
Abstract
The hereditary disease cystic fibrosis (CF) is caused by mutations in the cystic fibrosis transmembrane conductance regulator (CFTR) gene. Understanding of the consequences of CFTR gene mutations is derived chiefly from in vitro studies on heterologous cell cultures and on cells hyperexpressing CFTR. Data from ex vivo studies on human tissue are scarce and contradictory, a fact which is in part explained by secondary tissue destruction in most affected organs. The purpose of this study was to establish conditions under which wild-type and mutated CFTR can be studied in affected human tissue. Sweat glands carry the basic defect underlying CF and are not affected by tissue destruction and inflammation. Therefore, we used this tissue to test a panel of eight different CFTR antibodies under various fixation techniques. The antibodies were tested on skin biopsy sections from healthy controls, from CF patients homozygous for the most common mutation, ΔF508, and from patients carrying two nonsense mutations. Of the eight CFTR antibodies, only three—M3A7, MATG 1104, and cc24—met the criteria necessary for immunolocalization of CFTR in sweat glands. The labeling pattern in the CF sweat glands was consistent with the postulated processing defect of ΔF508 CFTR. The antibodies exhibited different sensitivities for detecting ΔF508 CFTR.
C
In contrast to the bulk of evidence from in vitro studies, data from in vivo experiments are scarce. Direct studies on affected human tissue are hampered by several obstacles. First, the cell defect of CF causes tissue destruction in affected organs such as the airways, pancreas, and biliary system. Second, CFTR protein appears to be expressed at low densities (Trezise and Buchwald 1991; Kartner et al. 1992). Antibodies capable of detecting CFTR in CFTR-hyperexpressing cells may not be sensitive enough to detect CFTR in human tissue. Finally, the complex nature of the CFTR protein has rendered the production of antibodies extremely laborious. The epitopes recognized by the antibodies frequently become inaccessible after routine tissue preservation (Kartner et al. 1992; Hanaoka et al. 1996).
Despite these difficulties, mislocalization of ΔF508 CFTR was demonstrated in the sweat gland (Kartner et al. 1992) and in airway epithelium (Puchelle et al. 1992). However, the concept of mislocalization of ΔF508 CFTR in airways has been challenged by the finding that epithelial remodeling had a stronger influence on the subcellular CFTR localization than the CFTR genotype (Brézillon et al. 1995; Dupuit et al. 1995). Moreover, a recent study on several tissues from CF patients and controls showed that the trafficking defect of ΔF508 CFTR was tissue-dependent (Kälin et al. 1999). Therefore, the findings from CFTR-hyperexpressing cells do not seem to reflect the in vivo situation. Clearly, more in vivo studies on human tissue are mandatory to reach an understanding of the impact of the most common CFTR gene mutation on the subcellular localization and, eventually, on cellular function of the protein product. A question of major concern is the extent to which different techniques and antibodies influence the differing findings of the ex vivo analyses.
The purpose of this study was to compare the different fixation techniques and antibodies as applied in the studies mentioned above and to determine the value and specificity of the CFTR antibodies commonly in use for detection of CFTR in sweat glands.
Materials and Methods
Tissue Samples
Full-thickness skin biopsies were taken from the right shoulder of six healthy volunteers, four ΔF508 homozygous CF patients, and four patients bearing two nonsense mutations within the CFTR gene (G542X/G542X; n=2; R553X/R553X; and G542X/W1282X). Informed consent was obtained from all volunteers; approval of the local ethical committee was obtained for the procedure.
CFTR Antibodies
MATG monoclonal antibodies (MAbs) were from Transgène (Strasbourg, France) with CFTR epitopes at amino acids 107-117 (MATG 1016, IgG2a and MATG 1031, IgG1 unpurified), amino acids 503-515 (MATG 1061, IgG2a), and amino acids 722–734 (MATG 1104, IgG1). The MATG 1061 and 1104 antibodies were purified by affinity chromatography on protein G. MAb G1660, purified by DEAE fast flow column chromatography, recognizes a CFTR epitope within the R domain at amino acids 729–736. MAb G2503, purified by protein A-Sepharose column chromatography, recognizes a CFTR epitope at the C-terminus, amino acids 1466-1480. Both MAbs were from Genzyme (Cambridge, MA).
Unpurified MAb M3A7 was made available by Dr. J. R. Riordan (Mayo Clinic; Scottsdale, AZ). It was raised against the region from the second nucleotide binding fold to the C-terminus of CFTR (amino acids 1195-1480).
The cc24 affinity-purified polyclonal rabbit antibody was donated by Dr. A. Nairn (Rockefeller University; New York, NY), and recognizes a CFTR epitope in the R-domain at amino acids 693–716.
IgG concentrations and the corresponding dilutions used are given in Table 1.
Immunohistochemistry
Biopsy specimens were cryofixed in liquid nitrogen. Four-μm cryosections were air-dried, fixed in acetone for 10 min at room temperature (RT) and stored at −70C until use. Some sections were additionally fixed for 5 min at −20C in methanol, as indicated.
Immunohistochemical analysis was performed using the enhanced enzymatic alkaline phosphatase-anti-alkaline phosphatase (APAAP) method (Cordell et al. 1984). Endogenous enzyme activities were blocked with 1.7 mM levamisol. Negative controls were performed in parallel for specific labelings by replacing the primary antibody with the isotype control or nonimmune serum, or by peptide competition. For this, 1 μg of CFTR antibody in the dilution used was preincubated for 1 hr at RT with an excess of the peptide antigen (5 μg).
Results
A pilot study on paraffin-embedded skin sections yielded no sweat gland labeling with any of the eight antibodies against CFTR. Therefore, the antibodies were tested on cryofixed skin sections. Fixation techniques were modified by fixing the air-dried sections solely in acetone or subsequently in methanol at −20C, as frequently practiced with MATG antibodies. The results of all CFTR antibodies are summarized in Table 1.
Labeling in Acetone-fixed Control Skin Sections
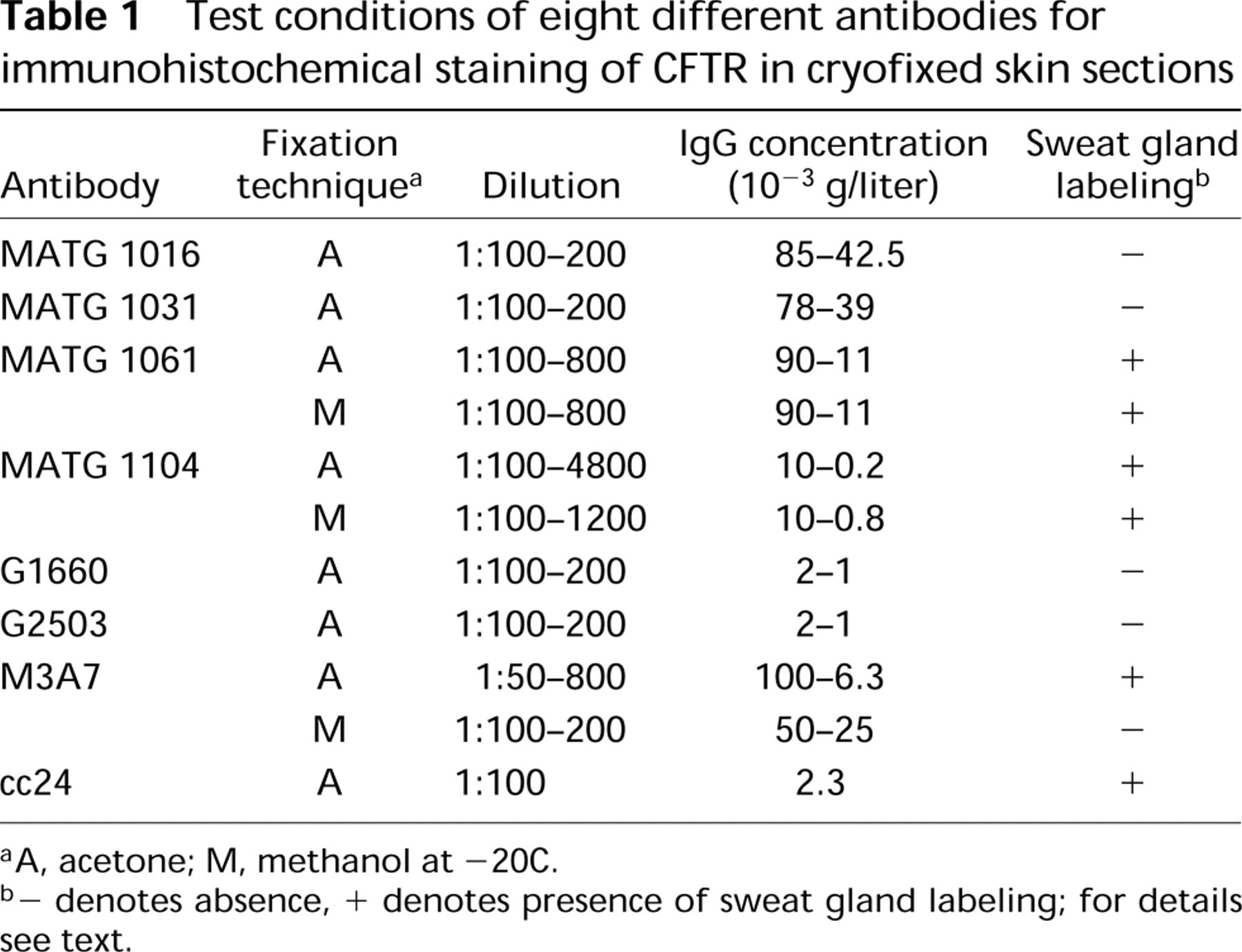
The CFTR antibodies were first tested on acetone-fixed skin sections of healthy controls. Signals were observed with the CFTR antibodies MATG 1061, MATG 1104, M3A7, and cc24. The results are shown in Figures 1A and 1D. M3A7 stained only the reabsorptive epithelium of sweat glands. Intensive labeling of the luminal surface of the duct epithelium was noted but also of the perinuclear regions of basal and luminal duct cells. The secretory cells were negative as were all other structures of the skin sections. Occasionally, weak IgG-crossreactive material was observed in the secretory cells. The results with the MATG 1104 antibody were comparable, provided it was applied at final concentrations below 10−3g/ liter (corresponding to dilutions above 1:1000). At higher concentrations, crossreactive material in the connective tissue was observed, but epithelial labeling was absent. With cc24, similar signal patterns were obtained. The antibody gave some additional background staining of all epithelial cells, including the whole sweat gland, but clearly distinguished from the intense sweat duct labeling. MATG 1061 differed from these three antibodies in that it preferentially stained the secretory coils and, to a lesser extent, the reabsorptive ducts of the sweat glands. Competition experiments were performed with MATG 1104 and cc24. Sweat duct labeling was suppressed by the specific antigenic peptides (Figure 2). With the antibodies G1660, G2503, MATG 1016, and MATG 1031, no signals above background level were observed.
Test conditions of eight different antibodies for immunohistochemical staining of CFTR in cryofixed skin sections
aA, acetone; M, methanol at −20C.
b- denotes absence, + denotes presence of sweat gland labeling; for details see text.
Labeling in Methanol-fixed Control Skin Sections
The antibodies were subsequently tested on skin sections that were fixed in methanol at −20C (Figure 3). All antigenic sites, as detected by M3A7 were completely abolished by this procedure. The labeling by MATG 1061 was altered in that strong luminal signals were observed in both the secretory and reabsorptive sweat gland epithelium. However, labeling was not confined to the luminal membranes but extended to intraluminal material. Crossreactive material was also observed in the connective tissue, most prominent in the direct periglandular region. The same signal pattern was observed with the MATG 1104 antibody on methanol-fixed tissue, irrespective of antibody concentration.
Labeling in ΔF508 CF Patients' Skin Sections
Skin sections of CF patients were examined using the four antibodies that had proved to produce labeling of control sweat glands (Figure 4). M3A7, MATG 1104, and cc24 were incubated on acetone-fixed sections, whereas MATG 1061 was tested on methanol-fixed sections.
M3A7 and MATG 1104 failed to label sweat glands from patients homozygous for the ΔF508 deletion. In contrast, intracytoplasmic staining was clearly observed above background level in the reabsorptive sweat ducts after incubation with cc24. A distinct zone at the luminal sweat duct membranes remained unlabeled (Figure 4C). MATG 1061 produced the same staining pattern as in control tissue.
Labeling in Skin Sections from CF Patients with Nonsense Mutations in the CFTR Gene
All four antibodies labeled sweat glands in skin sections. However, the results on cellular localization, on the labeling of secretory epithelial cells, and on the labeling of ΔF508 sweat glands were contradictory. To address these issues, the antibodies were tested on skin sections from patients with nonsense mutations in the CFTR gene. In these patients, the mutations cause CFTR protein synthesis to cease prematurely. Figure 5 shows that M3A7 and MATG 1104 produced no labeling in the sweat glands from these patients, and cc24 gave only the background staining that was noted in all epithelial cells. However, the MATG 1061 antibody labeled luminal epitopes in sweat glands and connective tissue. The signals obtained with this antibody in skin sections must therefore be considered to be unspecific.
Thus, we were able to select three different antibodies for immunohistochemical detection of wildtype CFTR in sweat glands. However, the sensitivity in detecting ΔF508 CFTR differed for the three antibodies.
Discussion
We have undertaken a study on the feasibility of CFTR antibodies currently in use for immunohistochemistry on human tissue. Skin sections were chosen for three reasons. First, they are easy to obtain, with minimal discomfort for patients. Second, they contain structures that are expected and have been shown to express CFTR—the sweat glands—as well as structures free of CFTR, which can serve as internal controls (e.g., blood vessels, nerves). Finally, the structure of the sweat gland is not affected by secondary effects such as inflammation and remodeling. We have examined eight CFTR antibodies under two fixation techniques, including those that were used in the studies by Kartner et al. (1992), Dupuit et al. (1995), and Kälin et al. (1999).
The Genzyme antibodies G1660 and G2503 are commercially available and have been used for immunocytochemical analysis in CFTR-hyperexpressing cells (Gregory et al. 1990; Denning et al. 1992). In the initial descriptions, extremely high antibody concentrations were applied on single cells. Such concentrations are not attainable in human tissue because of serious background staining. In our hands, these antibodies produced no specific labeling in skin sections at the concentrations tested (Table 1).
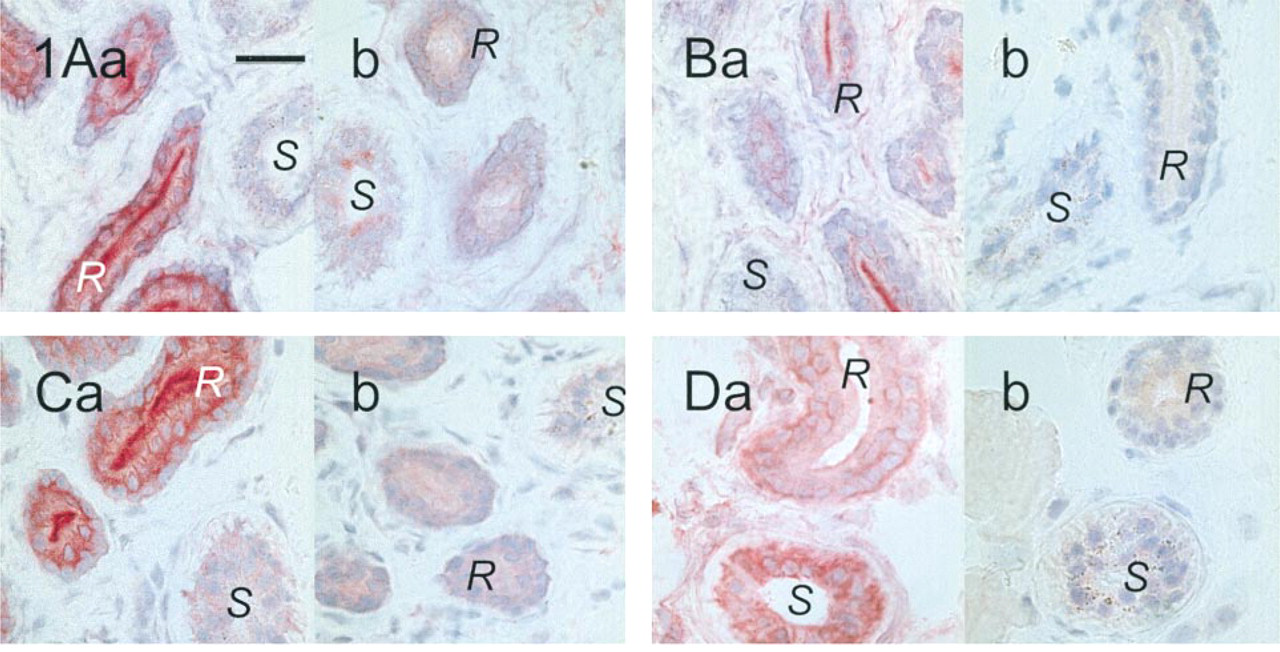
Labeling of sweat glands in acetone-fixed control skin sections. Antibodies with M3A7 (1:200; (
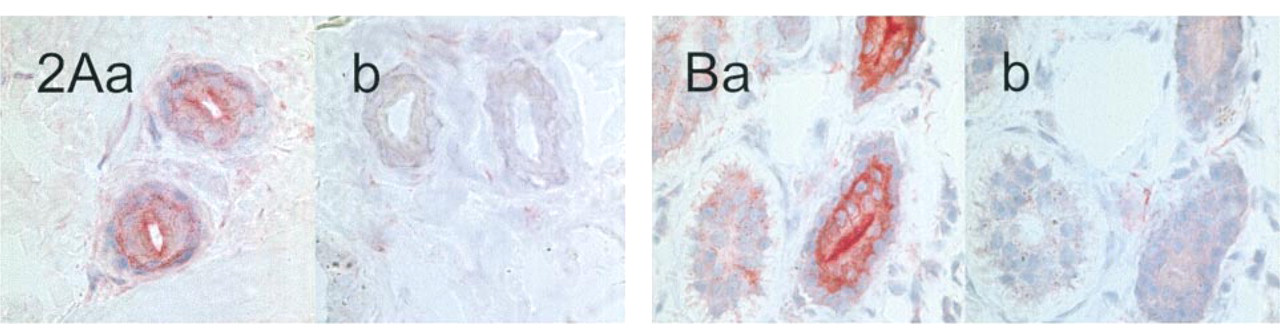
Labeling of sweat glands in acetone-fixed control skin sections. Competition experiments with MATG 1104 (1:2400; (
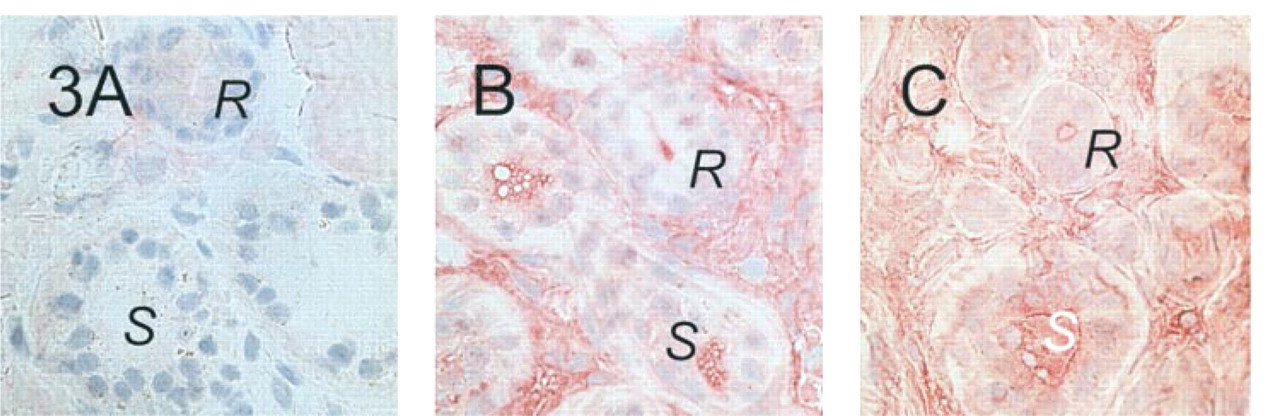
Labeling of sweat glands in methanol-fixed control skin sections. No signal is obtained with M3A7 (1:200;
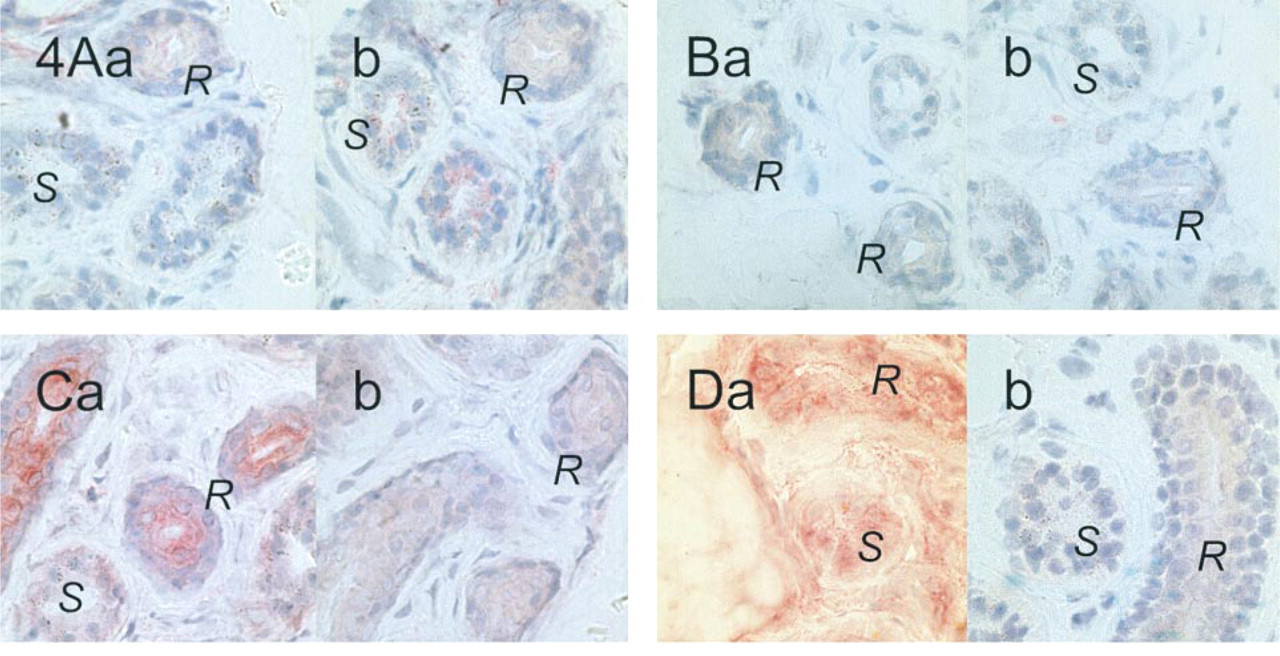
Labeling of sweat glands in ΔF508 homozygous CF patients' skin sections. M3A7 (1:200;
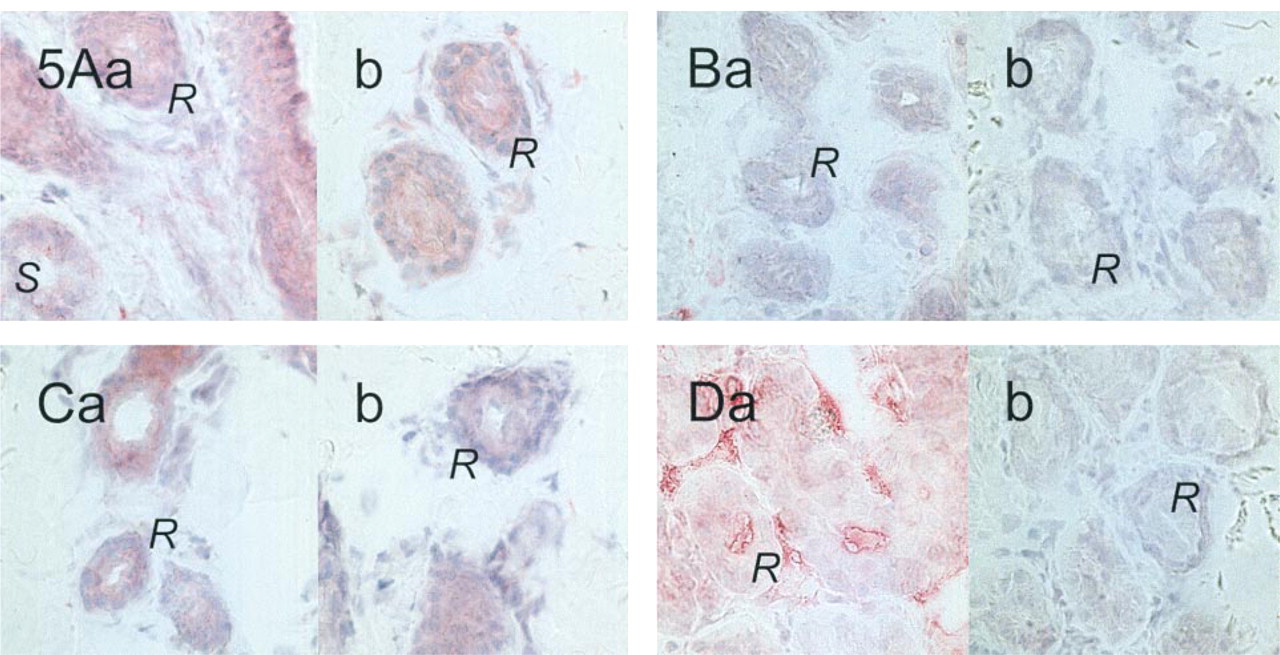
Labeling of sweat glands from CF patients carrying two nonsense mutations. M3A7 (at very low dilution of 1:50;
A variety of CFTR antibodies that were raised against defined oligopeptides can be obtained through Transgène. The MATG 1016 and MATG 1031 antibodies are directed against the extracellular loop of the first transmembrane domain of CFTR (Dalemans et al. 1992; Demolombe et al. 1994). They have mainly been used in cell cultures but obviously did not reach its epitope in the skin sections. The MATG 1061 and MATG 1104 antibodies were applied in several studies on airway epithelium (Puchelle et al. 1992; Brézillon et al. 1995; Dupuit et al. 1995). Specific labeling in skin sections was obtained with the MATG 1104 antibody, as demonstrated by immunological (nonimmune isotype IgG1 and peptide competition studies) and biological (CFTR nonsense mutations) negative controls and by conformity with literature data (Kartner et al. 1992; Cohn et al. 1991).
In contrast, we were not able to achieve specific labeling by MATG 1061. In an attempt to reconcile these results with the data from airway epithelium, we added a methanol fixation step according to the literature protocols (Kälin et al. 1999). Unexpectedly, luminal labeling was then observed in the secretory and the reabsorptive coils. Because the standard immunological controls were negative, we could not decide whether MATG 1061 was more sensitive for detection of CFTR in the secretory coil as well or whether it bound to crossreactive material. We addressed this issue by looking for individuals homozygous for nonsense mutations. In these mutations, a stop codon leads to premature arrest of transcription and thereby to the expression of no protein or a truncated protein. Although overreading of stop codons has been reported to occur in mammalian cells (McCaughan et al. 1995), ample evidence exists that the nonsense mutations R553X and G542X of the CFTR gene result in reduced to undetectable levels of mRNA transcripts (Hamosh et al. 1992; Will et al. 1995) and absence of full-length protein (Howard et al. 1996). With respect to the W1282X mutation, conflicting data are reported in the literature. mRNA levels were found to be severely decreased by Will et al. (1995) and Hamosh et al. (1992) but within normal range by Shoshani et al. (1994). However, expression of full-length protein has never been demonstrated under normal conditions in this stop mutation (Bedwell et al. 1997).
Tissue from CF patients carrying two nonsense mutations can therefore be expected to represent perfect “biological” negative controls. We could trace five patients in Germany, of whom four were willing to cooperate in the study. This biological negative control clearly showed that the MATG 1061 labeling was nonspecific for CFTR in the sweat glands. These results stress the importance of valid controls. Methodically negative controls, including the competition studies, were not sufficient to demonstrate or rule out the specificity of the labeling. M3A7 was produced by Dr. N. Kartner and is directed against a large epitope ranging from the second nucleotide binding fold to the C-terminal end of CFTR. Its specificity in detecting CFTR in sweat glands has been documented in the studies by Kartner et al. (1992) and by Kälin et al. (1999), and could be confirmed here. However, in contrast to Kartner's study, ΔF508 CFTR was not detectable with either the M3A7 or the MATG 1104 monoclonal antibody. Absence of CFTR-reactive signals in sweat glands from ΔF508 homozygous patients was also shown by Kälin et al. (1999). The major difference between our study and that of Kartner et al. (1992) is that we had ascites available rather than purified antibody, as in Kartner's work. We were able to detect ΔF508 CFTR only with the cc24 antibody, whose epitope is located in the regulatory domain of CFTR. This polyclonal antibody was produced in New Zealand White rabbits against a synthetic peptide of the R domain (Picciotto et al. 1992). In our hands, this antibody was the most sensitive in detecting both wild-type and ΔF508 CFTR and produced no unspecific signals on the biological controls, except for the faint background staining that was observed in all controls. It clearly differed from the specific signals in that it was invariably observed irrespective of the patient's genotype. The increase in sensitivity of the antibody is obviously accompanied by a slight decrease in specificity, which can easily be compensated for by the appropriate controls, as demonstrated.
Of eight CFTR antibodies established in the literature, three were suitable for immunodetection of wildtype CFTR, and one of ΔF508 CFTR, in sweat glands. As discussed in Kartner's publication, M3A7 gains sensitivity on antibody purification (Kartner et al. 1992). It is therefore justified to assume that the difference among the three antibodies with respect to detection of ΔF508 CFTR lies in their sensitivity for CFTR rather than in differences between the wild-type and ΔF508 CFTR proteins. All three epitopes are situated outside the ΔF508 deletion, which is located in the first nucleotide binding fold.
Footnotes
Acknowledgements
We thank A. Nairn, J. R. Riordan, and Transgène for providing antibodies, and H. Bolte for technical assistance with cryosections.