
Abstract
Melanocytes are cells of neural crest origin. In the human epidermis, they form a close association with keratinocytes via their dendrites. Melanocytes are well known for their role in skin pigmentation, and their ability to produce and distribute melanin has been studied extensively. One of the factors that regulates melanocytes and skin pigmentation is the locally produced melanocortin peptide α-MSH. The effects of α-MSH on melanogenesis are mediated via the MC-1R and tyrosinase, the rate-limiting enzyme in the melanogenesis pathway. Binding of α-MSH to its receptor increases tyrosinase activity and eumelanin production, which accounts for the skin-darkening effect of α-MSH. Other α-MSH-related melanocortin peptides, such as ACTH1–17 and desacetylated α-MSH, are also agonists at the MC-1R and could regulate melanocyte function. Recent evidence shows that melanocytes have other functions in the skin in addition to their ability to produce melanin. They are able to secrete a wide range of signal molecules, including cytokines, POMC peptides, catecholamines, and NO in response to UV irradiation and other stimuli. Potential targets of these secretory products are keratinocytes, lymphocytes, fibroblasts, mast cells, and endothelial cells, all of which express receptors for these signal molecules. Melanocytes may therefore act as important local regulators of a range of skin cells. It has been shown that α-MSH regulates NO production from melanocytes, and it is possible that the melanocortins regulate the release of other signalling molecules from melanocytes. Therefore, the melanocortin signaling system is one of the important regulators of skin homeostasis.
M
There is increasing evidence that melanocytes are not simply melanin-producing cells but may have a number of functions. Melanocytes are capable of secreting a wide range of signaling molecules and it has been suggested that they could function as regulatory cells in maintaining epidermal homeostasis (Slominski et al. 1993a). Their ability to respond to regulatory peptides such as α-MSH might be an integral part of such a function. This article reviews the role of the epidermal melanocyte in human skin pigmentation, its control by the melanocortin signaling system, and explains how, by studying this control system, we are beginning to appreciate more about the functions of this particular cell.
Melanocytes and Skin Pigmentation
Melanocytes are derived from the neural crest. During development, presumptive melanocytes (melanoblasts) migrate to various sites including the skin, where they proliferate and then differentiate into melanin-producing cells.
Under normal conditions it is not the numbers of melanocytes in the skin that determine the degree of pigmentation but their levels of activity. Although there are regional variations in the density of epidermal melanocytes, their numbers are consistent even in different skin types and ethnic groups. Therefore, constitutive or basal skin pigmentation is considered to depend on the level of melanogenic activity and the transfer of melanin into the neighboring keratinocytes. The type of melanin is also likely to be important, and it is recognized that human melanocytes produce both the brown-black eumelanin and reddish-yellow phaeomelanin (Hunt et al. 1995). As might be expected, phaeomelanin is the major type in red hair and also predominates in the epidermis of skin types I and II (Thody et al. 1991). Eumelanin, on the other hand, is present in large amounts in individuals with dark skin and hair. It is generally accepted that eumelanin is the more photoprotective of the two melanins.
The synthesis of melanin takes place in the melanosome. This is a specialized intracellular membrane-coated organelle that originates from the endoplasmic reticulum. During its development the melanosome acquires tyrosinase and the tyrosinase-related proteins 1 and 2 (TRP1, TRP2). Tyrosinase is the rate-limiting enzyme for melanogenesis and catalyzes the conversion of L-tyrosine to dopaquinone, which is required for the synthesis of both eumelanin and phaeomelanin. It may also catalyze later steps specific to the eumelanin pathway and this could explain why eumelanogenesis is especially dependent on tyrosinase. Less is known about the control of phaeomelanin synthesis, although it appears to be less dependent on tyrosinase and can proceed even when the levels of tyrosinase activity are virtually undetectable (Burchill et al. 1986). The functions of TRP-1 are not clear, and although there are reports that it has 5,6 dihydroxyindole carboxylic acid (DHICA) oxidase activity, this has yet to be confirmed. It has been suggested that TRP-1 functions to stabilize tyrosinase (Kobayashi et al. 1998; Manga et al. 2000).
Once melanin is produced the melanosomes are transferred into the neighboring keratinocytes. The size of these organelles and their numbers are important in determining pigmentation. The melanosomes in black skin are larger than their counterparts in white skin and are packaged as single units rather than in groups. This has the effect of retarding their degradation in the keratinocytes and contributes to a higher level of skin pigmentation. At present, little is known about the mechanisms and regulation of melanosome transfer. It appears that association of melanosomes with microtubules and actin filaments via motor proteins, such as kinesin, dynein, and myosin V, is important for melanosome movement along the dendrites and for subsequent transfer to keratinocytes (Provance et al. 1996; Wu et al. 1997,1998; Lambert et al. 1998; Hara et al. 2000; Vancoillie et al. 2000a,b). Melanocyte dendricity and contact with keratinocytes is likely to be essential for the transfer of melanin-containing melanosomes. A recent study showed that activation of the protease-activated receptor 2 (PAR-2), which is expressed only on keratinocytes, increases melanin transfer to keratinocytes (Seiberg et al. 2000).
An increase in skin pigmentation over the basal constitutive level is referred to as facultative pigmentation. A major stimulus of facultative pigmentation in humans is UVR. UVR-induced skin pigmentation or tanning, as it is commonly known, involves several processes. There is an increase in the numbers of active epidermal melanocytes, but whether this is a result of increased proliferation, enhanced recruitment, or both events is not entirely clear. The expression of tyrosinase and other related melanosomal proteins are also increased in response to UVR and, as a result, melanogenesis is stimulated. Although phaeomelanogenesis and eumelanogenesis are increased in response to UVR, it is the concentrations of eumelanin that correlate better with the degree of tan. Therefore, eumelanin is believed to make the greater contribution in the tanning response (Thody et al. 1991). The melanocytes also become more dendritic in response to UVR, leading to greater interaction with keratinocytes and an extension of the epidermal-melanin network. It is also likely that there is increased transfer of melanosomes into the keratinocytes after UVR.
Melanocortins Function as Mediators in the Pigmentary Response
α-MSH is produced, together with several other peptides, by the proteolytic cleavage of the large precursor protein pro-opiomelanocortin (POMC). The main site of α-MSH production is the pars intermedia of the pituitary gland. However, because of its poorly developed pars intermedia, the human pituitary secretes only small amounts of α-MSH except under pathological conditions (Thody et al. 1985; Eberle 1988; Pears et al. 1992). Nevertheless, α-MSH and other melanocortin peptides are produced at extrapituitary sites, including the skin (Thody et al. 1983; Wakamatsu et al. 1997; Slominski et al. 2000). Epidermal keratinocytes are a major source of these peptides (Slominski et al. 1992,1993a,b,1996; Bhardwaj and Luger 1994), although they are produced in other cell types in the skin, including melanocytes (Lunec et al. 1990; Wakamatsu et al. 1997) and Langerhans cells (Morhenn 1991). It has been suggested that the pigmentary effects of UVR are mediated via melanocortin peptides such as α-MSH (Bolognia et al. 1989; Chakraborty et al. 1995). Keratinocytes and melanocytes secrete melanocortin peptides in response to UVR (Schauer et al. 1994; Chakraborty et al. 1996). Therefore, α-MSH could act as a paracrine and an autocrine factor in the regulation of melanocytes and skin pigmentation. In addition to its effects on melanocortin peptide secretion, UVR upregulates the expression of the α-MSH receptor on mouse melanoma cells (Chakraborty et al. 1995) and increases the binding of α-MSH to human melanocytes (Thody et al. 1993). Therefore, UVR elevates not only the levels of α-MSH in the skin but also the responsiveness of the melanocytes to this peptide. It therefore appears likely that α-MSH and possibly other melanocortins function as mediators of the pigmentary response in the skin.
How Does α-MSH Act to Regulate Skin Pigmentation?
Melanocortin peptides exert their effects through melanocortin receptors (MC-Rs). These are seven transmembrane domain G-protein-coupled receptors, and to date five subtypes have been cloned. The MC-1R, which was the first subtype to be sequenced and characterized (Mountjoy et al. 1992), is encoded by a gene that is localized on the 16q24.3 chromosome (Gantz et al. 1994). Although expressed by melanocytes and melanoma cells and considered to be a control point for pigmentation (Siegrist et al. 1989; Donatien et al. 1992; Lunec et al. 1992; Eberle et al. 1993; Thody et al. 1993), the MC1-R is also present on other cell types, such as human monocytes (Bhardwaj et al. 1997), endothelial cells (Hartmeyer et al. 1997; Scholzen et al. 1999), and keratinocytes (Chakraborty and Pawelek 1993; Jiang et al. 1996; Chakraborty et al. 1999). On binding to the MC1-R, α-MSH activates adenylate cyclase which, in turn, causes an increase in intracellular cAMP. This is the classical pathway by which α-MSH is believed to mediate its melanogenic effects on melanocytes. Increases in cAMP result, via protein kinase A (PKA), in the activation of tyrosinase, the rate-limiting enzyme in the melanin pathway. Evidence suggests that α-MSH increases the expression, de novo synthesis, and activation of tyrosinase (Burchill et al. 1989; Hunt et al. 1994a; Suzuki et al. 1996). It has been shown that in B16 melanoma cells specific inhibition of phosphatidylinositol-3-kinase (PI3-kinase) and its target, the serine/threonine kinase p70S6-kinase, by the cAMP-elevating agent forskolin markedly increases the melanin content of these cells (Buscá et al. 1996). Therefore, the PI3-kinase/p70S6-kinase pathway may be involved in regulating melanogenesis. On the other hand, activation of the MAP-kinase pathway by cAMP may in some cases downregulate melanogenesis, because it causes phosphorylation and degradation of the microphthalmia-associated transcription factor (MITF), a factor required for transcription of the melanogenic enzymes TRP-1 and TRP-2 (En-glaro et al. 1998; Buscá and Ballotti 2000). The possibility that other intracellular pathways are also activated on binding of α-MSH to its receptor must not be excluded. For example, there is evidence that protein kinase C is involved in mediating the melanogenic actions of α-MSH (Buffey et al. 1992; Park et al. 1996). In addition to its receptor-mediated effects, there is evidence to suggest that α-MSH is able to regulate tyrosinase activity independently of the MC1-R. α-MSH binds (6R)-L-erythro 5,6,7,8 tetrahydrobiopterin (6-BH4) and the latter has been shown to regulate the availability of <sc<L-tyrosine and the activity of tyrosinase in melanocytes (Schallreuter et al. 1994,1997,1999; Wood et al. 1995). Because α-MSH is particularly abundant in human melanocytes (Wakamatsu et al. 1997) and serves as a chaperone for 6-BH4, it could, through this interaction, modulate the synthesis of melanin (Moore et al. 1999). It has further been suggested that if the concentrations of α-MSH exceed those of 6-BH4, then the peptide itself could function as a substrate for tyrosinase by virtue of its tyrosine residue in position 2. The availability of 6-BH4 in the melanocyte may therefore be important for melanogenesis and in determining responsiveness to α-MSH.
Rather than producing large increases in melanin, α-MSH regulates the pattern of melanogenesis by preferentially stimulating the synthesis of eumelanin at the expense of phaeomelanin (Hunt et al. 1995). This would account for its skin-darkening effects because eumelanin is the most important of the two melanins for skin pigmentation.
Stimulation of melanogenesis is not the only effect of α-MSH on human melanocytes. For example, there is evidence that α-MSH stimulates melanocyte dendricity (Hunt et al. 1994a, c; Wakamatsu et al. 1997). Because dendrites are important for the formation of the epidermal-melanin unit and the transfer of melanin, the regulation of dendricity by α-MSH may be vital for the pigmentary response. It is not clear how α-MSH acts to stimulate melanocyte dendricity, but this process may be dependent on activation of several intracellular signaling pathways. It appears likely that cAMP is important, and there is evidence that, by acting on the GTP-binding proteins Rac and Rho, cAMP increases actin disorganization and promotes melanocyte dendricity (Buscá et al. 1998; Scott and Cassidy 1998; Buscá and Ballotti 2000). This action of cAMP, like that on melanogenesis, could involve an inhibition of PI3 kinase (Buscá and Ballotti 2000). Another way in which α-MSH affects melanocytes, and possibly pigmentation, is by protecting these cells from the damaging effects of oxygen radicals, such as the superoxide anion (Valverde et al. 1996a,b). This could depend on the activation of tyrosinase because this enzyme is able to utilize superoxide anion as a substrate for melanogenesis (Wood and Schallreuter 1991; Tobin and Thody 1994). α-MSH could also protect melanocytes through its ability to complex with 6-BH4. As discussed above, the latter controls melanogenesis but its oxidized product, 6-biopterin, is cytotoxic to melanocytes (Schallreuter et al. 1994). By complexing with 6-BH4, α-MSH could control the redox status of the pterin and in this way affect both melanogenesis and melanocyte survival.
The acetylated form of α-MSH has been the most well studied of the pigmentary melanocortins and is considered to be an important regulator of the pigmentary response in lower vertebrates and many mammals. However, the predominant form of α-MSH in human skin, as in the hypothalamus, is desacetyl α-MSH (Thody et al. 1983; Wakamatsu et al. 1997). This peptide binds to the human MC1-R with an affinity lower than that of acetylated α-MSH (Tsatmali et al. 1999a) and for this reason is often dismissed as having any pigmentary significance. Recent evidence suggests that desacetyl α-MSH acts as a partial agonist at the human MC1-R and thus, by opposing the actions of acetylated α-MSH, could influence melanogenic responses (Yukitake et al. 1999). It has been shown that desacetyl α-MSH acts in a similar way to oppose the actions of α-MSH on the melanophores of the Anolis lizard (McCormack et al. 1982).
ACTH peptides, e.g., ACTH1–39 and ACTH1–17, are also found in human skin and in greater abundance than α-MSH (Wakamatsu et al. 1997). ACTH1-39 has melanogenic activity (Hunt et al. 1994b,c; Abdel-Malek et al. 1995) and ACTH1–17, which binds to the human MC-1R with an affinity comparable to that of acetylated α-MSH, is particularly active in this respect (Wakamatsu et al. 1997; Tsatmali et al. 1999a,b). However, unlike α-MSH, which produces a typical sigmoidal dose-response curve, ACTH peptides produce a biphasic dose-response melanogenic curve in human melanocytes (Hunt et al. 1994b; Tsatmali et al. 1999a). This could be explained by their ability to activate other signaling systems that are not activated by α-MSH peptides. Therefore, whereas α-MSH activates the cAMP pathway, ACTH1–17 may stimulate MC-1R coupling to both the cAMP-and IP3-dependent pathways (Tsatmali et al. 1999b).
Is the MC-1R a Control Point in the Regulation of Pigmentation?
As mentioned above, activation of the MC-1R could be pivotal for UVR-induced melanogenesis. However, at present little is known about the transcriptional regulation of MC-1R expression beyond the effects of UVB and peptides, such as endothelin-1, that are released in the skin in response to UVR (see review by Abdel-Malek et al. 1999). However, there appears to be little doubt that the melanocortins are of significance as pigmentary peptides. It follows that POMC expression and its processing could represent important control points for skin pigmentation. Support for this was provided by the recent report of red hair and light skin in a patient with a nonsense mutation in the POMC gene that resulted in a complete absence of α-MSH and ACTH (Krude et al. 1998). Earlier observations had suggested that the pigmentation phenotype is associated with melanocyte responsiveness to α-MSH. Melanocytes from red-haired persons were totally unresponsive, failing to show melanogenic and dendritic responses to α-MSH and/or ACTH when in culture (Hunt et al. 1996). These findings implicated the MC-1R as a control point for pigmentation phenotype (Hunt et al. 1996) and were consistent with the finding of several MC-1R variants in red-haired individuals (Valverde et al. 1995).
Many MC-1R mutants have been identified in humans and mammals and were shown to be associated with pigmentation phenotypes (Valverde et al. 1995; Koppula et al. 1997; Smith et al. 1998; Flanagan et al. 2000). More than 30 human MC-1R variants have been reported to date and there is evidence for selection in several populations (reviewed by Rees 2000). In Europeans there is a high level of diversity, and many variants affect MC-1R function. Certain of these mutations (Arg142His, Arg151Cys, Arg160Trp, and Asp294His) are common among individuals with red hair and poor tanning ability. It was subsequently shown in mutagenesis studies that these mutations result in reductions of intracellular cAMP levels in response to α-MSH (Frändberg et al. 1998; Schiöth et al. 1999) which, in turn, would predictably lead to reduced eumelanogenesis. As a result, phaeomelanin production prevails, and this would explain not only the red hair phenotype but also the poor tanning ability in people carrying these mutations.
There are some doubts as to whether the MC-1R is the only control point for the red-hair phenotype. There have been reports of homozygous and compound heterozygous mutations of the MC-1R in individuals that do not have red hair (Smith et al. 1998; Rees et al. 1999; Rees 2000). Furthermore, melanocyte cultures from red-heads that were unresponsive to α-MSH also failed to respond to cAMP with increases in melanin content, suggesting that the unresponsiveness was due to factors downstream of the MC-1R (Hunt et al. 1996).
MC-1R alleles, including the Val60Leu variant, have also been identified in an Asian group (Harding et al. 2000). In an earlier study by Hunt et al. (1996), it was observed that one of eight melanocyte cultures established from Asian donors failed to show a melanogenic response to α-MSH. This is consistent with the view that Asians may express non-functional MC-1R alleles (Harding et al. 2000). The situation with black Africans is less clear. In the study by Harding et al. (2000), no loss of function MC-1R mutations were found in a group of 106 Africans. However, the Val60Leu and Arg151Cys variants were found as alleles of the MC-1R in an African donor (Dr. N. Smit personal communication). Such mutations might be expected to affect melanogenesis but, surprisingly, this was not the case. Furthermore, the presence of these alleles contradicts the view expressed above that MC-1R mutations are absent in black African populations.
The above observations on Asian and African populations suggest that, in dark skin types, the inherited high level of melanogenesis is not necessarily dependent on the functionality of the MC-1R receptor. Recent evidence suggests that there are control points downstream of the melanocortin signaling system that could be important in the regulation of tyrosinase and hence melanogenesis. One such control point is the active transport of <sc<L-phenylalanine into melanocytes and its conversion to <sc<L-tyrosine by phenylalanine hydroxylase (Schallreuter and Wood 1999). It is interesting that, as well as being a substrate in the melanin pathway, <sc<L-tyrosine (and <sc<L-dopa) have been shown to act as positive regulators of melanogenesis (Slominski et al. 1998; Slominski and Paus 1994). Melanosomal pH is also another downstream control point. It has been shown that near-neutral melanosomal pH facilitates melanogenesis independently of transcriptional control (Ancans and Thody 2000; Ancans et al. 2001). These observations have been confirmed in a recent study by Fuller et al. (2001). Therefore, although signaling MC-1R is relevant for pigmentary processes in the majority of light skin types, the debate as to the importance of the MC-1R signaling system in dark skin types is still open and needs further investigation.
α-MSH Modulates NO Production by Melanocytes
As well as producing melanin, melanocytes are able to secrete a wide range of signal molecules in response to UVR and other stimuli. One such substance is nitric oxide (NO) (Graham et al. 1997; Tsatmali et al. 2000), a potent signaling molecule known to be involved in the regulation of a wide variety of cellular processes (for reviews see Stuehr and Nathan 1989; Liew and Cox 1991; Lowenstein and Snyder 1992).
Human melanocytes produce NO in response to UVR and bacterial lipopolysaccharide (LPS) (Tsatmali et al. 2000). In the same study it was shown that α-MSH could modulate these effects of UVR and LPS. It was found that α-MSH alone could increase NO production but, when present together with LPS, it inhibited the stimulatory effect of the latter. How α-MSH acts to affect the production of NO in melanocytes is not clear. NO is produced via the enzymatic action of nitric oxide synthase (NOS), of which both constitutive (cNOS) and inducible (iNOS) isoforms exist. In melanocytes iNOS expression was seen in control unstimulated cells, and this suggests that, contrary to the situation in most cell types, low levels of iNOS are constitutively expressed in melanocytes (Tsatmali et al. 2000). It is possible that α-MSH modulates the activity of iNOS and hence NO production from melanocytes. It has been recently shown that α-MSH is indeed able to inhibit NFκB in melanocytes, a transcription factor directly involved in the activation of the iNOS gene transcription (Haycock et al. 1999). Whether this action is mediated via the MC-1R/cAMP signaling pathway is still unclear. Expression of the constitutive isoform of NOS, brain NOS (bNOS), has also been detected both at the mRNA and the protein level in human melanocytes (unpublished observations), and it is possible that α-MSH may also act via this NOS isoform.
One should not exclude the possibility that α-MSH could regulate NO production through a mechanism independent of the MC-1R. As discussed above, there is evidence that α-MSH can act intracellularly to regulate melanogenesis through interactions with 6-BH4 (Schallreuter et al. 1997). 6-BH4 is a co-factor for iNOS and if, as discussed above, α-MSH complexes with this pterin, then this would affect the activity of iNOS and hence NO production.
It is not clear why melanocytes and melanoma cells produce NO. Romero-Graillet et al. (1996,1997) have shown that NO stimulates melanin production and because it is released from keratinocytes in response to UVR they suggested that it could serve as a paracrine factor in UVR-induced melanogenesis. The fact that human melanocytes are capable of producing NO in response to UVR and in higher amounts than those produced by keratinocytes points to the possibility that NO acts as an autocrine factor to regulate melanogenesis.
An alternative possibility is that the NO produced by melanocytes serves as a second messenger in regulating their differentiation, analogous to the situation in neuronal cells. In the latter, the induction of NOS by nerve growth factor causes growth arrest and differentiation (Peunova and Enikolopov 1995). α-MSH has similar effects in melanocytes (Eberle 1988), so it is possible that these effects are mediated by NO. However, as we discuss below the main importance of NO from melanocytes may not be as an autocrine mediator but as a signaling molecule linking the melanocyte with other systems in the skin (Figure 1).
What is the True Role of the Melanocyte?
There is no doubt that melanocytes are able to produce and distribute melanin via their dendrites to surrounding keratinocytes, and hence function as key components of the pigmentary system. However, melanocytes are also able to produce a wide range of signal molecules, such as cytokines (Förster et al. 1991; Armstrong et al. 1992), melanocortin peptides (Wakamatsu et al. 1997), catecholamines (Iyengar and Misra 1987), serotonin (Johansson et al. 1998), and eicosanoids (Okano-Mitani et al. 1997) and, as discussed above, NO (Graham et al. 1997; Tsatmali et al. 2000) (Figure 1). It is not yet clear whether these different secretory products are released from the same melanocyte or from separate populations of melanocytes.
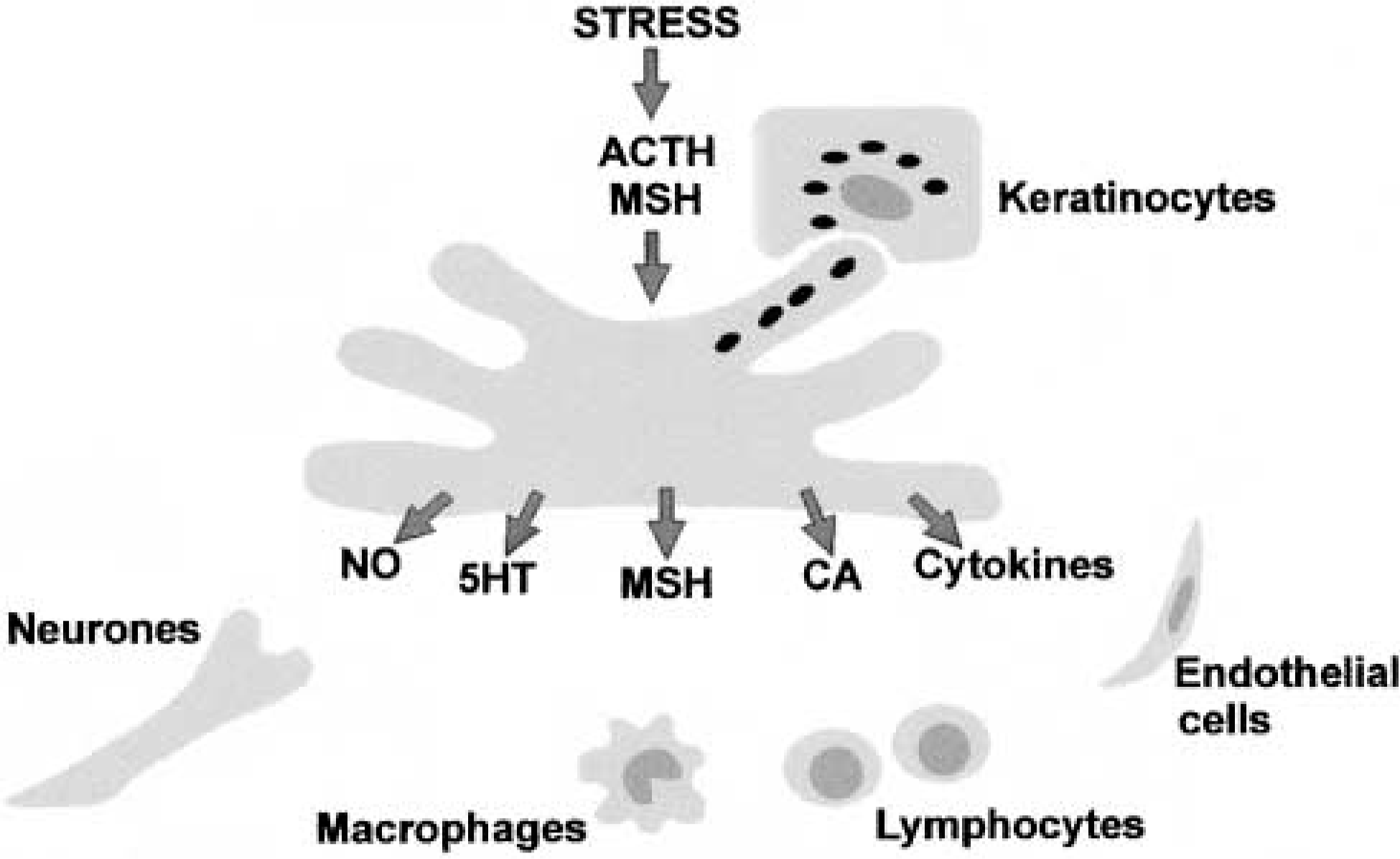
Melanocytes produce a wide variety of substances, including α-MSH, and in this way could regulate many different cell types in the epidermis. α-MSH and ACTH peptides are produced in the epidermis in response to environmental stress and act on melanocytes to regulate melanin and NO production. Melanocortins may also regulate the release of other substances from melanocytes, such as cytokines, catecholamines (CA), and serotonin (5HT).
Nevertheless, it appears that melanocytes are not simply melanin-producing cells and may have some other physiological significance. It has been proposed that melanocytes act as local “stress sensors” in the epidermis (Slominski et al. 1993a,b) and provide communicatory links with several different systems (Figure 1). For example, their close anatomic associations with nerve endings (Hara et al. 1996) and their ability to produce neuropeptides and neurotransmitters suggest a role as a neuroendocrine cell and thus as a key component of a communication pathway between the skin and the central nervous system. Melanocytes could also act as regulators of the skin's immune responses by producing a number of cytokines, including IL-1, IL-6, IL-3, TNFα (Köck et al. 1991,1992) and NO (Graham et al. 1997; Tsatmali et al. 2000). The production of NO could be related to a phagocytic property (Le Poole et al. 1993), which suggests that melanocytes may have a role as accessory cells of the skin's immune system. The fact that melanocytes produce α-MSH could also reflect some role in the modulation of the immune system. There is considerable evidence that α-MSH has potent anti-inflammatory and immunomodulatory properties through its ability to antagonize the actions of proinflammatory cytokines (Catania and Lipton 1993; Watanabe et al. 1993).
It is perhaps significant that several of the substances produced by melanocytes have inflammatory properties. This therefore raises questions as to whether melanocytes might contribute to inflammatory dermatoses and UVR-induced erythema. Because the latter tends to be inversely related to tanning ability, one could speculate that the secretion of inflammatory mediators and the production of melanin by melanocytes are reciprocally related. α-MSH may well have a role in coordinating these responses. Therefore, on the one hand, α-MSH might stimulate melanin production by the melanocyte and, on the other hand, it might act to modulate the production of NO and other inflammatory mediators.
In conclusion it may be some time before we fully appreciate the precise significance of the melanocyte. Nevertheless, it is clear that the melanocyte is more than a melanin-producing cell. The melanocyte is likely to have a number of extra-pigmentary functions and, whatever these functions, it is reasonable to suppose that, like melanin production, they are regulated by the melanocortin signaling system.
Footnotes
Acknowledgements
The financial support of Stiefel International is gratefully acknowledged.