
Abstract
Cyclic nucleotides act as second messengers and regulate many functions in various tissues. Many kinds of cyclic nucleotide phosphodiesterases (PDEs) have been reported as regulators of intracellular cAMP and cGMP concentrations. On the basis of amino acid sequence analysis and biochemical properties, ten PDE families have been recognized in mammalian tissues (Beavo 1995; Soderling et al. 1998, 1999; Fujishige et al. 1999; Juilfs et al. 1999).
The cGMP-binding cGMP-specific PDE (PDE5) is highly specific for cGMP and is involved in regulation of the intracellular concentration of cGMP in various tissues. PDE5 was purified from rat and bovine lung (Francis and Corbin 1988; Thomas et al. 1990), and its enzymatic activity is found in the lung, vascular and tracheal smooth muscle cells, spleen, and platelets (Lincoln et al. 1976; Hamet and Coquil 1978; Coquil et al. 1985; Ahn et al. 1989). To obtain information about physiological roles of cGMP in various tissues, many pharmacological experiments using PDE5-specific inhibitors have been done. Zaprinast, a PDE5 inhibitor, increases cGMP concentration in vascular smooth muscle cells (VSMCs), which leads to vascular relaxation (O'Donnell and Owen 1986; Souness et al. 1989; White and Martin 1989). E4021, a potent PDE5-specific inhibitor, is effective in enhancing cGMP accumulation in VSMCs and reduces hypoxic vasoconstriction, pulmonary artery pressure, and total pulmonary resistance with no significant effect on cardiac output, systemic pressure, and resistance (Cohen et al. 1996). Another potent PDE5-specific inhibitor, sildenafil, is effective in treating male erectile dysfunction by increasing cGMP in corpus cavernosa smooth muscle (Boolell et al. 1996). Local administration of zaprinast promotes induction of cerebellar long-term depression in rat cerebellar slices (Hartell 1996). These findings indicate that the cGMP/PDE5 pathway plays an important role in controlling functions in vascular vessels and in the central nervous system.
A cDNA encoding PDE5 was first cloned from bovine lung and was found to encode a protein of 865 amino acid residues (McAllister-Lucas et al. 1993). We have isolated human, canine, and rat PDE5 cDNAs from lung cDNA libraries and here described cDNAs encoding two variants of PDE5 (PDE5A1 and PDE5A2) in these species (Kotera et al. 1997, 1998; Yanaka et al. 1998). Our studies revealed high levels of PDE5 transcripts in rat cerebellum and intestine in addition to tissues containing VSMCs. Low levels of PDE5 transcripts were detected in rat kidney and adrenal gland. In human tissues, PDE5A1 transcripts were particularly abundant in the pancreas (Kotera et al. 1999). In situ hybridization showed that rat PDE5 transcripts were localized to the Purkinje cells in the cerebellum (Kotera et al. 1997). However, we failed to reveal cellular localization of PDE5, except for the brain, by in situ hybridization. Elucidation of the cellular localization of PDE5 may help to explore the possibility of the presence of cGMP signaling in tissues. To study further the role of PDE5 in tissues where PDE5 transcripts were observed in Northern blot analysis, we required an antibody to the PDE5 that would offer high resolution and specificity.
Here we raised an antibody that bound to the PDE5 for use in immunohistochemistry. The antibody recognizes both human and rat PDE5 proteins in immunoblot analysis. Moreover, the antibody described here has been utilized to show the localization of PDE5 in the brain, kidney, and pancreas at the light microscopic level.
Materials and Methods
Production of Full-length Recombinant Human PDE5 Protein
To obtain a human PDE5A cDNA fragment for expression, we performed a polymerase chain reaction (PCR) with primers designed according to the sequence of human PDE5A1 reported previously (Yanaka et al. 1998). To generate a cDNA fragment coding for the N terminus of PDE5A1, PCR was performed using the 5′ primer 5′-GGGAGCTCCATGGAGCGGGCCGGCCCCAGC-3′, the 3′ primer 5′-TTTCAGTAAATGTCCCACCG-3′, and human PDE5A1 cDNA as a template. Denaturing, annealing, and polymerase reaction were done 25 times at 94C for 1 min, at 54C for 1 min, and at 72C for 1 min. An amplified fragment was digested by SacI and EcoRV, subcloned into the corresponding sites of pBluescript II SK(+) (Stratagene; La Jolla, CA), and then sequenced, producing pBlue-PDE5A1-N. To obtain a fragment coding for the C terminus of PDE5A1, PCR was performed using the 5′ primer 5′-CACTGTACATTAAGCGGCGAGG-3′, the 3′ primer 5′-GGGTCGACGGCATATGGCAGAACACACCATCTC-3′, and human PDE5A1 cDNA as a template. An amplified fragment was digested by PstI and SalI, subcloned into the corresponding sites of pBluescript II SK(+ ), and then sequenced, producing pBlue-PDE5A1-C. A 2.0-kb EcoRV-PstI cDNA fragment of human PDE5A cDNA, the 540-bp SacI and EcoRV fragment of pBlue-PDE5A1-N, and the 150-bp PstI-SalI fragment of pBlue-PDE5A1-C were inserted into the SacI and SalI sites of the maltose binding protein (MBP) fusion protein expression vector pMAL-cRI (New England BioLabs; Beverly, MA), resulting in pMAL-PDE5. The MBP fusion protein was produced in E. coli strain JM109 carrying the pMAL-PDE5 plasmid by isopropyl-β-D-thiogalactopyranoside induction, as described previously (Sambrook et al. 1989). The MBP fusion human PDE5 protein (MBP-PDE5) expressed in E. coli was purified by affinity chromatography on an amylose resin column according to the manufacturer's instructions.
Antibody Production
A rabbit polyclonal antiserum was raised against the purified recombinant MBP-PDE5. The MBP-PDE5 protein was mixed with Freund's complete adjuvant (Difco; Detroit, MI) for the first immunization and with Freund's incomplete adjuvant when boosted. After immunizing two Japanese White rabbits (Kitayama Laboratories; Nagano, Japan) four times, antiserum was collected. An anti-PDE5 antibody IgG fraction was prepared from the antiserum by affinity chromatography on a protein G column (Amersham Pharmacia Biotech; Poole, UK). The IgG fraction was eluted with 0.1 M glycine-HCl (pH 2.7), and the eluate was neutralized immediately with 1 M Tris-HCl (pH 9.0), and stored at -20C until use. Protein concentrations of antibodies were determined with a protein assay kit (Bio-Rad; Melville, NY) with bovine serum albumin as the standard.
For some immunohistochemical studies, an anti-PDE5 antibody was purified by affinity chromatography on an MBP-PDE5-conjugated column. The affinity column was prepared using a HiTrap NHS-activated column (Amersham Pharmacia Biotech) according to the manufacturer's instructions. The antibody was eluted with 0.1 M glycine-HCl (pH 2.7) and then neutralized immediately as described above.
Immunoblot Analysis
Cytosolic extracts of COS-7 cells expressing recombinant human PDE5A1 and recombinant rat PDE5A2 proteins were prepared as described previously (Kotera et al. 1997; Yanaka et al. 1998). Male Sprague-Dawley rats at 10 weeks of age (Japan SLC; Shizuoka, Japan) were anesthetized with sodium pentobarbital before various tissues were excised. Rat platelets were prepared by the methods of Baenziger and Majerus (1974). Fresh platelets were sonicated in ice-cold homogenization buffer [20 mM Tris-HCl (pH 7.4), 2 mM magnesium acetate, 0.3 mM CaCl2, 1 mM dithiothreitol, 40 ± leupeptin, 1.3 mM benzamidine, 0.2 mM phenylmethyl-sulfonylfluoride, and 1 mM NaN3] for 1 min and the lysates were stored at -20C until use. Rat lungs and cerebellums were dissected, washed in ice-cold PBS (pH 7.2), and frozen in liquid nitrogen. They were homogenized with a polytron homogenizer in the ice-cold homogenization buffer. The homogenate was centrifuged at 1000 × g for 20 min at 4C to remove nuclei and cell debris. The supernatant was further centrifuged at 100,000 × g for 1 hr at 4C. For SDS-PAGE analysis, the extracts were denatured by heating at 100C for 5 min in sample buffer [2% sodium dodecyl sulfate, 2% 2-mercaptoethanol, 60 mM Tris-HCl (pH 6.8), and 10% (v/v) glycerol]. The proteins were separated by electrophoresis on 7.5% gels and transferred onto Immobilon-P (Millipore; Bedford, MA). The membranes were blocked in Block Ace (Snow Brand Milk Products; Tokyo, Japan) overnight, followed by incubation with anti-PDE5 antibody IgG fraction, preimmune IgG fraction (5 μg/ml each), or anti-PDE5 antibody (500 ng/ml) purified by the MBP-PDE5 affinity column, in PBS containing 0.1% Tween-20 (PBS-T). They were washed and incubated with biotin-conjugated anti-rabbit IgG antibody and avidin-biotin-peroxidase. The bound antibody was detected with enhanced chemiluminescence reagents from Amersham Pharmacia Biotech. The specificity of the immunoblot was determined by incubation with the anti-PDE5 antibody IgG fraction that was mixed at room temperature for 1 hr in the presence and absence of the purified MBP-PDE5.
Immunohistochemistry
Tissues freshly excised from 12-week-old male rats were fixed overnight with 10% formalin in 0.1 M phosphate buffer (pH 7.2). The tissues were dehydrated in graded ethanol solutions and embedded in paraffin. Four-micrometer sections were cut and mounted on Superfrost Plus slides (Fisher Scientific; Springfield, MA). The paraffin was removed with xylene and the tissues were rehydrated through graded ethanol to water. Endogenous peroxidase was blocked by 0.3% hydrogen peroxide in methanol for 20 min at room temperature. Nonspecific staining was blocked by incubation with 1.5% normal goat serum in PBS for 1 hr at room temperature in a humidified atmosphere. The sections were incubated with the anti-PDE5 antibody IgG fraction, the anti-PDE5 antibody purified by the MBP-PDE5 affinity column, or normal IgG fraction prepared from preimmune serum at the same concentration overnight at 4C. After three washes of 10 min each in PBS-T, the sections were incubated with biotinylated goat anti-rabbit immunoglobulin G (Vector Laboratories; Burlingame, CA) for 30 min at room temperature and washed three times, followed by incubation for 30 min at room temperature with the avidin-biotin-peroxidase complex (Vector Laboratories). After three washes, visualization was carried out by incubating the sections with a solution of 50 mM Tris-HCl (pH 7.5), 0.1% 3,3′-diaminobenzidine tetrahydrochloride (Wako Pure Chemicals; Osaka, Japan), and 0.03% hydrogen peroxide. The sections were stained with hematoxylin before they were mounted.
Reverse Transcriptase-PCR and Southern Blot Analysis
Reverse transcriptase (RT)-PCR and Southern blot analysis were performed to detect PDE5A1 and PDE5A2 transcripts in rat tissues. To examine the efficiency of PCR amplification, we first produced reference DNAs. A cDNA fragment coding for N-terminal regions of rat PDE5A1 (amino acid residues 1-79 in addition to the 5′-untranslated region of 21 bp) was amplified using the 5′ primer 5′-AAAACTCGAGCAGAAACCCGCGGCAAACACC-3′, the 3′ primer 5′-GCATGAGGACTTTGAGGCAGAGAGC-3′, and rat PDE5A1 cDNA isolated previously (Kotera et al. 1999) as a template. The rat PDE5A2 cDNA fragment (amino acid residues 1-47 in addition to 5′-untranslated region of 9 bp) was produced using the 5′ primer 5′-ACCTCTGCTATGTTGCCCTTTGG-3′, the 3′ primer 5′-GCATGAGGACTTTGAGGCAGAGAGC-3′, and rat PDE5A2 cDNA isolated previously (Kotera et al. 1999) as a template. PCR was carried out for 25 cycles of denaturation at 94C for 1 min, annealing at 55C for 1 min and extension at 72C for 1 min. Two amplified fragments were subcloned into the TA cloning vector pGEM easy (Promega; Madison, WI) and then sequenced, resulting in reference DNAs, pGEM-5A1N and pGEM-5A2N. The efficiency of PCR amplification was estimated using various concentrations of pGEM-5A1N and pGEM-5A2N as templates under the same conditions as described above. The PCR products were separated by electrophoresis and detected with ethidium bromide staining. The gels were scanned and quantitated by a PDI 420oe Scanner (PDI; Huntington, NY) and Quantity One Program (PDI).
Rat PDE5A1 and PDE5A2 transcripts in tissues were detected by RT-PCR and Southern blot analysis. After RT reaction was carried out by using 1 μg of total RNA prepared from rat cerebellum or 1 μg of poly A+ RNA from rat kidney and pancreas, PCR amplification was performed under the same conditions as described above. The PCR products were subjected to 2% agarose gel electrophoresis and the fractions were transferred onto Hybond-N+ nylon membrane (Amersham Pharmacia Biotech). To confirm that PCR products were derived from PDE5A transcripts, we detected both PCR products by Southern blot analysis using a 32P-labeled DNA probe prepared using a Tth111I-SpeI fragment of rat PDE5A cDNA. After hybridization, the membranes were exposed to X-ray film at RT for 15 min. The resultant films were scanned and quantitated as described above.
Results
Immunoblot Analysis
A polyclonal antibody towards the purified MBP-PDE5 was produced, and anti-PDE5 antibody IgG fraction was purified from the antiserum by affinity chromatography on a protein-G column. The antigenic specificity was investigated with COS-7 cells that express recombinant human and rat PDE5 proteins. Cytosolic extracts of transfected COS-7 cells were electrophoretically separated, blotted onto membranes, and incubated with the anti-PDE5 antibody IgG fraction. The IgG fraction showed immunoreactive bands of ~110 kD and ~97 kD in the cytosolic extracts of transfected COS-7 cells expressing human and rat PDE5 proteins, respectively (Figure 1). No band was detected either with preimmune rabbit IgG fraction or with the mixture of the anti-PDE5 antibody after preincubation with the purified MBP-PDE5 protein used to raise the antibody (Figure 1). These findings showed that the anti-PDE5 antibody bound to both human and rat PDE5 proteins. Immunoblot analysis of lysates from rat platelets, in which PDE5 is abundant, also gave a specific staining of a ~97-kD band (Figure 1). The band of ~97 kD was diminished with a preadsorbed anti-PDE5 antibody IgG fraction. This antibody was therefore considered to be specific to PDE5 and not to react with any other proteins in rat platelets. To reduce nonspecific staining, we purified an anti-PDE5 antibody by an affinity column conjugated with the purified MBP-PDE5. The purified anti-PDE5 antibody detected a major band at ~97 kD in cytosolic extracts from rat lung and cerebellum (Figure 1). Therefore, the antibody was also considered to be specific to PDE5 and not to react with any other proteins in these tissues.
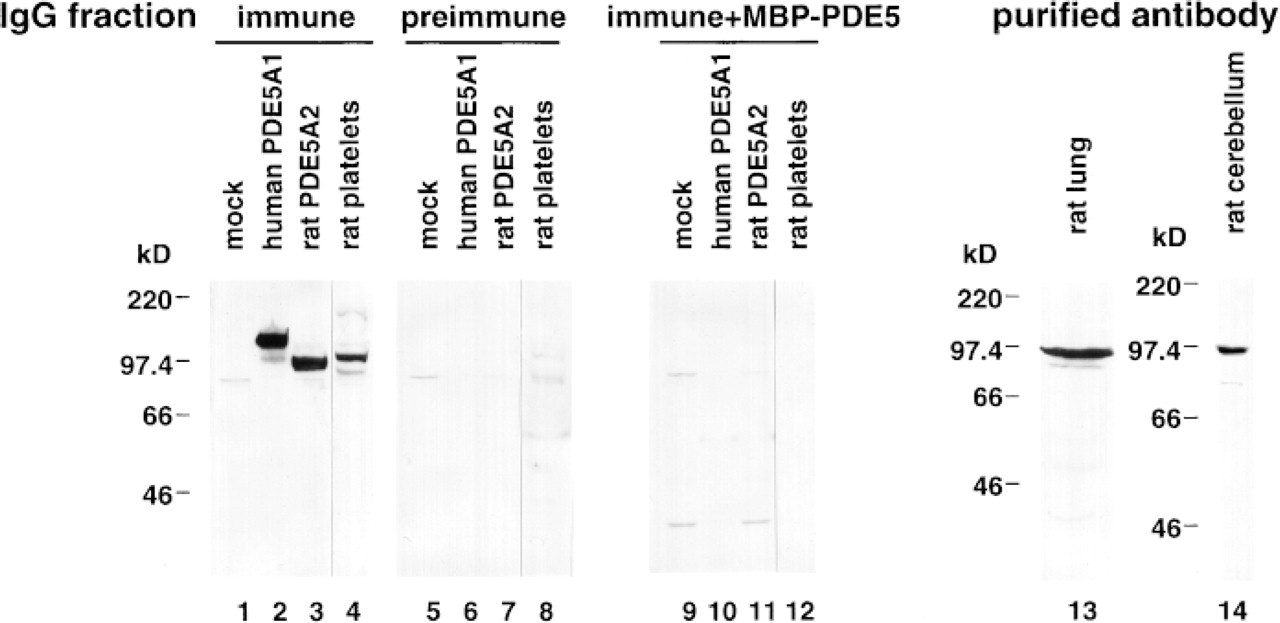
mmunob ots probed with the anti-PDE5 antibody. Cytosolic fractions from transfected COS-7 cells expressing human (Lanes 2, 6, 10) and rat (Lanes 3, 7, 11). PDE5 proteins were analyzed by immunoblotting using the IgG fractions. Mock-transfected cells are shown in Lanes 1, 5, and 9. Protein fractions showing approximately equal PDE5 activities were subjected to 7.5% SDS-PAGE (Lanes 2, 3, 6, 7). The amounts of loaded protein of mock transfected cells were equal to those of extracts expressing rat PDE5A2. Lysates of rat platelets are shown in Lanes 4, 8, and 12. Cytosolic extracts prepared from rat lung (Lane 13) and cerebellum (Lane 14) were examined by immunoblotting using the anti-PDE5 antibody purified by an MBP-PDE5 affinity column. One hundred μg of lysates from rat platelets and 300 μg of extracts from rat lung and cerebellum were loaded on 7.5% SDS-PAGE.
Immunohistochemical Analysis
In rat cerebellum, immunohistochemistry at the light microscopic level using the anti-PDE5 antibody IgG fraction showed intense labeling of most Purkinje cell layers. However, other Purkinje cells, whose nuclei were visualized by hematoxylin, did not appear to be stained by the antibody (Figure 2A). No specific staining was observed when the sections were incubated with preimmune IgG fraction (Figure 2B). The PDE5 protein was expressed in the cell bodies and the dendrites of cerebellar Purkinje cells, and the expression was observed in the first, second, and third branches of the dendrites (Figure 2C). In contrast, no signal was seen in the axons of cerebellar Purkinje cells at any region of the cerebellum. The PDE5 protein was not detected in other parts of brain using the anti-PDE5 antibody IgG fraction (not shown).
To detect PDE5 protein clearly in tissues except for the brain, the anti-PDE5 antibody purified by the MBP-PDE5 affinity column was used for immunostaining. The purified anti-PDE5 antibody demonstrated that PDE5 was localized on the proximal renal tubules and medullary collecting ducts (Figures 3A and 3C). No signal was detected in glomeruli. Preimmune IgG fraction gave no positive signal (Figures 3B and 3D).
In the pancreas, epithelial cells of pancreatic ducts were apparently immunostained with the anti-PDE5 antibody purified by the MBP-PDE5 affinity column (Figures 3E and 3G). The antibody immunostained specifically neither pancreatic islet cells nor pancreatic acinar cells. In our experiment, the PDE5 activity was not observed in RIN-m cells derived from rat pancreatic islets (not shown). This finding is in accordance with the result from the immunohistochemistry using the anti-PDE5 antibody. No signal was detected in the pancreas by using the preimmune IgG fraction (Figure 3F).
RT-PCR and Southern Blot Analysis
The relative amounts of PDE5A1 and PDE5A2 transcripts in rat tissues were examined by RT-PCR. The efficiency of PCR amplification using specific primer sets for PDE5A1 and PDE5A2 was first estimated (Figure 4A), and an optimal linear range for PCR amplification of reference DNAs was obtained up to 3 pg of pGEM-5A1N and 10 pg pGEM-5A2N, which was then used for determination of the relative amounts of PDE5A transcripts in tissues. The amplification using the primer set for PDE5A1 was found to be approximately twice as efficient as that using the primer set for PDE5A2 in the optimal range. By contrast, each PCR amplification using 10 pg and 30 pg of pGEM-5A1N and 30 pg pGEM-5A2N appeared to reach saturation.
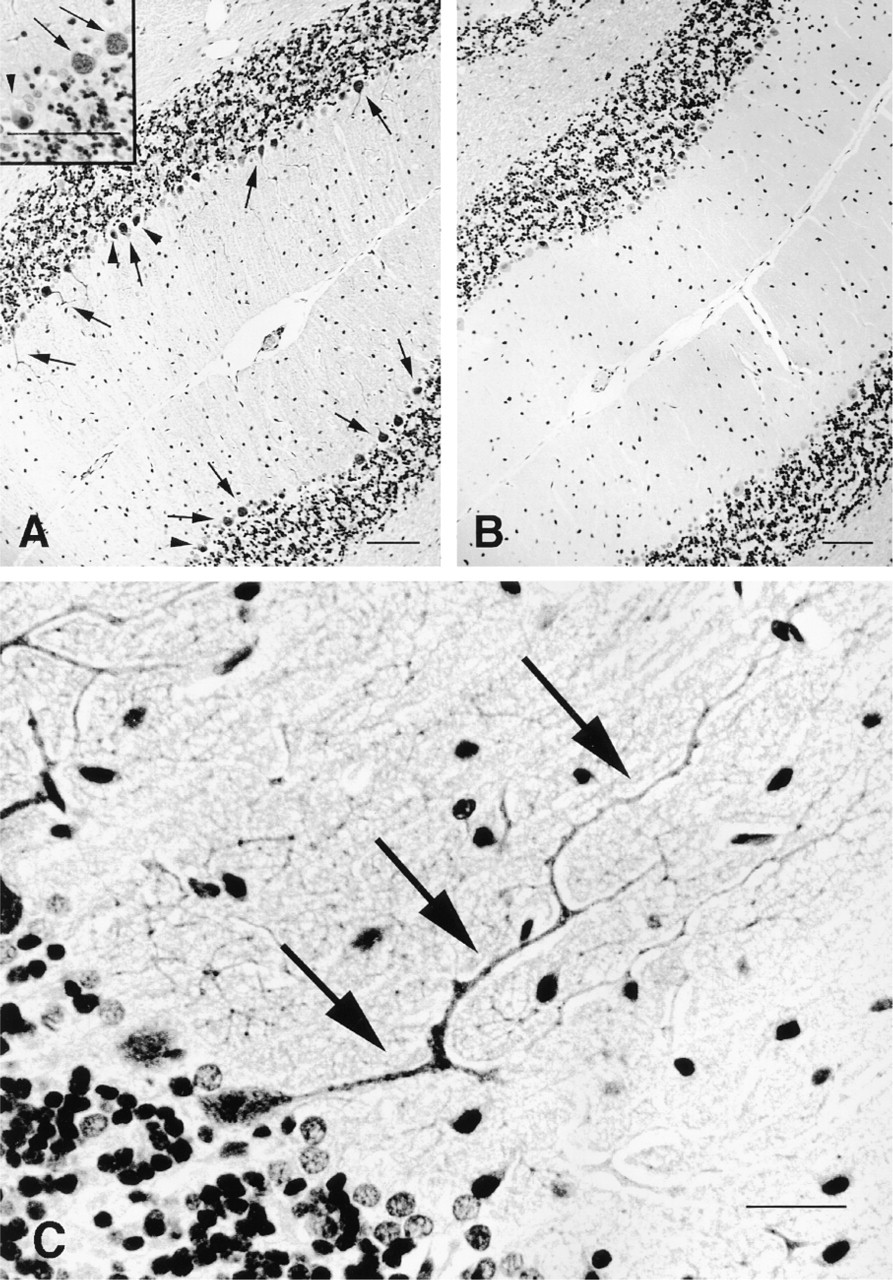
Localization of cells immunoreactive with the anti-PDE5 antibody in rat cerebellum. (
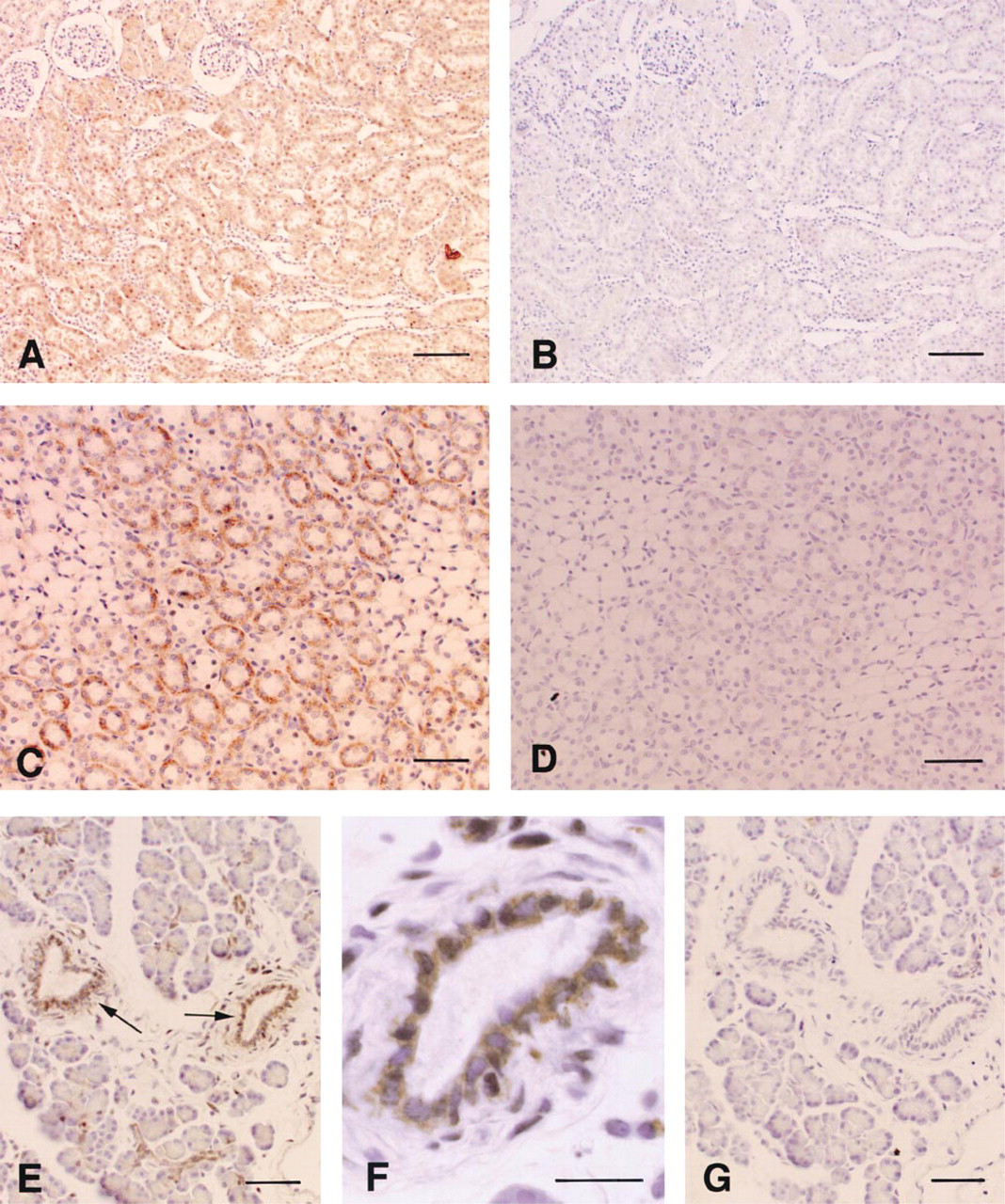
Localization of cells immunoreactive with the anti-PDE5 antibody in rat kidney and pancreas. Immunolocalization of PDE5 in proximal tubules (
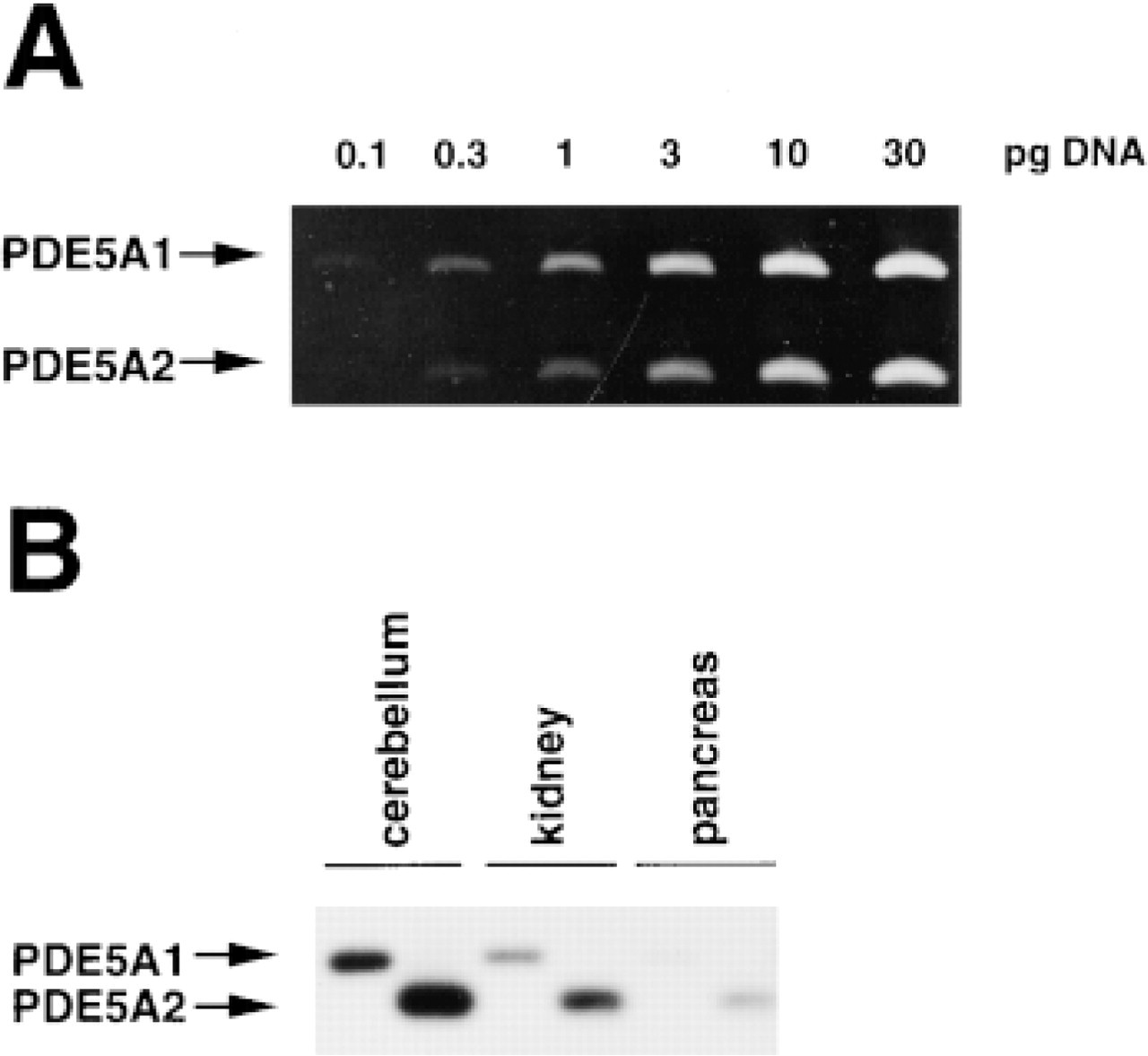
Detection of rat PDE5 transcripts (PDE5A1 and PDE5A2) in tissues by RT-PCR. (
RT-PCR and Southern blot analysis demonstrated the presence of both PDE5A1 and PDE5A2 transcripts in the rat cerebellum, kidney, and pancreas (Figure 4B), in which the PDE5 protein was detected by immunostaining. Semiquantitation of gene expression of two types of PDE5A in rat tissues by RT-PCR fell within the optimal range of reference DNA amplification. The PCR product derived form PDE5A2 transcripts appeared to be two to three times more abundant in the rat cerebellum, kidney, and pancreas compared with that from PDE5A1 transcripts. Considering the efficiency of reference DNA amplification, the amount of PDE5A2 transcripts in these tissues was four to six times greater than that of PDE5A1 transcripts in rat tissues.
Discussion
To precisely determine the cell types producing PDE5 in rat tissues, we have developed a polyclonal antibody against recombinant MBP fusion human PDE5. In immunoblot analysis, the anti-PDE5 antibody immunoreacted with both recombinant human and rat PDE5 proteins. We also detected PDE5 immunostaining by the antibody in rat platelets, lung, and cerebellum. Immunoblot analysis using the anti-PDE5 antibody showed the presence of an immunoreactive band at approximately the expected molecular size of PDE5 (~95 kD), whereas no strong signal was detected at any other molecular size. Because the band disappeared with the preadsorbed anti-PDE5 antibody, the ~97-kD protein stained by the antibody would be expected to be derived from rat PDE5.
The precise localization of PDE5 in the cerebellum was visually revealed with the anti-PDE5 antibody. The cerebellum is well known as a memory center for coordinating and learning movements. Nitric oxide (NO) and cGMP are associated with the induction of long-term depression among some neurons of the cerebellum (Shibuki and Okada 1991). Neuronal NO synthase is found in parallel fibers but not in Purkinje cells and climbing fibers (Rodrigo et al. 1994). In contrast, expression of soluble guanylate cyclase is restricted to Purkinje cells (Ariano et al. 1982). NO released from parallel fibers evokes cGMP generation in Purkinje cells by soluble guanylate cyclase. An increase in cGMP, via activation of parallel fibers and climbing fibers, evokes long-term depression in Purkinje cells. In situ hybridization demonstrated that the cerebellar expression of PDE5 transcripts is limited to Purkinje cells (Kotera et al. 1997). Local administration of zaprinast, a PDE5 inhibitor, leads to an increase of cGMP release from cerebellar slices and promotes induction of cerebellar long-term depression in rat cerebellar slices when paired with parallel fiber stimulation (Hartell 1996). Here we have described that the anti-PDE5 antibody bound specifically to most Purkinje cells in the cerebellum. Interestingly, the PDE5 protein was localized not only in the cell body but also in the dendrites of Purkinje cells in molecular layers, whereas no staining was observed in the axons of Purkinje cells in granule layers at any region of the cerebellum. The anti-PDE5 antibody showed the presence of PDE5 protein at least in the first, second, and third branches of dendrites of Purkinje cells, although in situ hybridization analysis in our previous studies could not reveal the intracellular localization of PDE5 protein. Recently, similar results from another group have been reported in mouse cerebellum (Juilfs et al. 1999). Therefore, PDE5 may be important in degradating cGMP as soon as cGMP generates near each synapse by the stimulation of parallel fibers, and the presence of PDE5 in the dendrites may be attributed to cGMP signaling for the induction of local long-term depression between specific synapses. Moreover, our immunohistochemical analysis using the anti-PDE5 antibody also revealed that some Purkinje cells in the rat cerebellum did not appear to stain, although it is not clear which types of Purkinje cells do not express PDE5. We postulate that this finding indicates that the cGMP/PDE5 pathway may be not associated with functions in some Purkinje cells.
PDE isozymes in suspension of renal tubules and glomeruli have been reported. As for the population of PDE1 and PDE5, a different profile was shown between tubules and glomeruli. In glomeruli, PDE5 is more abundant than PDE1 (Matousovic et al. 1997). In contrast to glomeruli, cGMP is hydrolyzed equally by PDE1 and PDE5 in tubules (Robison et al. 1971). Immunohistochemical analysis demonstrated that, under our experimental conditions, the anti-PDE5 antibody detected PDE5 protein in proximal tubules, whereas PDE5 protein in glomeruli was not stained by the antibody. An increase in cGMP by stimulation of atrial natriuretic peptide (ANP) inhibits the Na+ channel in the luminal membrane of the inner medullary collecting ducts (IMCD) cells (Dousa 1999). The activity of cGMP-hydrolyzing PDEs in homogenates of IMCD in rats with nephrotic syndrome produced either by adriamycin or by immune complex Heymann nephritis is increased (Valentin et al. 1992, 1996). Zaprinast restored cGMP accumulation in response to ANP in IMCD incubated in vitro, and in vivo restored the ability to increase salt and cGMP excretion in response to a volume load in rats with nephrotic syndrome (Valentin et al. 1992, 1996). Here we have detected PDE5 protein in medullary collecting ducts by the anti-PDE5 antibody. These findings suggest the presence of the cGMP/PDE5 pathway in renal functions such as sodium excretion. It will be interesting to reveal the precise expression pattern and regulation of PDE5 in renal dysfunction by using the anti-PDE5 antibody.
Epithelial cells of pancreatic ducts secrete bicarbonate into pancreatic ducts in response to cAMP elevation by the stimulation of secretin (Schulz 1980). The bicarbonate concentration in pancreatic juice is increased by Ro20,1724, a potent PDE4 inhibitor, and by 3-isobutyl-1-methylxanthine, a nonselective PDE inhibitor (Iwatsuki et al. 1994), via the elevation of the intracellular concentration of cAMP. With regard to cGMP function, KRN2391, a synthesized vasodilator, and nitroprusside, which produces NO, increase the secretion of pancreatic juice with a high concentration of protein and a low concentration of bicarbonate with the increase of the cGMP levels in pancreatic tissues in anesthetized dogs (Iwatsuki et al. 1993). The K+ channel opening action or increasing of blood flow rate by NO does not participate in the secretion. In these studies, it is not clear whether cGMP is directly related to the secretion of bicarbonate. Here, we first demonstrated that the expression of PDE5 was limited to epithelial cells of pancreatic ducts, suggesting that the cGMP/PDE5 pathway exists in these cells. This result raises the possibility that NO produced by nitroprusside has a direct effect on cell functions via cGMP elevation in epithelial cells of pancreatic ducts.
Previously, we reported that many regulatory elements, including cAMP response elements, were present in the 5′-untranslated region and intron of the human and rat PDE5A genes (Kotera et al. 1999). The level of PDE5 transcripts, especially the PDE5A2 transcripts, is increased by the stimulation of a cAMP analogue in cultured rat VSMCs, suggesting that the PDE5A2 is an inducible variant of PDE5A. Here we have demonstrated that PDE5A2 transcripts were more abundant in the cerebellum, kidney, and pancreas of rats than PDE5A1 transcripts. Intracellular cAMP produced by certain stimulants may regulate PDE5A2 expression in these tissues. The amounts of PDE5 transcripts increase during perinatal pulmonary development, reaching their maximal levels in rat neonates at 8 days after birth (Sanchez et al. 1998). In the rat cerebellum, PDE5 transcripts increase markedly between 4 days and 28 days of age, and none were observed in newborn rats (Kotera et al. 1997). Therefore, PDE5 expression is also regulated during peri- and postnatal development. It will be intriguing to examine the changes of PDE5 expression in various tissues in development and certain diseases at the microscopic level. The anti-PDE5 antibody that we have produced here clearly has potential for precise cellular localization of PDE5 protein in certain tissues and might provide the most relevant information on the role of the cGMP/PDE5 pathway in pharmacological studies using specific PDE5 inhibitors.