
Abstract
γ-Aminobutyric acid (GABA) is a neurotransmitter that also occurs in a few non-neuronal cell types, where it may serve as a paracrine modulator. GABA is biosynthesized from glutamate by glutamate decarboxylase (GAD) and from putrescine via diamine oxidase (DAO). GAD is demonstrable in several GABA-positive cell types but is undetectable in the GABA-containing gastrin cells and somatostatin cells of the antropyloric mucosa of the stomach. Using two antisera raised against synthetic peptides corresponding to two different regions of rat DAO, we now demonstrate strong reactivity for DAO in gastrinpositive cells of the rat antropyloric mucosa, whereas somatostatin-positive cells as well as other structures of the antrum are unreactive. Western blotting analysis of antrum and colon demonstrate that both antisera react with a single band of 85 kD, consistent with the predicted molecular weight of DAO. Expression of DAO mRNA in the antrum is demonstrated by reverse transcriptase polymerase chain reaction (RT-PCR). Our results strongly indicate that gastrin cells produce GABA via DAO-catalyzed oxidation of putrescine, and experimental data moreover suggest that the biosynthesis of GABA is regulated by the prandial state. Because GABA modulates release of somatostatin, these results point to a new mechanism of paracrine interaction between gastrin cells and somatostatin cells.
Keywords
P
GABA is an inhibitory neurotransmitter that also functions as a neuromodulator in the gastroenteric nervous system, where it occurs mainly in nerves in the myenteric plexus and in circular muscle layers of the muscularis externa (Davanger et al. 1987; Hills et al. 1987). In addition, GABA is detected in certain cell types outside the nervous system, including pancreatic islet β-cells, and somatostatin cells and gastrin cells in the antropyloric mucosa of the stomach (Gilon et al. 1988; Davanger et al. 1994). Its function outside the nervous system is uncertain but lines of evidence indicate that it may act as a paracrine modulator (Harty and Franklin 1983,1986; Erdö and Wolff 1988; Harty et al. 1991; Weigert et al. 1998). Glutamate decarboxylase (GAD) is considered to be the principal enzyme of GABA biosynthesis and has been demonstrated in many GABA-positive cell types in the brain, gastroenteric nervous system, and pancreatic β-cells. However, it has not been possible to demonstrate GAD in antropyloric somatostatin and gastrin cells (Gilon et al. 1991). Because of this, and because feeding and exogenous gastrin inhibit DAO-catalyzed formation of GABA from putrescine in the rat stomach (Hougaard et al. 1992), we have investigated if DAO is present in antropyloric somatostatin and gastrin cells and whether the immunohistochemical staining is affected by the prandial state.
Materials and Methods
Tissue Material
Adult freely fed Wistar rats of either sex were sacrificed with CO2 and intracardially perfused with 1-2 ml of saline, followed by 30 ml of 4% paraformaldehyde in 0.1 M sodium phosphate buffer, pH 7.4 (PFA), or 0.5-2.5% glutaraldehyde with 4% paraformaldehyde in 0.1 M sodium phosphate buffer, pH 7.4. Specimens from the antrum were post-fixed in the respective fixatives overnight, dehydrated in graded ethanol, and embedded in paraffin.
Specimens for Western blotting and RT-PCR were taken from unfixed rats and immediately chilled on ice or frozen in liquid nitrogen, respectively, before further processing.
For quantitative analysis, 18 adult Wistar rats were fasted in wire-bottomed cages with access to drinking water only. After 24 hr, nine rats were sacrificed directly and nine rats were re-fed (in darkness to facilitate eating) with standard rat chow for 3 hr before sacrifice. During sectioning, it was verified that the nine re-fed rats had eaten well and that the stomach of eight of the fasted rats was empty. The stomach of the ninth fasted rat contained hair and other material and was consequently excluded from the study. The remaining 17 rats were processed for immunohistochemistry as described above, using 4% paraformaldehyde as fixative. Paraffin-embedded specimens of antrum were cut longitudinally in sections 5 μm thick and approximately 3-4 mm long. For each of the 17 rats, four sections were cut from different areas of the curvature major region of the antrum and mounted on a single slide.
Diamine Oxidase Antisera
Two groups of five rabbits each were immunized with synthetic peptides (KJ Roos-Petersen; Hörsholm, Denmark) corresponding to the predicted amino acid sequence position 625-639 (SERYSSSLYNQNDPW) (DAO625) and 713-730 (PQDKGLNRVQRWIPEDRR) (DAO713) of rat colon/lung DAO, which is identical to the amiloride binding protein EMBL accession number P36633 (Lingueglia et al. 1993; Chassande et al. 1994; Novotny et al. 1994). The peptide sequences were selected on basis of antigenicity index using the computer program PeptideStructure [Genetic Computer Group (GCG); Madison, WI)]. DAO625 was conjugated to keyhole limpet hemocyanine (KLH) and DAO713 to bovine serum albumin (BSA) by glutaraldehyde. Three ml of ice-cold 0.2% glutaraldehyde in 0.1 M phosphate buffer, pH 7.5, was slowly added to a solution of 3 mg peptide and 3 mg KLH or BSA in 3 ml ice-cold phosphate buffer, pH 7.5. The mixture was rotated for 2 days at 4C and diluted to 9 ml with PBS. Each rabbit was immunized with conjugate corresponding to 100 μg peptide using SC injections at four different locations every second week. For the first immunization, complete Freund's adjuvant (Statens Serum Institut; Copenhagen, Denmark) was used. Subsequent immunizations employed incomplete Freund's adjuvant. After five immunizations, rabbits from both groups produced antiserum useful for immunocytochemistry and Western blotting. One antiserum from each group, designated anti-DAO625 and anti-DAO713, respectively, was selected (see below).
Immunocytochemistry
Five-μm paraffin sections of antrum were dewaxed in xylene, hydrated in graded ethanol, and incubated in 1% nonfat milk or 1% BSA in TBS (0.05 M Tris; 0.15 M NaCl, pH 7.4) for 30 min at RT. Sections were then incubated for 20 hr at 4C with one of the DAO antisera diluted 1:500-1:2000 in TBS. The DAO antisera were absorbed overnight with 200 μg/ml KLH (anti-DAO625) or 0.25% BSA (anti-DAO713) before use. The site of the antigen-antibody reaction was revealed by sequential incubation at RT with biotinylated goat anti-rabbit (DAKO; Glostrup, Denmark) 1:500 in TBS containing 0.25% BSA, 3% rat serum for 1 hr, and alkaline phosphatase-conjugated streptavidin (DAKO) 1:100 in TBS containing 0.25% BSA for 1 hr, followed by development in bromochloroindolyl phosphate-nitroblue tetrazolium (BCIP-NBT) (Sigma; St Louis, MO) medium containing levamisole. Sections stained for GABA were digested with 0-0.04% pepsin (Sigma) in 0.2 M HCl for 20 min at RT before incubation with guinea pig anti-GABA antiserum (Affinity; Surrey, UK) followed by Texas Red-conjugated goat anti-guinea pig IgG (Jackson ImmunoResearch Laboratories; West Grove, PA).
For quantitative analysis of DAO-reactive cells in fasted (n = 8) vs re-fed (n = 9) rats, 17 slides with four sections on each (approximately 12-16 mm antral mucosa per slide) were stained with anti-DAO625 as described above, and the number of DAO-positive nucleated cells per mm length of mucosa was counted. The Mann-Whitney U-test was used for statistical comparisons.
Double immunofluorescence stainings were performed with the rabbit DAO antisera, used in combination with guinea pig antiserum to synthetic human gastrin (Milab; Malmö, Sweden) or with monoclonal mouse antibodies to synthetic somatostatin (Novo Nordisk; NovoClone, Bagsværd, Denmark) (Larsson and Hougaard 1994). The site of the antigen-antibody reaction was detected with species-specific secondary antibodies, conjugated to fluorescein isothiocyanate (FITC) (DAKO) or Texas Red (Jackson), yielding green and red fluorescence, respectively. Sections were examined in a Leica DMR microscope equipped for epi-ilumination using selective FITC and Texas Red filter sets. Acquisition of fluorescence images was performed with a Leica Q550CW workstation.
Crossreactivity of secondary antibodies with relevant primary antibodies was ruled out by separate staining experiments. Controls also included preabsorption of primary antisera overnight against 150 μg/ml of synthetic DAO peptides (positions 625-639 and 713-730), gastrin, or somatostatin (Peninsula Laboratories; Belmont, CA), as well as conventional staining controls as described (Larsson 1988).
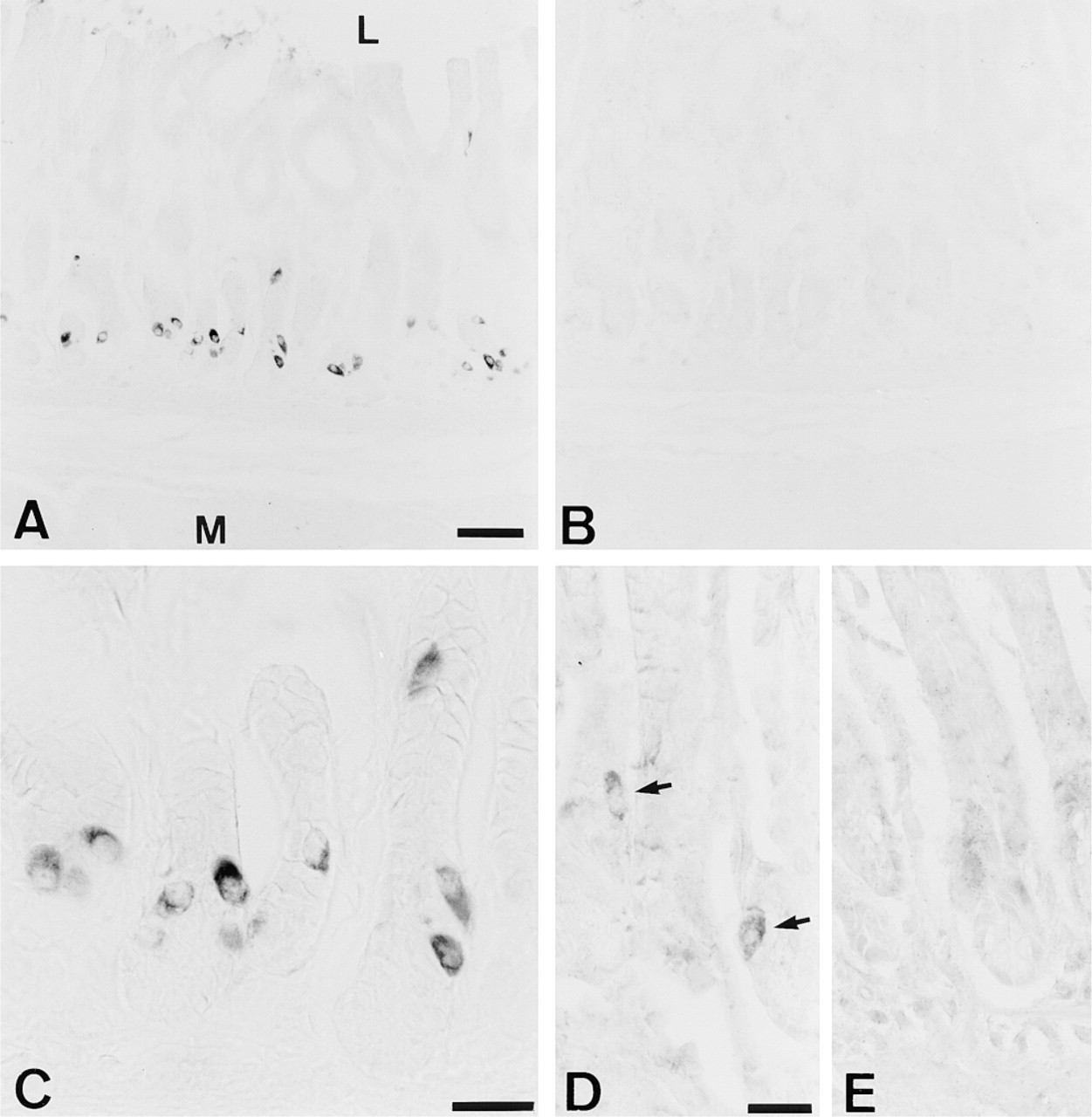
Sections of rat antrum immunohistochemically stained with (
Western Blotting
Homogenized samples of antrum and colon were sonicated in a water bath at 30C for 5 min, boiled for 5 min, and loaded on a 7.5% polyacrylamide gel. The sample buffer was 62.5 mM Tris-HCl, pH 6.8, 0.7 M 2-β-mercaptoethanol, 10% (v/v) glycerol and 2% (w/v) SDS (Bio-Rad; Hercules, CA). After electrophoresis, the proteins were transferred to a nitrocellulose membrane (Millipore; Bedford, MA) using a semi-dry cell (Pharmacia Biotech; Uppsala, Sweden). The membrane was incubated overnight with 3% nonfat milk in PBS (PBS/milk) at 4C, washed in PBS containing 0.1% Tween-20 (Merck; Darmstadt, Germany) (PBS/ Tween), and incubated overnight with the DAO antisera diluted 1:10,000 in PBS/milk at 4C. Detection was achieved by biotinylated goat anti-rabbit (DAKO) 1:1000 in PBS/ milk, followed by alkaline phosphatase-conjugated streptavidin (DAKO) 1:200 in PBS/milk, using bromochloroindolyl phosphate (BCIP) and nitroblue tetrazolium (NBT) (Sigma) as substrate. Controls included preabsorption of antisera overnight against 150 μg/ml of the synthetic peptides.
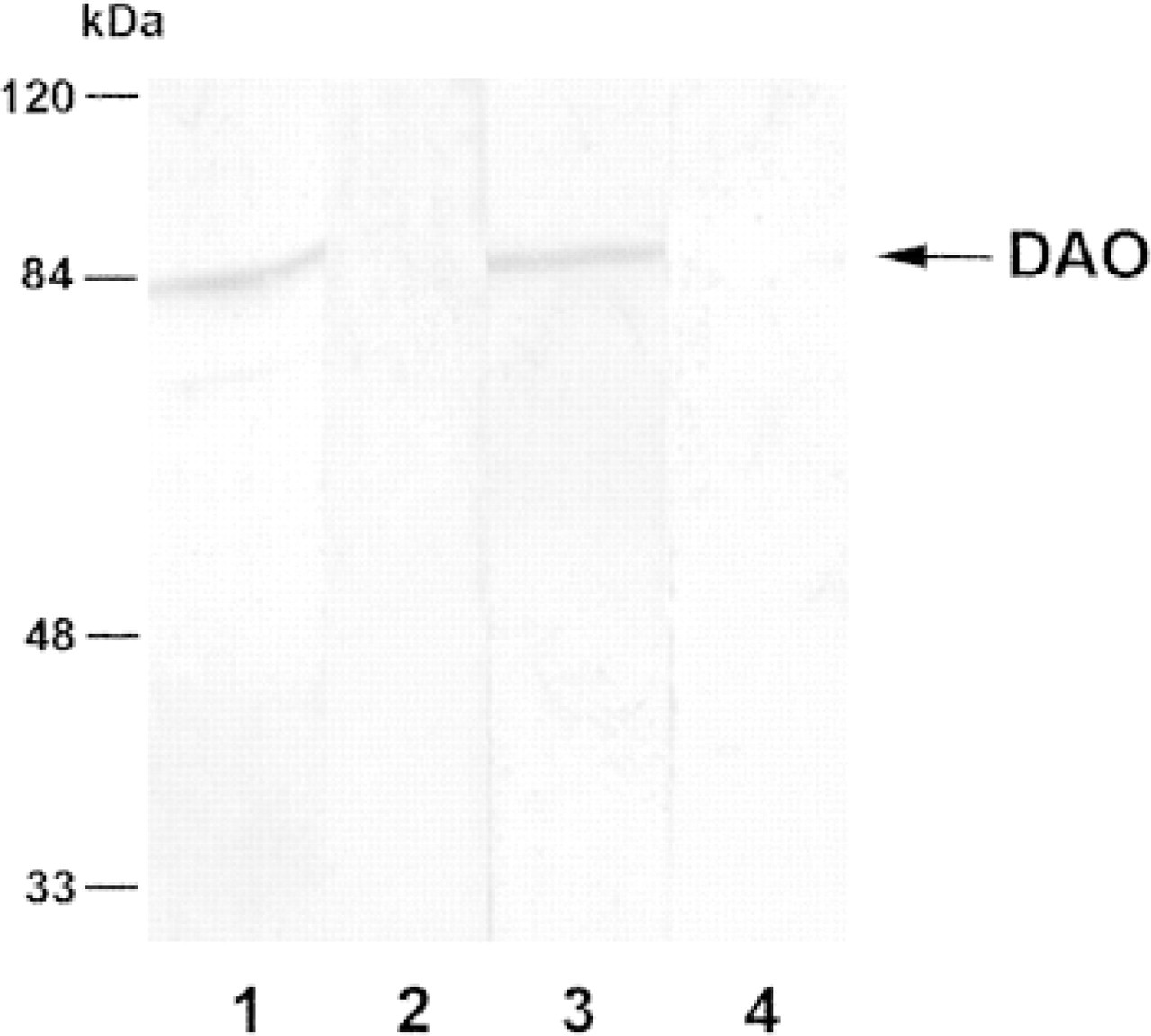
Western blots of rat antrum. Lane 1 is reacted with anti-DAO713, Lane 2 with anti-DAO713 preabsorbed with synthetic peptide DAO713, Lane 3 with anti-DAO625, and Lane 4 with anti-DAO625 preabsorbed with peptide DAO625. Both antisera demonstrate a single band at 85 kD protein, corresponding to the molecular weight of DAO, and staining is abolished if the antisera are preabsorbed with their corresponding peptides.
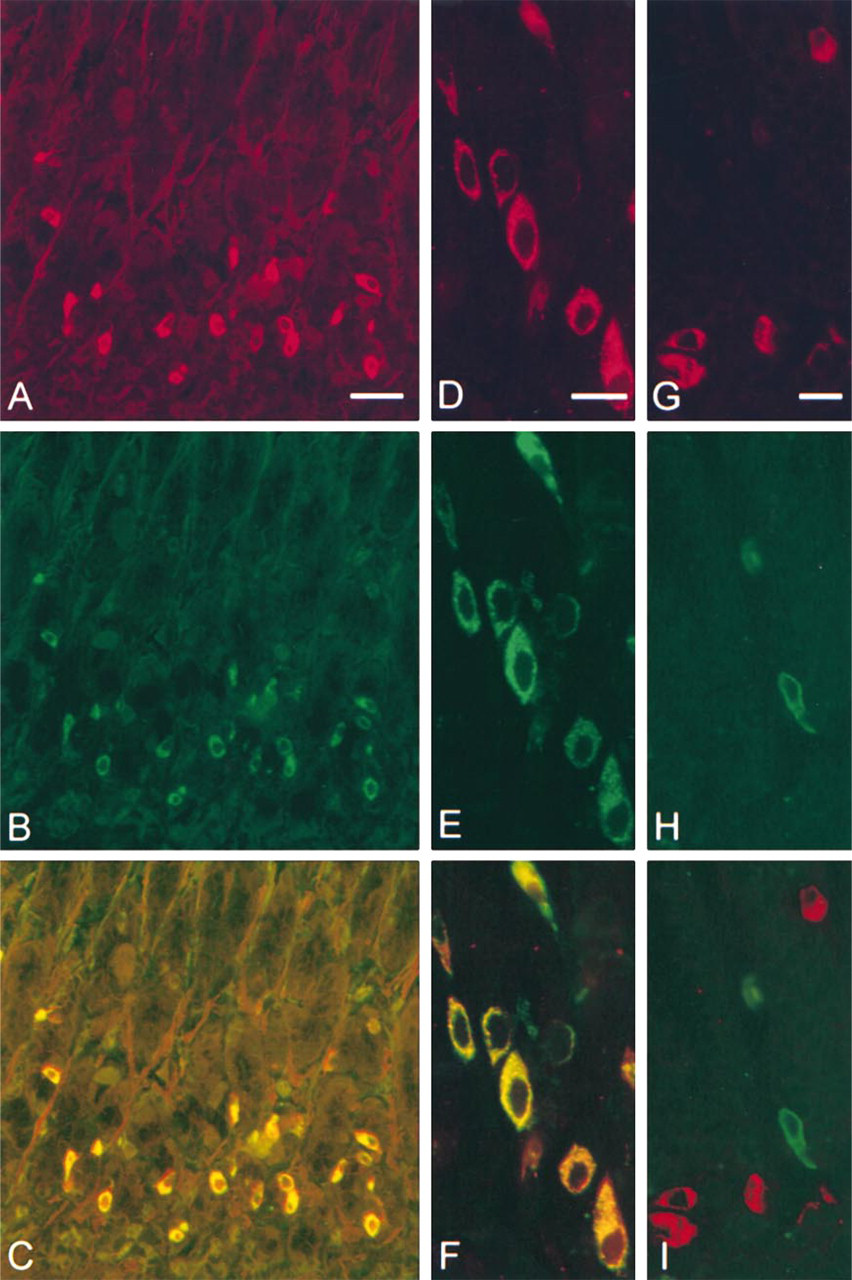
Sections of rat antropyloric mucosa double immunofluorescence stained for diamine oxidase/gastrin and for diamine oxidase/ somatostatin. (
Reverse Transcriptase Polymerase Chain Reaction (RT-PCR)
Total RNA from rat antropyloric mucosa and colon was isolated using TRIzol Reagent (Life Technologies; Grand Island, NY). cDNA was produced employing random hexamers and reverse transcriptase from SuperScript (Life Technologies; cat. no. 18089-011). Primers used in the polymerase chain reaction (PCR) were DAO upper primer, position 1629-1649 and lower primer, position 2123-2143, EMBL mRNA sequence accession no. X73911, and β-actin upper primer, position 2394-2415, and lower primer, position 3088-3106, GeneBank DNA sequence accession no. J00691 (primers spanning two introns of total 212 bp).
Amplicons were electrophorized in a 1.5% agarose gel and occasionally transferred to a positively charged nylon membrane (Boehringer Mannheim; Mannheim, Germany) in 0.5 M NaOH, 0.15 M NaCl by vacuum blotting (Vacu-Aid; Hybaid, Teddington, UK). The membrane was heated at 80C for 30 min, exposed to UV radiation for 2 min and prehybridized in hybridization buffer containing 2 × SSC (0.3 M NaCl and 0.03 M sodium citrate), 0.2 mg Ficol (type 400; Sigma), 0.2 mg polyvinylpyrrolidone (MW 25,000; Merck), 0.2 mg BSA, 100 mg dextran sulfate (MW 500,000; Sigma), and 0.1 μg sheared salmon sperm DNA (Sigma) for 2 hr at 60C (Hougaard et al. 1997). A chemically biotinylated oligodeoxynucleotide probe (Hougaard et al. 1997) complementary to rat DAO mRNA (position 1878-1907, EMBL accession no. X73911) was added to the hybridization buffer to a final concentration of 5 ng/ml and hybridized overnight at 60C. The membrane was washed twice for 15 min in 2 × SSC, 0.1% SDS, and twice for 15 min in 0.5 × SSC, 0.1% SDS at 50C before it was blocked in 3% BSA, 0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl for 1 hr at 60C. Alkaline phosphatase-conjugated streptavidin was added to the blocking buffer to a final dilution of 1:1000 and the membrane was incubated for 10 min at RT. Alkaline phosphatase activity was visualized by BCIP-NBT development for 20 min.
Results
The two antisera, raised against different regions of rat DAO (anti-DAO625 and anti-DAO713), stained endocrine-like cells in the antropyloric mucosa. Anti-DAO625 produced a strong, distinct staining confined to the cytoplasm of several endocrine-like cells (Figures 1A and 1C). Preabsorption of anti-DAO625 against its corresponding synthetic peptide DAO625 abolished the staining (Figure 1B). Anti-DAO713 produced a considerably weaker staining of the endocrine-like cells and a diffuse non-absorbable, and hence unspecific, background staining in many structures (Figures 1D and 1E). Preabsorption of the antisera against unrelated peptides was without effect. No other specific staining was observed in the mucosa or in the submucosa and muscularis externa. Notably, neuronal cells of the submucous plexus and the myenteric plexus present in muscularis externa were unreactive. It was not possible to obtain staining in the antropyloric mucosa with the DAO antisera when glutaraldehyde was included in the fixative, whereas, in contrast, glutaraldehyde was required for a positive staining with the GABA antiserum (not shown).
The specificity of the DAO-antisera was further corroborated by Western blotting analysis of antrum and colon. Both antisera detected a single band of 85 kD that was not detected by sera preabsorbed with their corresponding peptides (Figure 2). Because the anti-DAO625 produced the strongest specific staining and the lowest background, it was selected for further analysis.
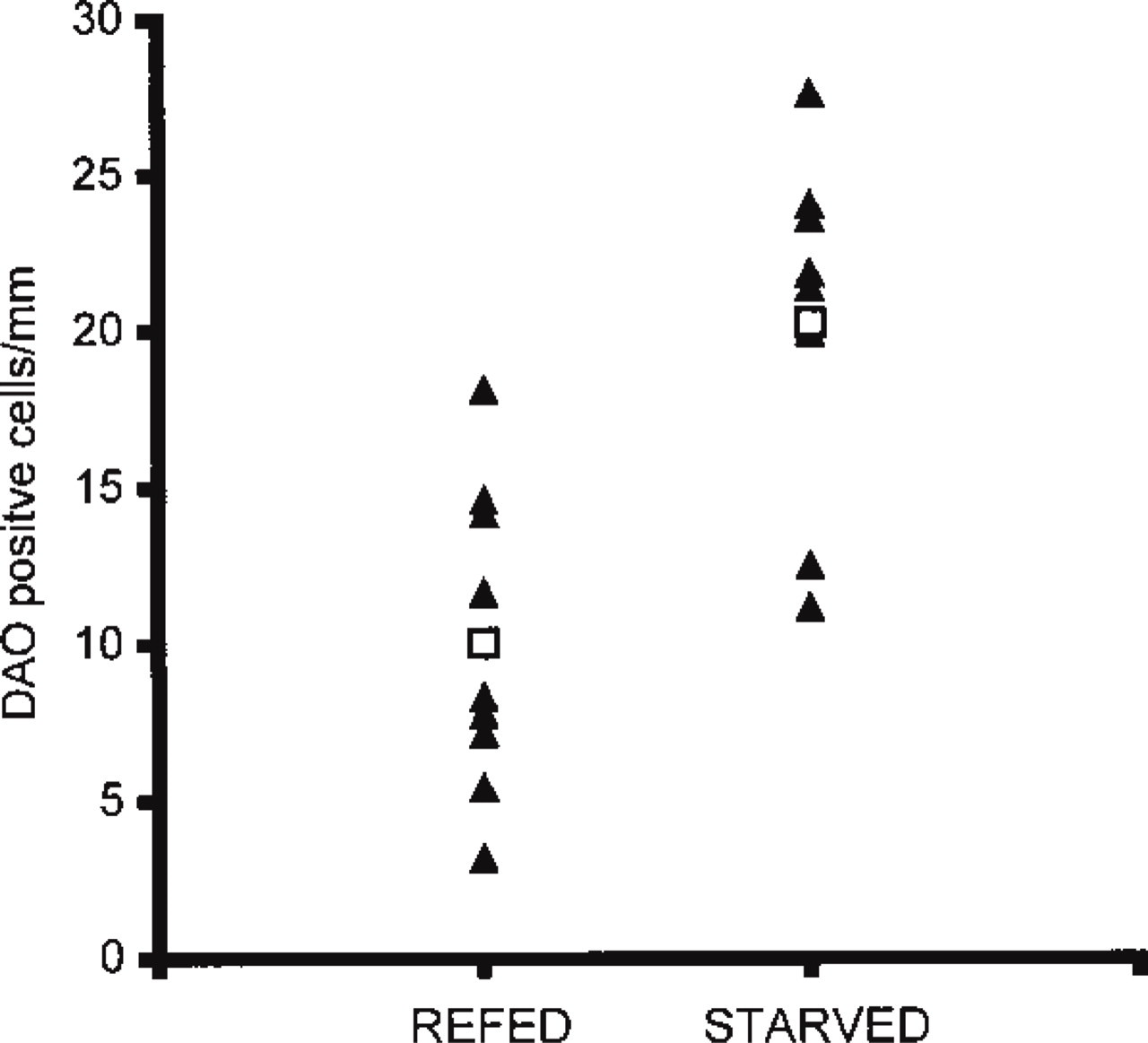
DAO-immunoreactive cells (anti-DAO625) per mm length of antropyloric mucosa in re-fed (n = 9) and fasted (n = 8) rats. ▴, individual rats; □, mean value in each group. The frequency of DAO-positive cells is significantly higher in the group of fasted rats (p<0.01, Mann-Whitney U-test).
Examination of antral sections from more than 10 individual rats stained by the double immunofluorescence technique, using anti-DAO625 and guinea pig anti-gastrin, revealed that cells positive for DAO (red) were also positive for gastrin (green) (Figures 3A-3F). In general, cells strongly reactive for one of the compounds were also strongly reactive for the other (composite color bright yellow) (Figure 3C). However, some cells stained with different intensities for DAO and gastrin, yielding composite colors ranging from orange to yellow. This variation in staining intensity was most clearly seen at the subcellular level, where compartments that stained orange, yellow, and green could be observed (Figure 3F). Double immunofluorescence staining for DAO and somatostatin, using anti-DAO625 and mouse anti-somatostatin as primary antibodies, revealed no reactivity for DAO in somatostatin-positive cells (Figures 3G-3I). Single staining experiments, systematically combining the different primary antisera with each of the fluorescence-labeled secondary antisera documented that all secondary antibodies reacted only with the primary antibody intended.
The frequency of antropyloric cells containing detectable amounts of DAO depended on the prandial state and varied among individual rats. For each rat, the total length of antral mucosa examined was 12-16 mm. In rats sacrificed immediately after 24 hr of fasting, the number of anti-DAO625-reactive cells ranged from 11.2 to 27.6 per mm length antropyloric mucosa, mean value 20.3 cells (Figure 4). After 3 hr of re-feeding, the number of cells detectable with anti-DAO625 was significantly lower (p<0.01), ranging from 3.2 to 18.1 cells per mm mucosa, mean value 10.1 cells.
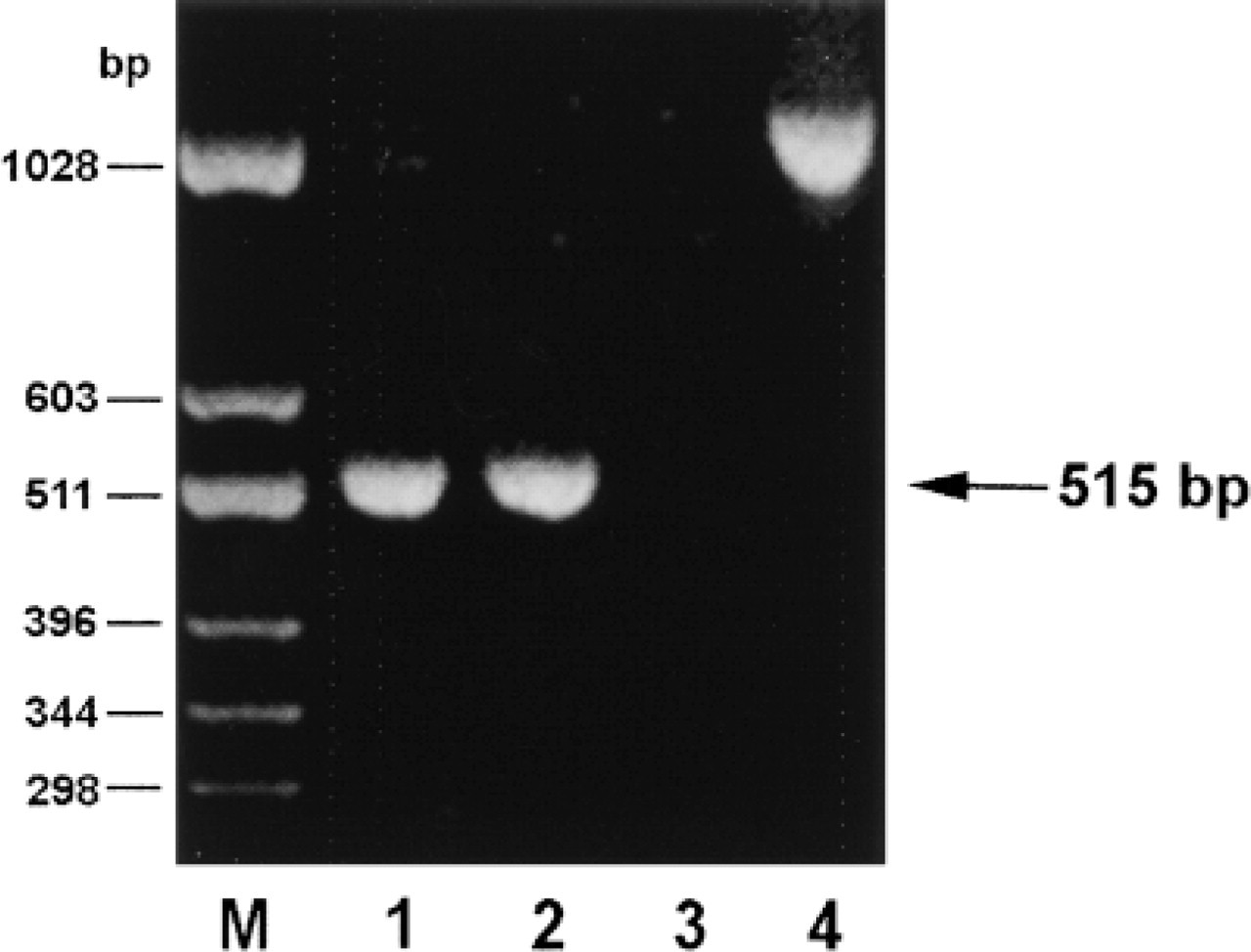
Expression of DAO mRNA in rat antrum and colon analyzed by RT-PCR. Lane 1, colon; Lane 2, antrum; Lane 3, antrum without reverse transcriptase. Lane 4, a PCR fragment produced by amplification of chromosomal DNA using the same DAO primer set. Note expression of DAO mRNA in antrum and colon represented by 515-bp amplicons, and that amplification of genomic DNA produces an approximately 1050-bp fragment.
The expression of DAO mRNA in the antrum was demonstrated by RT-PCR. Amplicons of the sizes predicted for rat DAO mRNA (515 bp) were produced using cDNA from the antropyloric mucosa and colon as template (Figure 5). No other bands appeared. The identity of the DAO amplicons was verified by Southern blotting. PCR using rat DNA as template produced an amplicon at 1045 bp, indicating that the DAO primers are spanning one or more introns of a total of approximately 530 bp (Figure 5). Possible contamination of the mRNA preparations with genomic DNA was also ruled out by omission of reverse transcriptase during preparation of cDNA, which abolished subsequent production of amplicons, and by control PCR using primers for β-actin spanning two introns of 212 bp total.
Discussion
Our results demonstrate that DAO is present in antropyloric gastrin cells. Both antisera recognizing DAO stain the cytoplasm of gastrin-producing cells, whereas antropyloric somatostatin cells and neuronal cells of the myenteric and submucous plexi are negative. The localization of DAO in gastrin cells is supported by Western blotting analysis and RT-PCR demonstrating the presence of DAO and DAO mRNA in the rat antrum.
The two antisera produce identical results on Western blots, but anti-DAO625 is preferred for immunohistochemistry because anti-DAO713 produces a disturbing diffuse unspecific background staining of the sections.
Double immunofluorescence staining demonstrates cellular co-localization of gastrin (green) and DAO (red). The resulting composite colors range from orange to bright yellow, indicating that the relationship between the concentration of the two compounds varies, and hence that the cellular level of gastrin and DAO most probably is regulated by separate mechanisms.
Because of different requirements for tissue fixation, it was not possible to stain for DAO and GABA simultaneously. However, it has previously been shown that gastrin-producing cells in the rat antrum contain GABA, predominantly in the extragranular cytoplasmic matrix (Davanger et al. 1994). GABA has also been demonstrated in two other antral cell types, classified as somatostatin cells and serotonin-producing ECn cells (Oomori et al. 1992; Davanger et al. 1994). It can therefore be concluded that DAO and GABA coexist in gastrin cells but not in somatostatin cells. Neither gastrin nor somatostatin cells contain detectable amounts of GAD (Gilon et al. 1991). Consequently, GABA in gastrin cells is most probably produced via DAO-catalyzed oxidation of putrescine, whereas GABA in somatostatin cells may have another origin. This hypothesis is well in line with the observations that somatostatin cells, but not gastrin cells, are capable of taking up exogenous GABA (Gilon et al. 1990). The physiological function of GABA in the antropyloric mucosa is unresolved, but several reports indicate that it may play an active regulatory role. Evoked release of GABA has been demonstrated from a presumably non-neuronal antral pool, and exogenous GABA decreases somatostatin and increases gastrin release from the stomach in vitro (Harty and Franklin 1983,1986; Erdö and Wolff 1988; Harty et al. 1991; Weigert et al. 1998). In many species, including rat, somatostatin and gastrin cells contact each other via slender cytoplasmic processes, and there is ample evidence indicating that local paracrine release of somatostatin inhibits production and secretion of gastrin (Larsson et al. 1979; Saffouri et al. 1979; Larsson 1980,1984; Karnik and Wolfe 1990; McIntosh et al. 1991; Larsson and Hougaard 1994). Therefore, the increase in gastrin secretion after exogenous GABA may be a consequence of decreased release of somatostatin.
Gastric mucosal growth, differentiation, and function are dependent on gastrin (Konturek et al. 1995; Koh et al. 1997; Langhans et al. 1997). The secretion of gastrin is increased by feeding, and a basal secretion is maintained during fasting. Feeding of rats with specific gastric nutrients increases antral gastrin mRNA levels and decreases somatostatin mRNA levels, whereas fasting has the opposite effect (Wu et al. 1991). Our present results show that the numbers of DAO-immunoreactive cells are increased during fasting, which agrees with studies showing that fasting increases DAO-catalyzed formation of GABA from putrescine in the rat stomach (Hougaard et al. 1992). It is therefore tempting to speculate that gastrin cells may relieve the tonic restraint on gastrin secretion by paracrine release of GABA, which would serve to inhibit the secretion of somatostatin.
Footnotes
Acknowledgements
Supported by the Danish Cancer Society and the Danish MRC.