
Abstract
We assessed the distribution and relative staining intensity of bone morphogenetic protein (BMP)-1–7 by immunohistochemistry in tibial growth plates, epiphyses, metaphyses, and articular cartilage in one 21-week and one 22-week human fetus and in five 10-week-old Sprague–Dawley rats. In the rats, articular cartilage was also examined. BMP proteins were mostly cytoplasmic, with negligible matrix staining. Highest BMP levels were seen in (a) hypertrophic and calcifying zone chondrocytes of growth plate (BMP-1–7), (b) osteoblasts and/or osteoprogenitor fibroblasts and vascular cells of the metaphyseal cortex and medulla (BMP-1–6), (c) osteoclasts of the metaphysis and epiphysis (BMP-1,-4,-5, and −6), and (d) mid to deep zone articular chondrocytes of weanling rats (BMP-1–7). BMP staining in osteoclasts, an unexpected finding, was consistently strong with BMP-4, −5, and −6 but was variable and dependent on osteoclast location with BMP-2,-3, and −7. BMP-1–7 were moderately to intensely stained in vascular canals of human fetal epiphyseal cartilage by endothelial cells and pericytes. BMP-1,-3,-5,-6, and −7 were localized in hypertrophic chondrocytes adjacent to cartilage canals. We conclude that BMP expression is associated with maturing chondrocytes of growth plate and articular cartilage, and may play a role in chondrocyte differentiation and/or apoptosis. BMP appears to be expressed by osteoclasts and might be involved in the intercellular “cross-talk” between osteoclasts and neighboring osteoprogenitor cells at sites of bone remodeling.
Keywords
C
Chondrocyte maturation and vesiculation are likely to be controlled by external molecular cues as well as by timed expression of internal genes. Expression by growth plate chondrocytes of parathyroid hormonerelated peptide (PTHrp) (Lee et al. 1996; Kartsogiannis et al. 1997), basic fibroblast growth factor (bFGF) (Twal et al. 1994; Wezeman and Bollnow 1997), and BCL-2 (Wang et al. 1997) are believed to promote maintenance of the proliferative state. The onset of maturation is thought to be promoted by BMPs (Enomoto-Iwamoto et al. 1998; Grimsrud et al. 1998, 1999; Yazaki et al. 1998), 1,25 OH2 vitamin D3 (Boyan et al. 1989), thyroxine (Ballock and Reddi 1994; Wakita et al. 1998), retinoic acid (Iwamoto et al. 1994), c-myc (Farquharson et al. 1992; Iwamoto et al. 1993), ascorbic acid (Leboy et al. 1989), and Indian hedgehog (Vortkamp et al. 1996). Factors suggested as promoters of chondrocyte programmed cell death include BMPs (Houston et al. 1994; Zou and Niswander 1996), Bax (Wang et al. 1997), c-myc (Loveridge et al. 1993), MMP-9/gelatinase-B (Vu et al. 1998), and FAS/FAS ligand (Hashimoto et al. 1997).
In recent years a growing list of BMPs has been isolated, cloned, and sequenced. The osteoinductive BMPs, especially BMP-2–7, which were shown to induce ectopic bone formation after subcutaneous implantation in rats, are all a part of the TGF-β super-family of proteins (Wozney and Rosen 1998). BMP-7 was designated osteogenic protein-1 (OP-1) at the time of its discovery (Sampath et al. 1992) and is frequently referred to as OP-1 in the current literature. Osteogenic BMPs have been identified in bone matrix (Suzawa et al. 1999: Thomadakis et al. 1999). They signal through serine–threonine kinase receptors that reside at the surfaces of osteoprogenitor cells (Dijke et al. 1994; Liu et al. 1995). BMP-1 is not a TGF-β-related protein. Its molecular structure is related to tolloid, an astacin-like metalloproteinase that functions in the development of sea urchin, fruit fly, and frog (Schimmell et al. 1991; Maeno et al. 1993; Hwang et al. 1994). Recombinant human BMP-1 exhibits Type I pro-collagen C-peptidase activity (Kessler et al. 1996). Although a direct role for BMP-1 in skeletogenesis is unconfirmed, tolloid has recently been shown to cleave and inactivate chordin, a BMP inhibitor, thus enhancing the ventralizing effect of BMP-4 during early embryonic development in zebrafish (Blader et al 1997).
An accumulating body of evidence indicates that BMPs not only function to promote osseous differentiation but also are involved in triggering apoptosis. In addition to the evidence linking BMP-7 and the BMP-IB receptor to apoptosis of growth plate chondrocytes (Yazaki et al. 1998), expression of BMP-2 protein is associated with apoptosis of interdigital mesenchymal cells of limb bud, thus creating spaces between the fingers (Zou and Niswander 1996). Implanted BMP-2-releasing agarose beads caused apoptosis of juxtaposed Meckels’ cartilage cells during jaw development (Ekanayake and Hall 1997), and BMP-4-releasing beads implanted into Stage 20–22 chick limb buds caused massive apoptosis of precartilaginous mesenchymal cells (Buckland et al. 1998).
Because of the apparent role of BMPs in promoting both differentiation and apoptosis of chondrocytes, it is not surprising that the first reports of BMP localization in growth plate have indicated greatest expression of BMP-1–7 and BMP receptors in hypertrophic cells where both processes occur (Houston et al. 1994; Hodges et al. 1997; Vu et al. 1998; Yazaki et al. 1998; Anderson et al. 1999). The main objective of the present study was to develop a comprehensive map of relative concentration levels of BMP-1–7 proteins in cells at comparative levels of the growth plate using immunohistochemistry. During the study we also were able to evaluate BMP expression in osteoblasts and osteoclasts of the metaphysis and in chondrocytes of the superficial and deep layers of articular cartilage of growing rats. Our major findings were confirmation of greatest concentration of most BMPs in the maturing and apoptotic chondrocytes of growth plate and articular cartilage, in osteoblasts and/or osteoprogenitor stromal or vascular cells of metaphyseal cortex and medulla and, unexpectedly, in medullary and epiphyseal osteoclasts.
Materials and Methods
Tissue Preparation
Upper tibial growth plates with attached metaphyseal and epiphyseal bone and articular cartilage were obtained from five weanling Sprague–Dawley rats, sliced once longitudinally, and fixed for 24–48 hr in 4% paraformaldehyde. The same region of upper tibia from two 21-week gestation human fetuses with normal limb development was removed and fixed in 4% paraformaldehyde. In both human cases, death occurred spontaneously in utero at approximately 12 hr before fixation. After fixation, the tissues were decalcified in 10% EDTA in distilled water for approximately 1 week at room temperature with daily agitation. The decalcified samples were dehydrated in changes of ethanol at increasing concentration, embedded in low melting temperature paraffin, and sectioned at 5-μm thickness.
Immunoperoxidase Staining
Embedding medium was removed from the sections by exposure to xylene, followed by incubation in ethanol solutions (100% ethanol followed by 95%) to begin rehydration. Deparaffinized, rehydrated sections were incubated in 3% Triton X-100 in PBS to permeabilize and were blocked with DAKO (Carpinteria, CA) blocking solution before incubation overnight in primary antibody solution. Primary antibodies were diluted in PBS as follows: anti-BMP-1, 1:2000; anti-BMP-2, −3, −5, −6, and −7, 1:100; anti-BMP-4, 1:5. After incubation in primary antibody, sections were briefly incubated in the appropriately diluted secondary antibody (biotinylated anti-rabbit, anti-mouse, or anti-goat immunoglobulins), followed by incubation in streptavidin–peroxidase and buffered substrate solution (pH 7.5) containing hydrogen peroxide and 3,3-diaminobenzidine chromogen solution. (All of the above reagents were purchased from DAKO). The sections were counterstained in 0.5% Mayers’ hematoxylin (DAKO) or, in some instances, in 5% toluidine blue in 100% ethanol.
Expression of BMP-1–7 proteins in weanling rat growth plate and metaphysis a
a + + +, maximal; ++, moderate; +, minimal.
bStaining quite variable.
Primary Antibodies
Anti-BMP-1/tolloid rabbit antibody was a polyclonal raised against recombinant human BMP and crossreacts with BMP-1 and tolloid but not with other BMPs or TGF-β (kindly provided by Dr. Paul Reynolds, University of Rochester). Rabbit anti-BMP-2 and −3 were polyclonal antibodies raised against recombinant human BMP-2 (serum ID W12) and −3 (serum ID W22) (kindly provided by Dr. I.K. Moutsatsos, Genetics Institute, Inc.). Anti-BMP-2 is specific for BMP-2 and anti-BMP-3 crossreacts with BMP-2 and −5. Anti-BMP-4 was a monoclonal raised against recombinant human BMP-4 and is monospecific for BMP-4 (kindly provided by Dr. K. Masuhara, Osaka University). Rabbit anti-BMP-6 polyclonal antibodies, raised against the precursor and mature forms of recombinant human BMP-6, are specific for these forms of BMP-6 and do not react with other BMPs (kindly provided by Dr. S. Gitelman, University of California, San Francisco). Anti-BMP-5 and −7 were goat polyclonal antibodies obtained from Santa Cruz Biotechnology (Santa Cruz, CA). Both anti-BMP-5 and −7 were raised against C-terminal peptides of the mature regions of BMP-5 and −7 and do not crossreact with other BMPs.
Human and Animal Subjects
Human embryonic tissues were obtained at autopsy under authorization by the University of Kansas Medical Center Human Subjects Review Board. Rat tissues were obtained after humane sacrifice using procedures authorized by the University of Kansas Medical Center Institutional Animal Care and Use Committee, ACUP No. 86-02-03-01.
Results
Growth Plate
There was a similar relative distribution of BMP-1–7 in rat and human growth plates (Tables 1 and 2). BMP staining was highest in the cytoplasm of maturing and late hypertrophic chondrocytes, with perhaps a slight predominance during maturation and early hypertrophy (Figures 1 and 2). Staining of all BMPs was light and variable in reserve and proliferative zone chondrocytes. The BMPs were not detected to any significant degree in cartilage or bone matrix.
Metaphysis
Osteoblasts showed moderate to intense staining for all the BMPs (Tables 1 and 2). However, with some BMPs (especially BMP-2, −3, −5, −6, and −7), endosteal osteoblast staining varied from intense to undetectable depending on their location (Figure 3). Osteoblasts of the metaphyseal cortex were more uniformly positive. With BMP-2, −3, −5, −6, and −7 in rat and BMP-3 and 7 in human, marrow stromal cells and/or vascular cells adjacent to endosteal osteoblasts were often positively stained (Figure 3B).
Osteoclasts
These resorptive cells were identified by their multinuclearity and their location close to trabecular and sub-periosteal bone surfaces of cortex, metaphysis, and epiphyseal secondary ossification centers. BMP-1–7 were stained in the cytoplasm of rat and human osteoclasts (Figure 4). Staining for BMP-1, −4, −5, and −6 was moderate to intense in virtually all osteoclasts (Tables 1 and 2; Figure 4). Immunostaining for BMP-2 and 3 was more variable, with some negatively-reacting osteoclasts. BMP-7 staining was minimal in rat osteoclasts (Table 1).
Expression of BMP-1–7 proteins in human fetal growth plate and metaphysis a
a+ + +, maximal; ++, moderate; +, minimal.
bStaining quite variable.
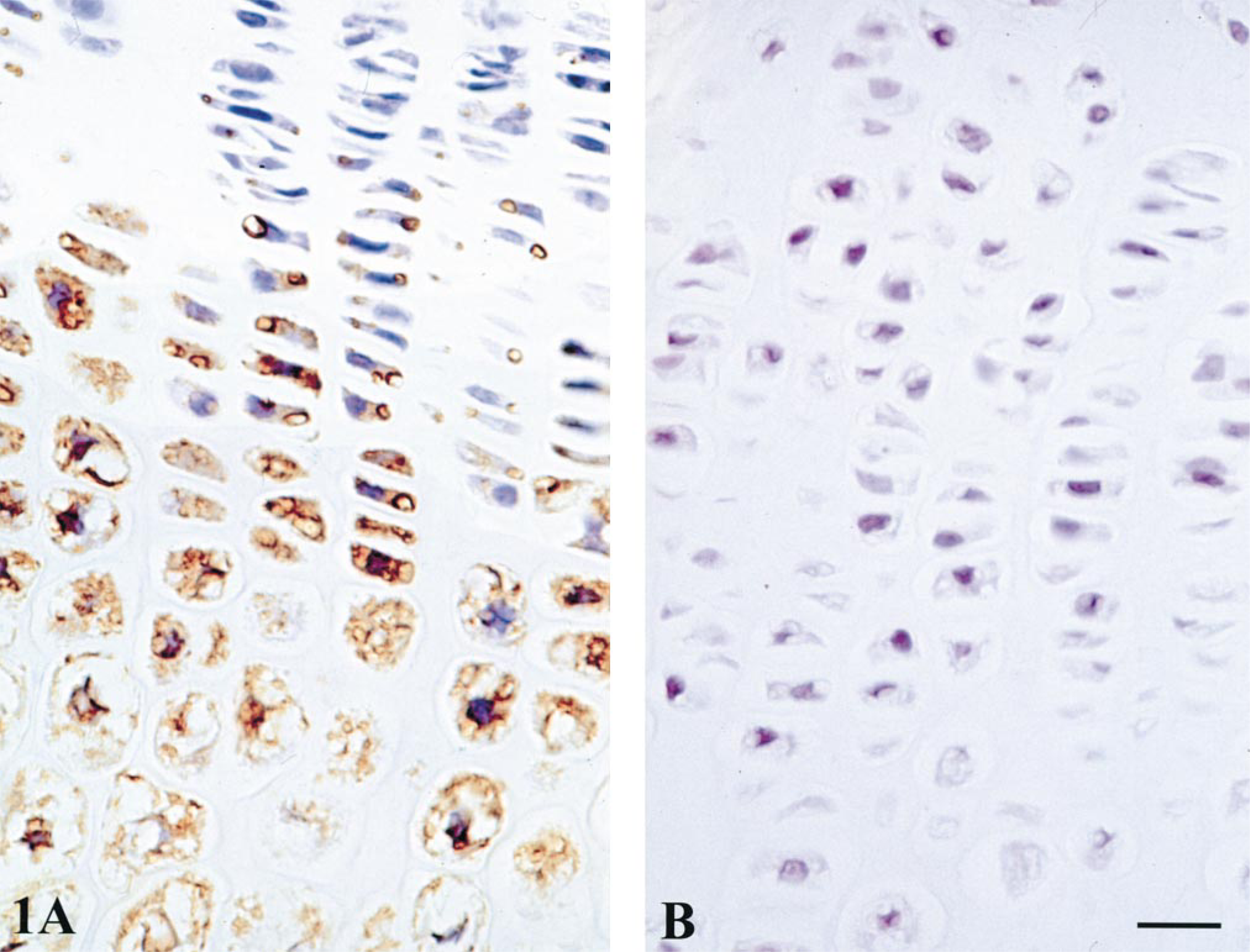
Rat growth plate immunostained for BMP-2. (
Articular Cartilage and Vascular Canals
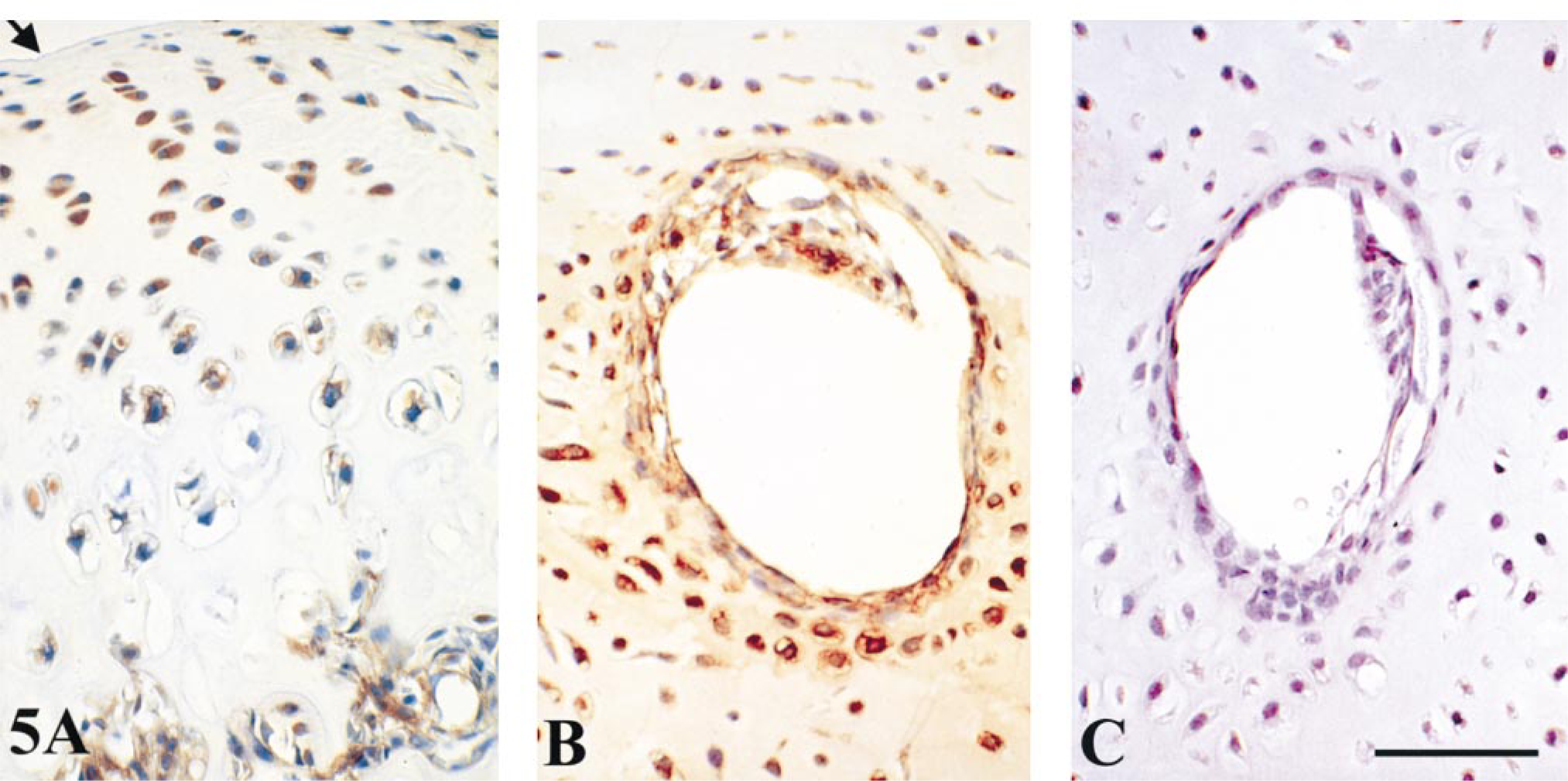
In weanling rat, BMP-1–7 immunostaining was most intense in the cytoplasm of mid-zonal (maturing) and mid- to deep-zonal (hypertrophic) chondrocytes of articular cartilage (Table 3; Figure 5A). BMP-1 staining was less intense and more variable in surface articular chondrocytes (Table 3).
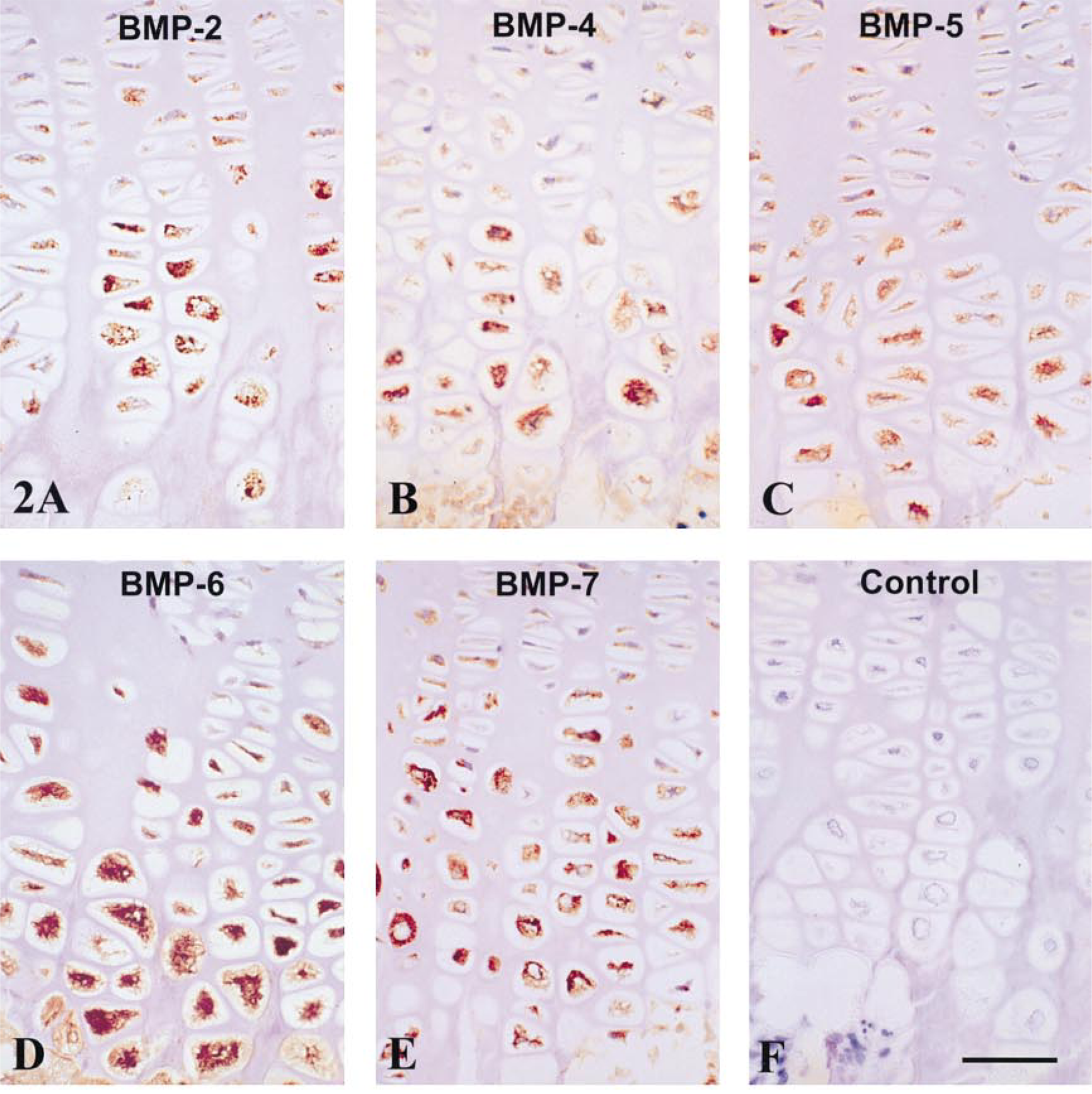
Human growth plate immunostained for BMP-2, −4, −5, −6, and −7. Staining for all BMPs was intense in the cytoplasm of maturing and hypertrophic chondrocytes but weak or absent in the flattened chondrocytes of the proliferative zone. Matrix staining for BMPs was not observed. (
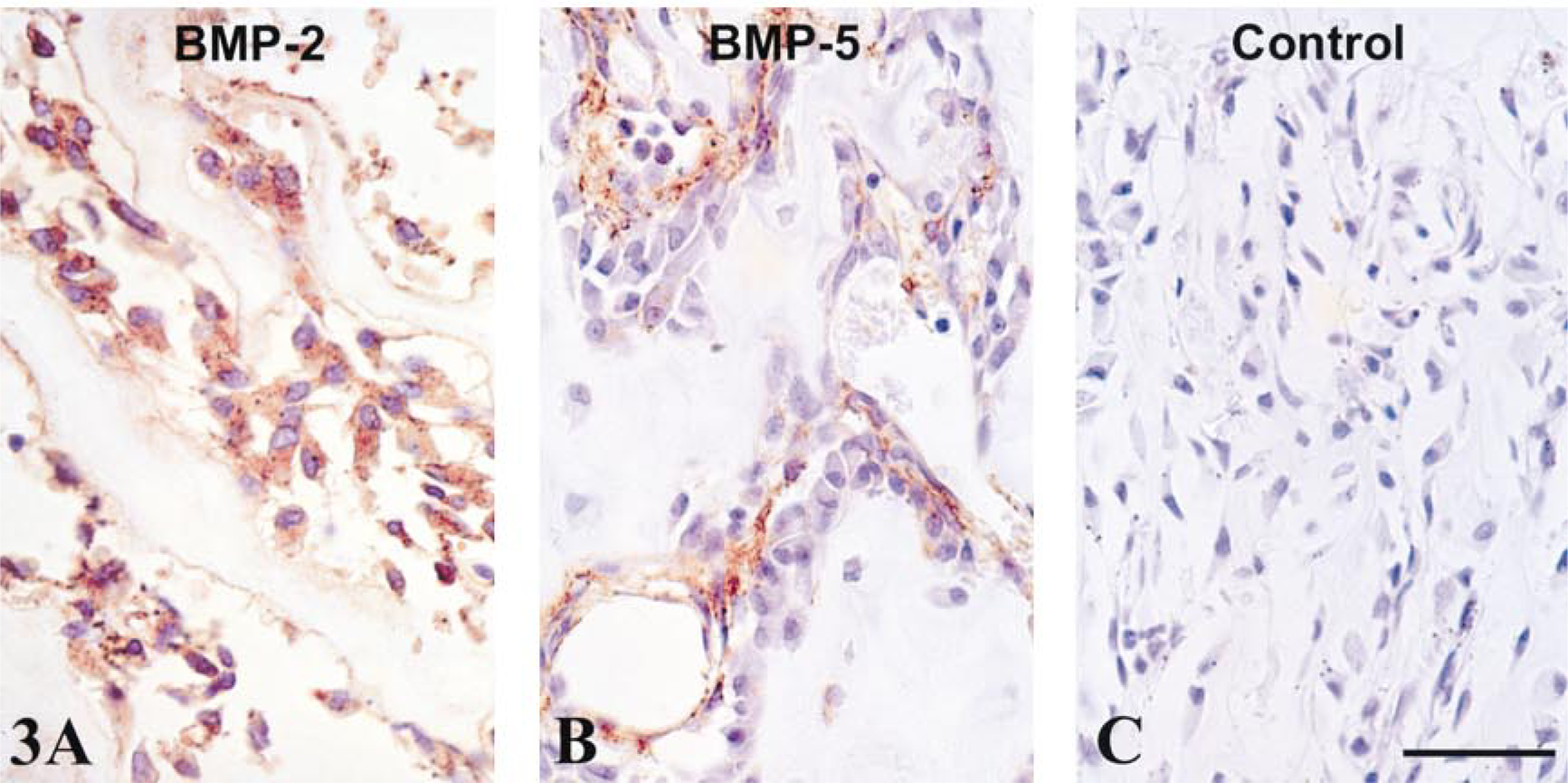
Most BMPs were localized in the cytoplasm of metaphyseal osteoblasts and/or perivascular marrow stromal cells. (
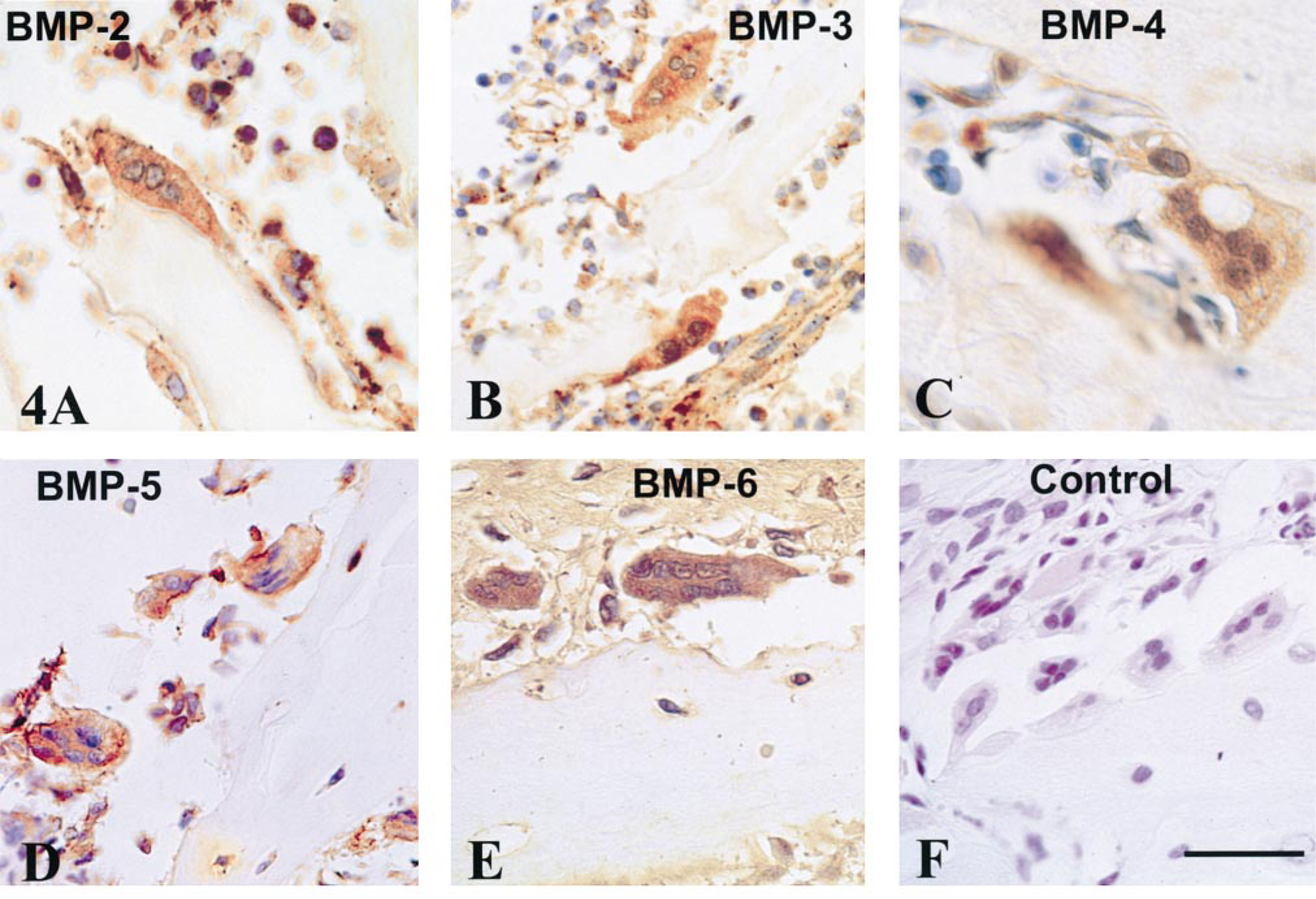
Osteoclasts of the metaphysis stained moderately to intensely for most BMPs. (
BMP-4 staining in rat articular cartilage (
Expression of BMP-1–7 proteins in rat articular cartilage and human epiphyseal vascular canals a
a + + +, maximal; ++, moderate; +, minimal.
bStaining quite variable.
In human fetal epiphyses, the secondary ossification centers had not yet formed and vascular canals penetrated these regions, which were composed primarily of relatively undifferentiated cartilage. Chondrocytes located 5–10 cell layers deep to the cartilage canals were larger in diameter, suggesting the onset of hypertrophy, and they stained moderately to intensely for BMP-1–7 (Table 3; Figure 5B). An unexpected finding was the presence of significant cytoplasmic immunostaining for BMP-1–7 in endothelial cells and/or pericytes of the vascular canals (Figure 5B).
Discussion
This study has confirmed earlier reports of localization of BMP-1, −4, −6, and −7 in maturing hypertrophic and apoptotic chondrocytes of growth plate (Hodges et al. 1997; Vu et al. 1998; Yazaki et al. 1998; Anderson et al. 1999), and demonstrated a similar staining pattern for BMP-2, −3, and −5. Thus, a comprehensive picture emerges indicating high expression levels of most BMPs by chondrocytes undergoing maturation and programmed cell death.
Much experimental evidence already reported indicates a role for BMPs in the promotion of skeletal cell differentiation. BMP-2, −4, −6, and −7 can induce heterotopic cartilage and bone formation when implanted subcutaneously into rats and mice (Wang et al. 1990; Sampath et al. 1992; Takaoka et al. 1993; Gitelman et al. 1994). During limb bud development, BMP-2, −4, and −7 are expressed in the apical epidermal ridge, an organizer tissue for the limb bud (Hogan 1996). BMPs are also expressed during tooth development (Heikinheimo 1994; Thesleff et al. 1996; Aberg et al. 1997) and in regenerating cartilage and bone cells during bone repair (Nakase et al. 1994; Bostrom et al. 1995; Urist et al. 1997; Kitazawa et al. 1998; Onishi et al. 1998). Our demonstration above of high levels of most BMPs during early maturation of growth plate chondrocytes and in differentiating osteoblasts of the metaphysis adds support to the hypothesis that BMPs are promoters of osseous differentiation during pre- and postnatal bone growth.
In growth plate, however, chondrocyte differentiation and maturation overlap and blend into the subsequent phase of programmed cell death, and both phases are associated with high intracellular levels of BMPs. Given the continuum between differentiation and programmed cell death in growth plate chondrocytes, and the overlap of high BMP expression levels during both phases, it may prove difficult to define and experimentally separate the differentiation-promoting from the cell death-promoting activities of BMPs in these cells. Certainly, there is evidence for a role of BMPs in promoting apoptosis during skeletal development (Hogan 1996; Zou and Niswander 1996; Ekanayake and Hall 1997; Buckland et al. 1998). In addition, the staining pattern of BMPs we observed in articular chondrocytes, with highest concentration in mid- and deep-zone cells, is consistent with the demonstration that most apoptosis occurs in these zones (Hashimoto et al. 1997, 1998; Kouri et al. 1997; Adams and Horton 1998).
To our knowledge, this study presents the first demonstration of BMPs in chondrocytes associated with epiphyseal cartilage vascular canals. Moderate to intense staining of BMP-1–7 was seen in perivascular hypertrophic chondrocytes and of BMP-1–7 in endothelium and pericytes of cartilage vessels (Table 3; Figure 5). Cartilage canals are believed to participate in the development of secondary ossification centers of the epiphysis. However, the interaction between these penetrations and the coincident hypertrophy of chondrocytes and the mineralization of juxtaposed cartilage matrix is poorly understood. As recently demonstrated by Roach et al. (1998), using an interesting in vitro experimental model, vascular canals multiply and converge at the site of formation of future secondary ossification centers. Perivascular chondrocytes adjacent to the convergent vascular canals became hypertrophic and were associated with matrix calcification. This developmental pattern suggested to Roach et al. (1998) that a release of diffusible factors from penetrating vessels may be the trigger for chondrocyte maturation and hypertrophy. Such a suggestion is supported by our demonstration that endothelium and perivascular cells of cartilage canals produce a range of BMPs that could trigger differentiation, matrix vesicle biogenesis and, ultimately, programmed cell death by neighboring chondrocytes. A similar phenomenon may be involved in the pathogenesis of calcific atherosclerosis. During this process, expression of BMP-2 by vascular cells results in bone-like metaplasia and calcification in atherosclerotic lesions (Bostrom et al. 1993).
It is not widely recognized that osteoclasts (as well as osteoblasts) contain significant amounts of BMPs. In this study we have demonstrated staining of BMP-1–7 by osteoclasts in human fetal and weanling rat metaphysis, a site at which active osteoclast-mediated bone remodeling occurs. Our finding of BMPs in osteoclasts could possibly be due to an artifact related to immunostaining procedures and does not prove that the presence of BMP proteins in osteoclasts is necessarily due to local BMP expression. It could, for example, be due to phagocytosis of extracellular BMPs by osteoclasts. Nevertheless, this supports our preliminary reports of BMP-1, −4, and −6 in growth plate osteoclasts (Hodges et al. 1997; Anderson et al. 1999) and confirms earlier observations by Nishimura et al. (1997), who immunohistochemically demonstrated BMP-2 in osteoclasts in ectopic bone induced by periosteum, by Onishi et al. (1998), who demonstrated expression of BMP-2/4 and −7 by osteoclasts in regenerating bone during fracture repair, and by Helder et al. (1998), who observed BMP-7 in osteoclasts of developing alveolar bone. (The latter two studies employed in situ hybridization.) BMP-4 also has been observed by in situ hybridization in osteoclasts of repair bone by Nakase (personal communication, 1998). Therefore, there is gathering evidence that active osteoclasts, during bone growth, repair, and remodeling, do express BMPs. BMP expression by osteoclasts may constitute an important component of the intercellular “cross-talk” that leads to osteoblast recruitment and differentiation at a local bone remodeling site. Aubin (1999) has reported that marrow monocytes, which are the precursor cells of osteoclasts (Suda et al. 1995), are capable of stimulating osteoblastic differentiation by marrow stromal osteoprogenitor cells in co-culture. In this model, BMP expression and release by osteoclast precursor monocytes could provide an important component of the intercellular communication that leads to osteoblastic differentiation.
It also has been suggested that BMPs, deposited in bone matrix by osteoblasts, might be released for paracrine osteogenic activity from osteoclastic resorption sites of bone remodeling (Manolagas and Jilka 1995). However, we found little immunohistochemical evidence of BMP entrapment in bone or cartilage matrix. The failure to detect stainable BMPs in bone and cartilage matrix could be due to technical artifact, e.g., a release of BMPs from matrix during decalcification. Nevertheless, assuming that osteoclasts themselves contain and may release BMPs, it would not be necessary to postulate BMP release from bone matrix to explain recruitment and promotion of differentiation by preosteoblastic marrow stromal cells. Further studies will be required to determine whether BMPs generated by osteoclasts are functioning in an autocrine manner, promoting osteoclast differentiation and survival, and/or acting as a paracrine factor in the recruitment and differentiation of osteoblast precursor cells at sites of bone remodeling.