
Abstract
Alpha-1 microglobulin (α1m), a lipocalin, is an evolutionarily conserved immunomodulatory plasma protein. In all species studied, α1m is synthesized by hepatocytes and catabolized in the renal proximal tubular cells. α1m deficiency has not been reported in any species, suggesting that its absence is lethal and indicating an important physiological role for this protein To clarify its functional role, tissue distribution studies are crucial. Such studies in humans have been restricted largely to adult fresh/frozen tissue. Formalin-fixed, paraffin-embedded multi-organ block tissue from aborted fetuses (gestational age range 7–22 weeks) was immunohistochemically examined for α1m reactivity. Moderate to strong reactivity was seen at all ages in hepatocytes, renal proximal tubule cells, and a subset of pancreatic islet cells. Muscle (cardiac, skeletal, or smooth), adrenal cortex, a scattered subset of intestinal mucosal cells, tips of small intestinal villi, and Leydig cells showed weaker and/or variable levels of reactivity. Connective tissue stained with variable location and intensity. The following cells/sites were consistently negative: thymus, spleen, hematopoietic cells, lung parenchyma, glomeruli, exocrine pancreas, epidermis, cartilage/bone, ovary, seminiferous tubules, epididymis, thyroid, and parathyroid. The results underscore the dominant role of liver and kidney in fetal α1m metabolism and provide a framework for understanding the functional role of this immunoregulatory protein.
A
In addition to its presence in mammals and birds (Åkerström 1985), an α1m gene has been identified in fish and amphibians (Hanley and Powell 1994; Leaver et al. 1994; Kawahara et al. 1997). The protein has been purified from plaice and was surprisingly similar to mammalian α1m (Lindqvist and Åkerström 1999). This evolutionary conservation suggests an important physiological role that remains undefined despite more than 25 years of investigation. Thus far, α1m deficiency has not been reported in any species, suggesting that its absence is lethal. Like other lipocalins, such as the retinol-binding protein, α1m may have a plasma transport function and might carry a hydrophobic ligand (Åkerström and Lögdberg 1990). Additional functions could be associated with the various complexed forms of the protein but remain to be defined. Of note, a series of in vitro studies has shown that α1m has immunomodulatory properties, including inhibition of antigen-stimulated lymphocyte and of granulocyte functions (Mendez et al. 1980; Åkerström and Lögdberg 1990; Wester et al. 1998). There is also evidence that the α1m gene, present in the lipocalin cluster on chromosome 9 (q32–33) (Diarra-Mehrpour et al. 1989; Traboni et al. 1989) is developmentally regulated. For example, a progressive increase in transcription rate in mouse liver, peaking at Day 5 after birth, was reported (Salier et al. 1993). In addition, thyroid hormone causes repression of α1m gene expression during amphibian metamorphosis (Kawahara et al. 1997). These data suggest an important functional role for α1m during growth and development.
Many studies of α1m in humans have measured its concentration in serum under a variety of normal and pathological conditions, or in urine as a marker of kidney pathology (Itoh and Kawai 1990). Impaired kidney function affects α1m levels in serum (increasing with reduced glomerular filtration rate) and urine (increasing with proximal tubule damage). However, levels of α1m in biological fluids are not markedly influenced by other pathological conditions (Itoh and Kawai 1990). Less is known about the normal variability of local α1m concentrations throughout the body and about changes in the relative distribution of α1m among its diverse complexes in health and disease.
To begin to clarify these issues, tissue distribution studies of the protein are crucial. Several biochemical and immunohistochemical studies have been published but are limited mainly to adult fresh or frozen tissue (Åkerström 1983; Bouic et al. 1984a,b,1985). These studies all showed the presence of α1m in hepatocytes (the primary site of synthesis) and in proximal renal tubules (the major site of catabolism).
Because of the limited supply of fresh and frozen tissue, little is known about the developmental expression of α1m in humans. The recent development (Odum and Nielsen 1994; Berggård et al. 1998, 1999b) of immunohistochemical methods for detection of α1m in formalin-fixed tissue and the enhanced detection with an antigen retrieval technique (unpublished observations) paved the way for investigating archival material. Using these methods, we have systematically explored the localization of α1m in the human fetus at various stages of development. These data will bear on the evolving understanding of the physiological and pathological role of α1m.
Materials and Methods
Tissues
Archival, formalin-fixed fetal tissues were obtained from 22 aborted fetuses (age range 7–22 weeks estimated gestational age) after approval by the Montefiore Medical Center Institutional Review Board. These fetuses were the subjects of surgical pathology review, and special care had been taken with the diagnostic tissue blocks to include representative tissues from several organs from each fetus, sampled by a 3-mm diameter skin punch. Organs sampled included liver, kidney, pancreas, heart, skeletal muscle, lung, esophagus, stomach, small and large intestine, bone/cartilage, skin, testis, ovary, thymus, spleen, adrenal, and thyroid. Multiple tissue samples (10–15) from an individual were placed in one cassette and processed according to standard protocol to generate paraffin blocks. Tissue sections for immunohistochemistry were cut at 4-μm thickness.
Antibodies
Chromatographically purified human urinary α1m (Åkerström et al. 1995) was injected into rabbits to generate anti-human α1m antibodies (Ekström and Berggård 1977). Binding properties of all antisera were analyzed by radioimmunoassay (Åkerström 1985) and solid-phase radioimmunoassay (Nilson et al. 1986). Similar titers and inhibition curves for the binding of purified human urinary α1m were obtained. The binding specificities were analyzed by Western blotting (Fredriksson et al. 1987), separating human plasma by SDS-PAGE under reducing conditions. All antibodies bound exclusively to free monomeric α1m (31 kD), IgA- α1m (90 kD, 180 kD), and albumin– α1m (120 kD). IgG from the rabbit sera was purified by anion exchange chromatography as described (Nilson et al. 1986). These purified IgG preparations were routinely used for staining. Anti- α1m IgG, exhaustively adsorbed on immobilized, purified urinary human α1m (3 mg α1m:1 ml CNBr-activated Sepharose; Pharmacia Fine Biochemicals, Uppsala, Sweden), served as a negative control in these experiments. In additional control experiments, we also employed anti-α1m IgG similarly adsorbed on immobilized, purified recombinant human α1m (Wester et al. 1997). These absorbed antibodies no longer showed anti-α1m activity in radioimmunoassays or on Western blots. In initial experiments, a monoclonal mouse anti-α1m, BN11.10 (Nilson et al. 1987), was also tested. The fetal material was completely analyzed with antibodies from one rabbit (K322), and selectively reanalyzed with antibodies pooled from four different rabbits (K406–9). Staining of pancreas was further investigated with a panel composed of six additional anti-α1m antibodies/antisera (K107, K108, K109, K201, K202, and K319+323).
Immunohistochemistry for α1m
α1m was readily and specifically detected by immunohistochemistry on formalin-fixed, paraffin-embedded tissues using polyclonal antibodies, with improvement in staining after antigen retrieval. In contrast, the monoclonal anti-α1m antibodies (BN11.10) failed to detect the protein under the same conditions. Therefore, polyclonal antibodies and antigen retrieval on tissue sections were employed throughout this investigation. The immunohistochemical assay used the avidin–biotin–peroxidase-method with diaminobenzidine (DAB) as the substrate (LSAB kit; DAKO, Carpinteria, CA). Sections were counterstained with hematoxylin. Briefly, 4-μm-thick sections were stained with anti-α1m IgG, in most experiments at a final concentration of 10 μg/ml (K322), at 100 μl/slide (30-min incubation at room temperature in a humid chamber). Before the staining, after blocking of endogenous peroxidase with H2O2 (15-min incubation in a 37C water bath), the tissue sections had been heat-reduced (by microwave) for 5 min in citrate buffer, pH 6.0, for antigen retrieval. Antigen retrieval conditions and antibody dilutions were standardized using tissue sections from normal adult kidney (tubules positive; glomeruli negative) and from a liver (hepatocytes positive) with metastatic colon adenocarcinoma (tumor negative), exploiting the high concentrations of α1m in adult human kidney and liver. Kinetic studies showed that near-complete antigen retrieval was achieved as early as after 5 min of microwave heating. Staining was scored as strong (3 +), moderate (2 +), weak (1+), or absent (0) by two independent observers.
Results
α1m Distribution in Human Fetal Tissues: Overview
α1m staining was found in organs at all ages studied (7–22 gestational weeks). As in adult tissues of many species, particularly strong staining was seen in the proximal tubules of the evolving kidney (Figure 1a), and cytoplasmic staining of fetal hepatocytes was also consistently present (Figure 1d). As detailed below, α1m staining was found at a number of other sites.
In all cases, staining for α1m was specific and was not seen with anti-α1m antibodies that had been exhaustively absorbed against purified α1m, as demonstrated for kidney in Figure 1b. Table 1 summarizes the localization of α1m in fetal tissues using anti-α1m K322 on tissue from 22 cases. In addition to hepatocytes and renal proximal tubules cells, moderate anti-α1m staining of a subset of pancreatic islet cells (Figure 1c) was observed in most cases. Other sites with weaker and/or variable staining included the tips of small intestinal villi (Figure 1e), a scattered subset of intestinal mucosal cells (Figure 1f), cardiac muscle (Figure 1g), skeletal muscle (Figure 1h), smooth muscle, adrenal cortex, and testicular Leydig cells. In addition, the perivascular connective tissue was variably positive in most organs throughout the fetal ages studied.
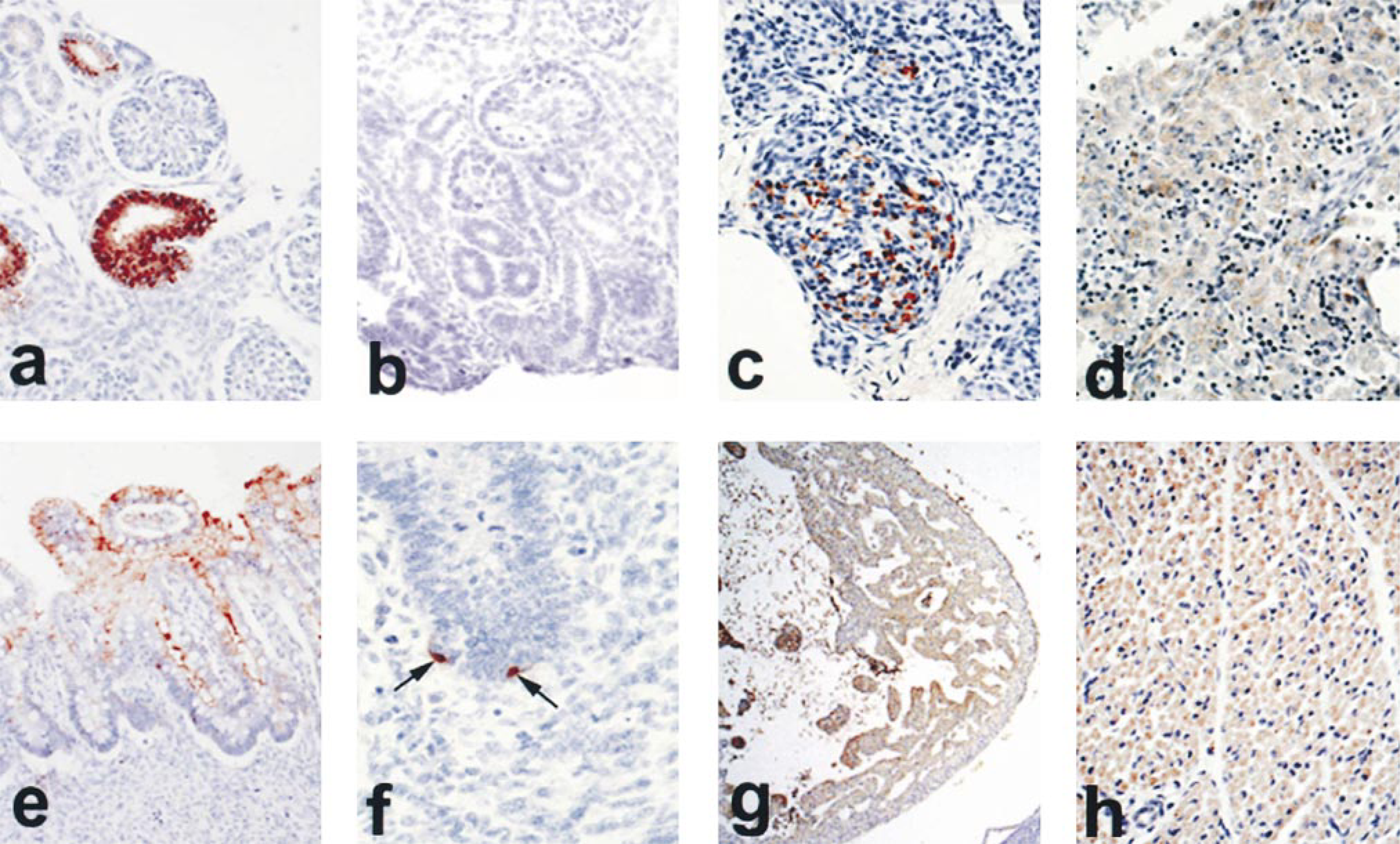
Immunohistochemical visualization of α1m in various fetal tissues. (
Organ-specific distribution of α1m expression
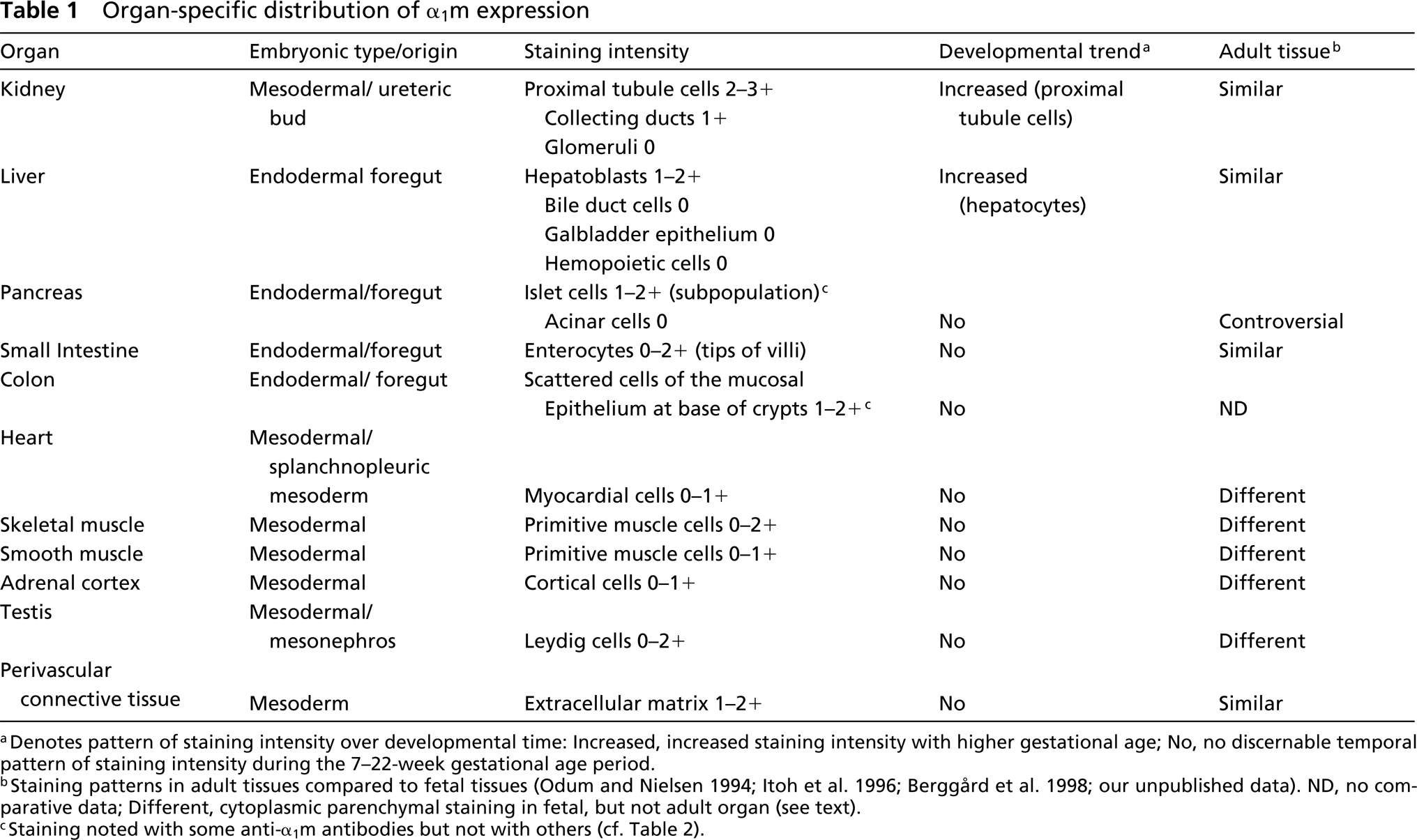
aDenotes pattern of staining intensity over developmental time: Increased, increased staining intensity with higher gestational age; No, no discernable temporal pattern of staining intensity during the 7–22-week gestational age period.
bStaining patterns in adult tissues compared to fetal tissues (Odum and Nielsen 1994; Itoh et al. 1996; Berggård et al. 1998; our unpublished data). ND, no comparative data; Different, cytoplasmic parenchymal staining in fetal, but not adult organ (see text).
cStaining noted with some anti-α1m antibodies but not with others (cf. Table 2).
The following cells/sites were consistently negative: thymus, spleen, hematopoietic cells (Figure 1d), lung, glomeruli (Figure 1a), exocrine pancreas (Figure 1c), epidermis, cartilage/bone, ovary, seminiferous tubules, epididymis, thyroid, and parathyroid. It is noteworthy that α1m reactivity is not restricted to tissues originating from one germ layer, because cells from both endoderm (liver, intestines) and mesoderm (testis, muscle) appear to express α1m during one or more stages of fetal development.
α1m Distribution in Human Fetal Tissues: Analysis by Organ System
Gastrointestinal System. There was no staining for α1m in the superficial squamous epithelium of the oral cavity, pharynx, or esophagus at any age. In the stomach, the parietal cells showed positive staining but the columnar cells of the surface epithelium and the neck glands were negative. The small intestine showed granular reactivity within the cytoplasm of the surface absorptive cells at the tips of the intestinal villi (Figure 1e), beginning at about the 14th week of gestational age. The intensity of this staining and the number of reactive villous cells increased in number with further development, but goblet cells and crypts were consistently negative. Moderate-intensity positive staining was seen in scattered individual cells located at the base of the colon crypts as early as 7 weeks of gestational age (Figure 1f). These cells had the morphological characteristics of neuroendocrine cells. All the other cells of the colon surface and glandular epithelium were negative at all ages.
At the age of 7 weeks, there was moderately intense α1m staining of a few hepatocytes scattered throughout the liver, with low-intensity staining of more than 50% of the hepatocytes. No staining of the extrahepatic bile duct was seen at this age. With development of the portal triads, reactivity for α1m was seen in all the hepatocytes (Figure 1d), with slightly stronger staining of the periportal hepatocytes. This pattern of staining persisted even after development of the ductal plates. In the region of the portal triads, staining was seen in the periportal hepatocytes, ductulocanalicular cells, and “oval cells.” The bile duct epithelium and the hematopoietic cells did not react with the anti-α1m antibodies at any age.
In the pancreas (see also below), moderate and occasionally strong α1m staining was seen in a subset of the cells in the islets of Langerhans (Figure 1c). These moderately staining cells were located mainly at the periphery of the islets but were also scattered within the center of the islets. The distribution of these cells did not follow that expected of any of the known subpopulations of islet cells. The rest of the cells in the islets showed very weak intensity staining or, more commonly, were unstained. Pancreatic acini and the ductule epithelium did not exhibit significant staining.
Genitourinary System. α1m staining in the kidney was restricted to the convoluted tubules and was strongly intense in this site (Figure 1a) and weakly intense in the collecting ducts. The glomerular tuft and Bowman's capsule were consistently negative (Figure 1a). There was moderately increased staining of the proximal tubule cells with increasing age. Weak staining of the interstitial Leydig cells of the testis was seen. No significant staining was seen in the seminiferous tubules or the rete testis.
Cardiovascular System. α1m staining of the myocardium was evident as early as 7 to 8 weeks of gestation (Figure 1g). This persisted with development. The endocardium and pericardium were negative. A variable-intensity staining of the intima and media of the medium-sized blood vessels was seen.
Respiratory System. There was no significant α1m staining of the bronchial epithelium or the lung in fetuses below the age of 15–16 weeks. In some cases, with development of the acini, the alveolar epithelium showed weak surface staining of the luminal cytoplasmic membrane. Staining of the pleura was not evident.
Reticuloendothelial System. No α1m staining was seen in the thymus, lymph nodes, spleen, or bone marrow.
Endocrine System. Diffuse weak-intensity cytoplasmic α1m staining was seen in the deeper layers of the adrenal cortex. No significant staining was seen in the thyroid or parathyroid tissues examined.
Musculoskeletal System. There was no α1m staining of the skin or the cutaneous appendages. Positive staining of weak to moderate intensity was seen in skeletal (Figure 1h) and smooth muscle at all ages of development. Although strong expression of α1m was seen in the primitive mesenchyme, the bone and cartilage did not react with the antibody. Perivascular connective tissue and connective tissue at the hilus of the organs, such as the porta hepatis, showed moderate expression. There was no staining of the basement membranes of the epithelia or the collagen in the skin or bone.
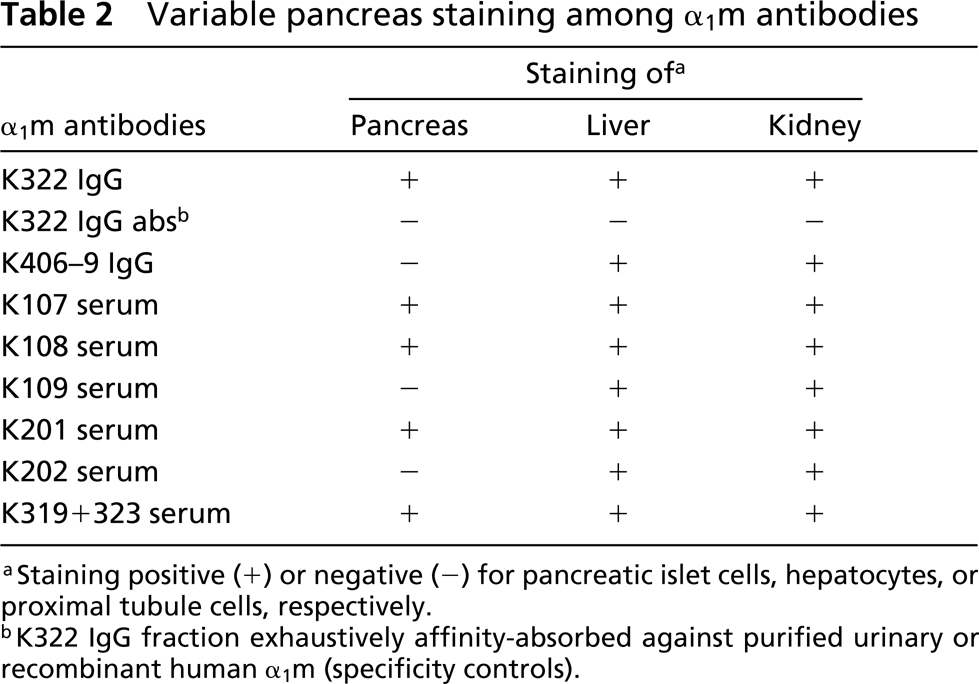
Staining of Pancreatic Islet Cells and Scattered Neuroendocrine-like Cells of the Large Intestine Was Restricted to Some α1m Antibodies and Was Not Seen with Others
When some of the staining experiments were repeated with a different source of anti-α1m antibodies (anti-α1m K406–9), we noted that the majority of tissues exhibited staining patterns identical to those previously observed with anti-α1m K322. However, the anti-α1m K406–9 antibodies stained neither the pancreatic islet cells nor the gastrointestinal “neuroendocrine” cells. Testing of additional antibodies/antisera raised against different preparations of purified human α1m revealed that some (5/8) lots were pancreas-reactive, whereas other lots (3/8) were nonreactive (Table 2). In these experiments, reactivity to kidney or liver was uniformly moderate to strong for all the antibodies/antisera. It was noted that adsorption of anti-α1m K322, not only with purified urinary α1m but, alternatively, with purified recombinant human α1m, completely eliminated its ability to stain the various fetal tissues, including pancreatic islet cells and gastrointestinal “neuroendocrine” cells.
Discussion
The primary site of synthesis of α1m in the adult is the liver, where it is translated as a diprotein precursor with bikunin, a Kunitz-type proteinase inhibitor and subunit of inter-α-inhibitor (Salier et al. 1996). The α1m–bikunin gene is divided into two separate clusters of exons, indicating an origin from independent lipocalin and Kunitz-type ancestral genes, respectively. This tandem arrangement is conserved among mammals and fishes (Hanley and Powell 1994; Leaver et al. 1994), suggesting that the fusion of the ancestral genes took place at least 450 million years ago. Therefore, the co-synthesis of these proteins may confer a significant functional advantage to the organism. The diprotein is cleaved into bikunin and α1m before they are released from the hepatocytes as separate proteins (Bratt et al. 1993). The major site of catabolism of α1m is the proximal convoluted tubules of the kidney.
The synthesis and/or expression pattern of α1m in other organs has not been well studied. This is largely due to the fact that tissue distribution studies have mostly been limited to fresh and frozen tissue, and conflicting data were reported (Åkerström 1983; Bouic et al. 1984a,b, 1985). We have recently found α1m to be readily and specifically detected on formalin-fixed, paraffin-embedded tissues by the use of polyclonal anti-α1m antibodies (Berggård et al. 1998, 1999b) and to be enhanced after antigen retrieval (unpublished data). Our study, for the first time, attempts to clarify the localization of α1m in the human fetus in a comprehensive manner and to elucidate its developmental expression, using a multi-organ block arrangement that allows efficient screening of large numbers of tissues.
Variable pancreas staining among α1m antibodies
aStaining positive (+) or negative (–) for pancreatic islet cells, hepatocytes, or proximal tubule cells, respectively.
bK322 IgG fraction exhaustively affinity-absorbed against purified urinary or recombinant human α1m (specificity controls).
α1m staining was found in fetal liver and proximal convoluted tubules of the kidney in all cases and at all developmental stages (our earliest stage was Week 7 of gestation). This suggests a dominant role for these organs in the metabolism of α1m in fetal as in adult life. The dominant role of the liver in the synthesis of α1m suggests that α1m expression may be useful as an indicator of hepatocyte differentiation. Our preliminary data demonstrate that α1m can be employed as an immunohistochemical marker to aid in distinguishing primary from secondary liver tumors (unpublished observations).
In certain other organs (cardiac, skeletal, or smooth muscle; adrenal cortex; testicular Leydig cells), specific cytoplasmic staining for α1m was noted in most cases. In the adult, using the same staining protocol, α1m is essentially absent from the parenchyma at these sites (own unpublished observations; see Table 1) and its synthesis cannot be demonstrated in heart, skeletal muscle, or adrenal gland by RT-PCR (Berggård et al. 1998). Therefore, α1m may exert functions specific to the fetus in these tissues. Staining of these organs was generally weaker than that of hepatocytes and proximal tubule cells, and a clear developmental trend in staining intensity, could not be ascertained for the time period studied (7–22 weeks).
The presence of α1m in fetal intestinal villi may correspond to the weak staining by anti-α1m in adult rat intestine (Bouic et al. 1984b). Using the same staining protocol, we also stained adult human small intestine (own unpublished observations). α1m does not appear to be synthesized at this site in the adult, as determined by RT-PCR (Itoh et al. 1996). These observations raise the issue of synthesis vs transport in the body, i.e., where is α1m present as a result of local synthesis and where is it present due to uptake/deposition of circulating α1m? For example, the intestinal α1m could originate from fetal swallowing of amniotic fluid, which contains the protein. Additional questions concern the extent to which putative α1m receptors (Fernandez-Luna et al. 1988; Babiker–Mohamed et al. 1990a; Wester et al. 1998) play a role in such uptake. In the intestinal mucosa, the presence of α1m may reflect a specialized functional involvement of the molecule, possibly by providing immunoregulation as a component of the gut-related immune system.
Significant synthesis of α1m has not been demonstrated in adult human tissues other than liver and possibly pancreas (Itoh et al. 1996; Berggård et al. 1998). Therefore, as discussed above for the intestinal mucosa, it is likely that the presence of α1m at some of the other tissue sites, notably the perivascular extracellular matrix in adults (Odum and Nielsen 1994; Berggård et al. 1998; Santin and Cannas 1999), as well as in fetal tissue (current study) results from transport and deposition of hepatocyte-synthesized α1m. The transport pathways for such deposition remain to be clarified but may involve certain high molecular weight α1m complexes (notably those with IgA, albumin, and some proteinase inhibitors; Berggård et al. 1997). In this context, it is interesting to note that there is considerable variability among tissues in their relative composition of high molecular weight complexes (Berggård et al. 1998). Thus, α1m may exert different functions via different high molecular weight complexes, a form of tissue-specific functional specialization.
Related to this possibility are the immunoregulatory properties of α1m. The protein can inhibit antigen-induced lymphocyte activation, leukocyte migration and chemotaxis (Mendez et al. 1980; Lögdberg and Åkerström 1981; Åkerström and Lögdberg 1984; Lögdberg et al. 1986; Babiker-Mohamed et al. 1990a,b; Wester et al. 1998). In its dual role as plasma protein and matrix protein, α1m may provide tissues with protection from damage caused by inflammatory and/or immune reactions. Such local immunoprotection could be differentially mediated by different α1m complexes. Although overall levels of α1m in biological fluids are not markedly influenced by pathological conditions other than kidney disease (Itoh and Kawai 1990), the variability of tissue-specific α1m concentrations in health and disease remains to be elucidated.
In this context, it is intriguing to find high local concentrations of α1m in extracellular matrix at boundaries of strategic immunological importance, such as the skin (epidermodermal basement membranes) (Bouic et al. 1985), the air-tissue boundary (alveolar septa within airways) (Berggård et al. 1998), the blood–tissue boundary (the perivascular extracellular matrix) (Odum and Nielsen 1994; Berggård et al. 1998; and the present study), and the maternal blood–fetal tissue boundary (the syncytiotrophoblasts of placenta) (Berggård et al. 1999b).
Staining of “neuroendocrine” cells of the gastrointestinal tract or pancreatic islets was noted with some but not other α1m antibodies. Possible explanations include that the moiety detected could represent (a) a processed form of α1m, exhibiting only a unique epitope(-s) or (b) a different molecule crossreactive with a unique α1m epitope. The observation that anti-α1m K322 antibodies, adsorbed on immobilized urinary or recombinant α1m both lost their reactivity argues against a reactivity of the antibodies to a contaminant in the α1m preparation used for immunization because urinary and recombinant human α1m would not be expected to contain the same minor impurities. An RT-PCR study performed on adult human tissues (liver, kidney, lung, heart, spleen, muscle, thyroid, pancreas, and adrenal gland) (Berggård et al. 1998) demonstrated the presence of α1m mRNA only in the liver and, to a lesser extent, the pancreas, lending support to the first explanation. Other studies in humans (Itoh et al. 1996) and other species (Yoshida et al. 1999) also find evidence for modest levels of α1m synthesis by pancreas. These authors (Itoh et al. 1996) did not observe islet cell staining in immunohistochemical experiments, suggesting that their antiserum was one with low pancreas reactivity (see Table 2). Planned future studies include analysis of neuroendocrine staining with anti-bikunin. Because α1m and bikunin are synthesized as diproteins, the later study will hopefully clarify the synthesis/uptake issues.
In summary, we have used a reproducible immunohistochemical method for detection of α1m in archival fetal human tissue to create the first map of the distribution of α1m in developing tissues. Although its function remains to be clarified, α1m, on the basis of structural, histological, immunological, and evolutionary evidence, is likely to play a role in development, molecular transport, and immunomodulation. With respect to these and related issues, this report sets the stage for further histological studies in the functional characterization of α1m and its role in pathological processes.
Footnotes
Acknowledgements
Supported by the Department of Pathology, Albert Einstein College of Medicine, and by a grant from the Swedish Medical Research Council (project no. 7144).
The study was approved by the Institutional Review Board at the Albert Einstein College of Medicine. Drs M. Prystowsky, CM. Salafia, and S. Gupta gave critical comments to various versions of the manuscript, and Dr L. Paul–Lögdberg reviewed the language.
We thank Rachel Tucker (Immunohistochemistry), Ernestine Middleton, Dinah Carroll (both Histology), Maria Allhorn, and Ann-Sophie Johnson (both Lund) for technical support.