in which MUC5AC is expressed in all epithelial cells of surface and pits, whereas MUC6 is expressed in mucous neck cells and in cardial and antral cells (Audié et al. 1993; Bartman et al. 1998). Moreover, we have shown that MUC5AC and MUC6 are the major mucin genes expressed in embryonic and fetal stomach. Although cytodifferentiation is known to proceed peripherally from the fundus (Deren 1971; Grand et al. 1976; deSa 1991), MUC5AC is expressed first in the region of stomach destined to become the antrum. However, similarly, using histochemistry, Stauffer et al. (1990) demonstrated that secretion of mucins initiates first in the antral mucosa (at 12 weeks of gestation) and 1 week later in fundic mucosa.
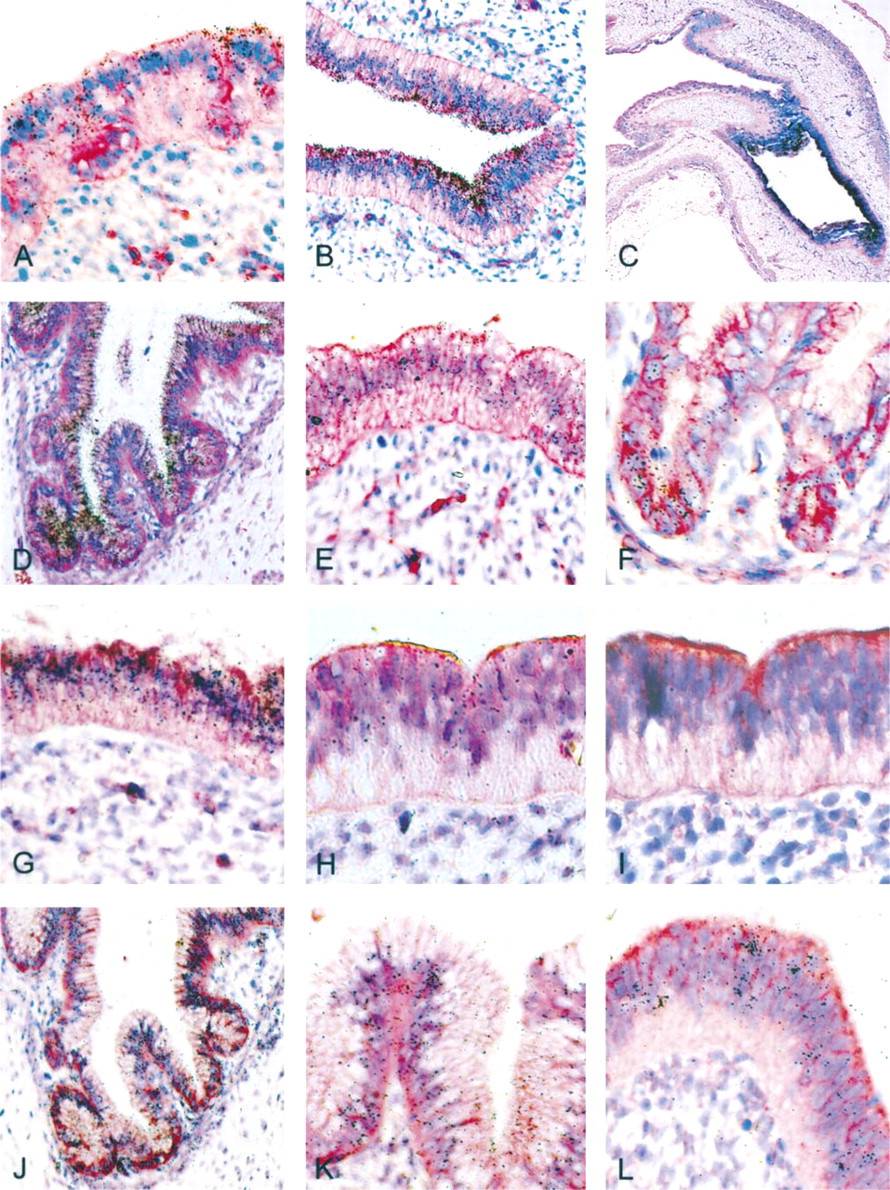
In situ hybridization for mucin gene mRNAs in embryonic and fetal stomach. (
MUC6 expression in the developing stomach is quite different from that observed in normal adult gastric mucosae. We have shown that MUC6 mRNAs are present first in all surface epithelial cells. After this time, MUC6 expression is restricted to epithelial folds and then to the developing mucous glands. This observation suggests that undifferentiated epithelial cells of the surface epithelium have the capacity to express MUC6 long before some of them migrate down into the mesenchyme, acquiring their cytodifferentiation into mucous gland cells (Deren 1971; Grand et al. 1976; deSa 1991). This pattern of MUC6 expression in the developing stomach is very close to that we have previously observed for MUC5B in the developing respiratory tract, where it is first expressed in epithelial cells of the surface epithelium and is restricted to submucosal glands and gland ducts in older fetuses (Buisine et al. 1999). Therefore, MUC6 may play a role in regulating the formation of gastric mucous glands. In contrast, MUC2 expression initiates late in fetal stomach, at 26 weeks of gestation, when it is associated with the presence of differentiated mucous glands. Therefore, MUC2 expression does not seem to be important for gastric epithelium morphogenesis and cytodifferentiation. Recently, MUC6 has been shown to be aberrantly expressed in the surface epithelium of stomach of Helicobacter pylori-infected patients (Byrd et al. 1997). H. pylori infection is characterized by an increase of epithelial proliferation which tends to expend towards the surface epithelium (Panella et al. 1996). Our present result suggests that the pattern of MUC6 mRNA expression in H. pylori-infected patients is reminiscent of that observed in embryonic stomach and may reflect excessive epithelial cell turnover.
MUC5B mRNAs are absent from normal adult stomach. Interestingly, we have shown that MUC5B is expressed in embryonic and fetal stomach from 8 to 27 weeks of gestation, in both antrum and fundus. Therefore, MUC5B expression is developmentally regulated during gastric ontogenesis. The persistence of MUC5B mRNA expression until near term is not surprising because differentiation of gastric mucosae is very slow compared to that of intestinal mucosae, complete maturation being attained only 3–4 years after birth (Stauffer et al. 1990).
Intestinal metaplasia is frequently associated with gastric carcinoma (Correa and Shiao 1994). In the present study, intestinal metaplasia was present in the stomach of three out of the five patients with gastric carcinoma. Only MUC2, MUC3 and, to a lesser extent, MUC4, were expressed in intestinal metaplasia, where MUC2 was associated with goblet cells and MUC3 and MUC4 with both goblet and columnar cells. This hybridization pattern corresponds to that observed in normal small intestine and is therefore consistent with the small intestine-type phenotype of intestinal metaplasia according to the classification of Baracchini et al. (1991). These data confirm previous observations of mucin gene expression in intestinal metaplasia (Ho et al. 1995; Reis et al. 1999). Therefore, according to the pattern of mucin gene expression, intestinal metaplasia appears to correspond to an aberrant lineage and is not reminiscent of an embryonic or fetal phenotype. Whatever their location in normal states (in surface epithelium of the respiratory tract, small intestine, and colon) or in pathological states (in stomach), cells with goblet cell phenotype are always associated with MUC2 mRNA expression. Interestingly, Gum et al. (1999) have recently demonstrated that the MUC2 gene promoter contains elements sufficient for the appropriate expression of MUC2 in small intestinal goblet cells.
Very few data are available regarding mucin gene expression in preneoplastic and neoplastic gastric epi- thelium. In the present study, we have shown that gastric carcinomas exhibit marked altered patterns of mucin gene expression compared with normal mucosa, with increased levels of MUC2, MUC3, and MUC4 mRNAs and decreased levels of MUC5AC and MUC6 mRNAs in some tumor glands. These findings are in accordance with those obtained by Ho et al. (1995) using Northern blotting analysis. Moreover, we have shown expression of MUC5B mRNAs in gastric carcinomas (3/5), confirming the notion that genes preferentially expressed in developmental stomach are expressed in gastric cancers (Tanaka et al. 1992). Further investigations concerning the expression of the mucin genes, especially the MUC5B gene, in gastric cancers should be performed to complete the present study. Such investigations might contribute to new insights in classification and histogenesis of gastric tumors.
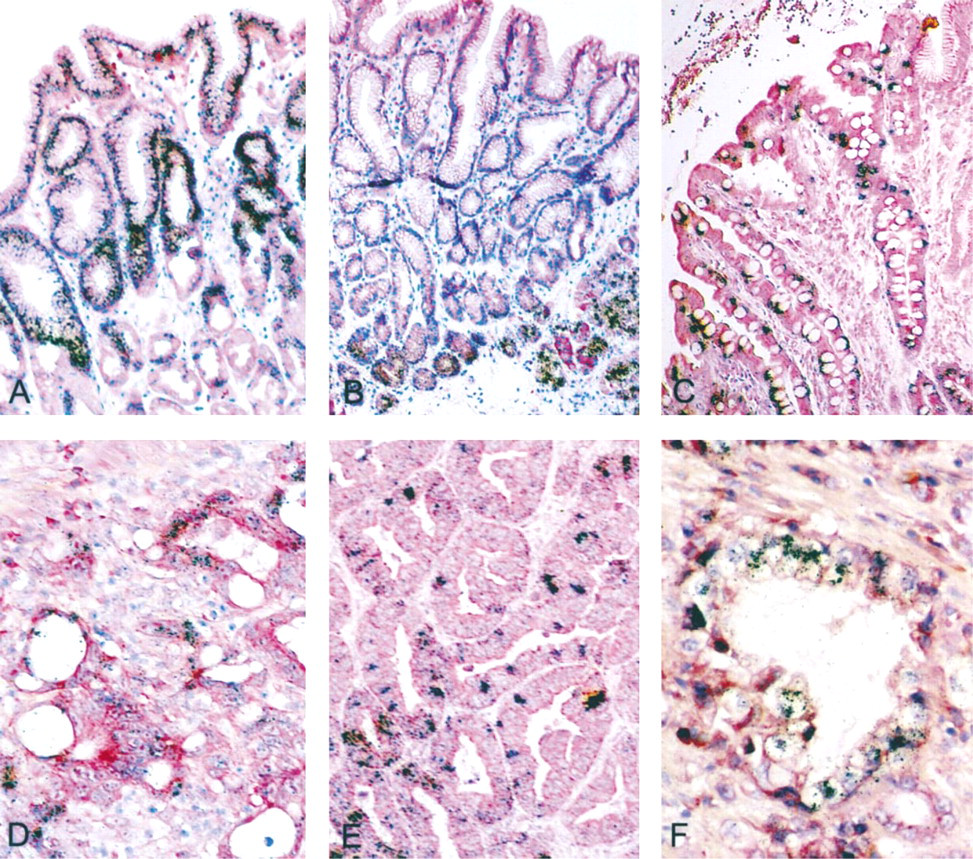
In situ hybridization for mucin gene mRNAs in adult stomach. (
In summary, these investigations have demonstrated that MUC1, MUC4, MUC5AC, MUC5B, and MUC6 are already expressed in the embryonic stomach at 8 weeks of gestation. In contrast, MUC2 is expressed at later stages, concomitant with mucous gland cytodifferentiation. Intestinal metaplasia is characterized by an intestine-type pattern, with MUC2, MUC3, and sometimes MUC4 mRNA expression. Gastric carcinomas exhibit altered mucin gene expression patterns, with disappearance of MUC5AC and MUC6 mRNAs in some tumor glands, overexpression of MUC2, and reappearance of MUC5B mRNAs. Promoter sequences of some of the mucin genes (MUC1, MUC2, MUC5AC, and MUC5B) are now available (Kovarik et al. 1993; Gum et al. 1997; Li et al. 1998; Van Seuningen et al. 2000). Elucidation of their regulatory mechanisms will provide insights into the molecular bases of their tissue- and cell-specific expression in normal adults as well as during normal developmental processes and tumorigenesis.
Footnotes
Acknowledgements
Supported by grants from the Association pour la Recherche sur le Cancer (contract no. 9643), the EC Concerted Action: Mucins in inflammatory disease, and the CHRU de Lille (contract no. 96/29/9595).
We thank Marie-Claire Dieu for excellent technical assistance and ICARE Multimédi@, particularly Gérard Espouy, for color prints.