
Abstract
Squamous cell carcinoma antigen (SCCA) serves as a serological marker for advanced squamous cell carcinomas (SCCs) and as an indicator of therapeutic response. Recent molecular studies show that the SCCA is transcribed by two almost identical tandemly arrayed genes, SCCA1 and SCCA2. These genes are members of the high molecular weight
Keywords
I
Several studies show that increased serum SCCA levels correlate with the extent of disease in patients with cervical SCC (Kato and Torigoe 1977; Senekjian et al. 1987; Duk et al. 1990; Bolli et al. 1994). For example, the incidence of elevated pretreatment serum SCCA levels range from 33% in Stage IB to 93% in Stage IV disease (Kato 1992). Higher SCCA levels are also indicative of deep tumor infiltration and lymph node involvement (Duk et al. 1990; Takeshima et al. 1998). Moreover, the measurement of posttreatment SCCA levels is useful in monitoring the response to therapy and in predicting tumor recurrence and metastasis. Duk et al. (1990) showed that 79% of patients who failed treatment had persistent elevations in serum SCCA levels, whereas 91% of those without clinical evidence of tumor had undetectable SCCA levels. Elevated serum SCCA levels are also found in patients with advanced SCCs of the head and neck, lung, and esophagus. In head and neck SCCs, posttreatment SCCA values are the most important factor in predicting disease-free survival as well as overall survival (Lara and Cuyas 1995).
In addition to squamous cell tumors, SCCA is also detected in the sera of patients with skin disorders, such as psoriasis and eczema (Duk et al. 1989; Campbell and De'Ambrosis 1990), and with lung diseases, such as bronchitis and pneumonia (Molina et al. 1990; Kato 1992). These findings raise the possibility that SCCA may serve as a marker for certain inflammatory conditions as well as SCCs.
Molecular cloning of the cDNA shows that SCCA belongs to the superfamily of high molecular weight
Although SCCA1 and SCCA2 are almost identical members of the serpin superfamily, significant differences in their reactive site loops (i.e., that part of the molecule that is bound by the active site of the proteinase) suggest that they inhibit different classes of proteinases. Biochemical analysis of recombinant proteins shows that SCCA1 is a cross-class inhibitor of papain-like cysteine proteinases, such as cathepsins L, S, and K, whereas SCCA2 inhibits chymotrypsin-like serine proteinases, cathepsin G, and mast cell chymase (Schick et al. 1997,1998). These findings may have important diagnostic and therapeutic implications, because the balance between proteinases and inhibitors can affect tumor cell motility, invasiveness, proliferation, and death.
Previous descriptions of the expression pattern of the SCCA isoforms in normal and malignant tissues were deduced from studies using nondiscriminatory monoclonal reagents or an antibody specific for the acidic isoform (SCCA2) only (Kato et al. 1987). However, the specificity of these reagents has not been confirmed using recombinant SCCA1 and SCCA2 proteins. The recent identification of distinct inhibitory functions for SCCA1 (neutral SCCA) and SCCA2 (acidic SCCA) underscores the importance of analyzing the tissue distribution of these serpins with reagents and methods that can discriminate between these two highly homologous proteins. The purpose of this study was to use a new set of discriminatory monoclonal antibodies (MAbs) and polymerase chain reaction (PCR) to define the distribution of SCCA1 and SCCA2 in normal adult tissues and SCCs of the head, neck, and lung.
Materials and Methods
Selection of Primers and PCR Conditions
PCR assays were performed in 10-μl volumes containing 0.1-0.2 ng first-strand cDNA, 200 μM of each dNTP, 0.5 μM of each primer, 1.5 mM MgCl2, 10 mM Tris-HCl, 50 mM KCl and 0.5 U of Thermus aquaticus (Taq) DNA polymerase. The reaction mixtures were cycled 35 times at 94C for 30 sec, 58C for 30 sec, and 72C for 30 sec. Genomic DNAfree first-strand cDNA was purchased from Clontech Laboratories (Palo Alto, CA). Negative controls (no cDNA) and positive controls [SCCA1 and SCCA2 cDNA in pBluescript plasmid vectors (Stratagene; La Jolla, CA)] were included for each primer set. Discriminatory sense (S) primers for SCCA1 (1-8S, 5'-CGCGGTCTCGTGCTATCTGG-3' (MA39) and SCCA2 (2-8S, 5'-CACGGTCTCTCAGTATCTAA-3') were from exon 8 and correspond to nucleotides 1022-1041 of the published cDNAs of SCCA1 and SCCA2 [GenBank accession numbers U19558 (SCCA1) and U19569 (SCCA2) (Schneider et al. 1995)]. A common antisense (AS) primer (8AS-1,5'-AGAAGAGGATGCTGTTGGTC-3') corresponds to nucleotides 1211-1192. These primers amplify a 191-bp product from SCCA1 and SCCA2. To ensure that positive signals were not due to contaminating genomic DNA, reactions were repeated using a different set of discriminatory sense primers from exon 6 for SCCA1 (1-6S, 5'-CCTGAAGGTAATATTGGCAGC-3') and SCCA2 (2-6S, 5'-CTGATGGGACTATTGGCAATG-3') and a different nondiscriminatory antisense primer from exon 8 (8AS-2,5'-GGGATGAGAATCTGCCATAG-3'). These primers amplify a 681-bp fragment from SCCA1 and SCCA2. To control for cDNA integrity, human β-actin primers were used to amplify a 616-bp fragment. The S (5'-CCTCGCCTTTGCCGATCC-3') and AS (5'-CCTAGAAGTACTCCATCAGTCAG-3') primers for β-actin amplification correspond to nucleotides 277-294 from exon 1 and nucleotides 2242-2220 from exon 4 from the published cDNA (GenBank accession number M10277) (Nakajima-Iijima et al. 1985).
Generation of MAbs Specific for SCCA1 and SCCA2
BALB/c mice (Charles River Laboratories; Stockbridge, MA) were injected with whole yeast extract containing full-length recombinant SCCA1 or SCCA2. Antigens emulsified in RIBI adjuvant were injected IP into mice on Days 1 and 21. Responders received an intrasplenic immunization on Day 35. Spleen cells were harvested on Day 38 and fused to P3×63Ag8U.1 mouse myeloma cells (American Type Culture Collection; Rockville, MD) in the presence of polyethylene glycol 1500 (Boehringher Mannheim; Indianapolis, IN) Hybridomas were selected for growth in Dulbecco's modified Eagle's medium (DMEM, 4.5% glucose) (BioWhittaker; Walkersville, MD) supplemented with 100 μg/ml endothelial cell mitogen (Biomedical Technologies; Stoughton, MA), 10 mM sodium hypoxanthine, 40 μM aminopteridin, 1.6 mM thymidine, 20% fetal bovine serum, 1 mM nonessential amino acids solution, 2 mM L-glutamine, 1 mM sodium pyruvate solution, 50 U/ml penicillin, and 50 μg/ml streptomycin (Life Technologies; Grand Island, NY). Hybridomas were screened by an antibody capture ELISA using purified recombinant SCCA1 or SCCA2 as the antigen. Positive clones were subcloned by two or three rounds of limiting dilution (one cell/well).
Specificity of SCCA1 and SCCA2 MAbs by Immunoblotting
Immunoblots containing recombinant proteins GST-SCCA1, GST-SCCA2, and a related serpin, chicken ovalbumin, were prepared to test the specificity of the MAbs (Figure 1). Proteins were separated by SDS-PAGE and electroblotted at 100 V for 1 hr at 4C onto reinforced nitrocellulose (Nitroplus; Micron Separations, Westborough, MA). The transfer buffer was 25 mM Tris base, pH 8.0, 190 mM glycine (pH 8.3). After three rinses in washing buffer (PBS, pH 7.4, 0.1% Tween-20), the blot was incubated for 1 hr in I-Block (Tropix; Bedford, MA) blocking buffer (PBS, pH 7.4, 0.1% Tween-20, 0.2% I-Block) and then incubated for 1 hr with purified mouse monoclonal ascites as the primary antibody. The primary antibodies were used at the following dilutions: anti-SCCA1 (clone 8H11) 1:2000 (1.7 μg/ml); anti-SCCA2 (clone 10C12) 1:100,000 (0.03 μg/ml). The blot was washed three times in wash buffer and then incubated for 1 hr with alkaline phosphatase-conjugated goat anti-mouse IgG (Jackson Immunoresearch Laboratories; Westgrove, PA) as the secondary antibody. After three rinses in wash buffer and two rinses in chemiluminescent assay buffer (0.1 M diethanolamine-HCl, pH 9.8, 1.0 mM MgCl2, 5% Nitroblock), the blot was incubated for 5 min with chemiluminescent substrate (CDP-Star; Tropix). Chemiluminescence was detected by autoradiography.
Cell Transfections and Immunocytochemistry
The MDA-MB-435 breast carcinoma cell line (kindly provided by Dr. Ruth Sager), which expresses no SCCA by immunoblotting or RT-PCR (unpublished data) was transfected stably with plasmids containing recombinant SCCA1 or SCCA2 cDNA under control of the EF-1 promoter (Mizushima and Nagata 1990). Mock transfectants were generated by transfection with the plasmid alone. Transfected cells were selected by culturing cells in DMEM supplemented with 10% fetal bovine serum, 1 mM nonessential amino acids solution, 2 mM L-glutamine, 1 mM sodium pyruvate solution, 50 U/ml penicillin, 50 μg/ml streptomycin, and 500 μg/ml hygromycin B. After 3–4 weeks of selection, single colonies were isolated by cloning cylinders and expanded for immunostaining.

Specificity of SCCA1 and SCCA2 MAbs by immunoblotting. rSCCAl (Lane 2), rSCCA2 (Lane 3) and a related serpin, chicken ovalbumin (Lane 1) were separated by SDS-PAGE and transferred to nitrocellulose membranes. Two identical membranes were incubated with purified mouse monoclonal ascites 8H11 (left) or 10C12 (right). Protein-bound antibodies were detected by alkaline phosphatase-conjugated goat anti-mouse IgG and chemiluminescent substrate.
For immunostaining, SCCA1, SCCA2, and mock-transfected MDA-MB-435 cells were grown to 50% confluency on 4-chambered slides (Nalge Nunc International; Naperville, IL). The cells were washed in PBS (0.01 M phosphate buffer, 27 mM KCl, 137 mM NaCl, pH 7.4), fixed by incubating in 10% neutral-buffered formalin for 15 min, and permeabilized by incubating in 0.2% Triton X-100 in PBS for 4 min. The immunocytochemistry was performed as described below, except that the peroxidase blocking step was omitted, undiluted hybridoma supernatants served as the primary antibody, and hematoxylin was used as the counter-stain.
Immunohistochemistry
Most of the tissues used for immunohistochemical analysis were obtained from surgical resection and diagnostic biopsy material from Beth Israel Deaconess Medical Center. All heart specimens (3/3), and some pancreas (1/4), liver (1/4), spleen (1/4), and adrenal (1/3) tissues were obtained from human adult autopsies performed within 10 hr of expiration. The normal tissue survey included specimens from three or more patients (except for tongue and urethra; see Table 1) and was confined to non-neuronal tissues. Three or more specimens of lung (squamous cell, adeno-, large-cell, and small-cell) and head and neck (tongue) SCCs of various histologic types and differentiation states were also examined (Table 2). All tissues were fixed in 10% neutral-buffered formalin and embedded in paraffin according to conventional procedures. Four-μm-thick tissue sections placed on polysine-coated slides (Fisher; Atlanta, GA) were deparaffinized in xylenes (two 5-min washes) and rehydrated in graded ethanol solutions (100%, two 5-min washes; 95% and 70%, two 1-min washes) and distilled water (one 5-min wash). Between the absolute and 95% ethanol washes, samples were incubated for 10 min in a 0.3% H2O2 (v/v) methanol solution to quench endogenous peroxidase activity. Antigen retrieval methods employing enzymatic digestion (0.05-0.25% trypsin; 0.1% pepsin at 37C) and microwave/steamer heating (10 mM citrate buffer, pH 6.0) were tested to optimize SCCA1 and SCCA2 immunostaining (Mepham et al. 1979; Shi et al. 1993). A 10-min incubation with 1 × trypsin-EDTA (0.05% trypsin, 0.53 mM EDTA.4Na; Gibco BRL, Grand Island, NY) at 37C enhanced SCCA2 staining, whereas neither procedure improved SCCA1 staining.
SCCA1 and SCCA2 immunoreactivity in normal human tissues

aNumber of cases examined.
SCCA1 and SCCA2 immunoreactivity in head, neck, and lung squamous cell carcinomas
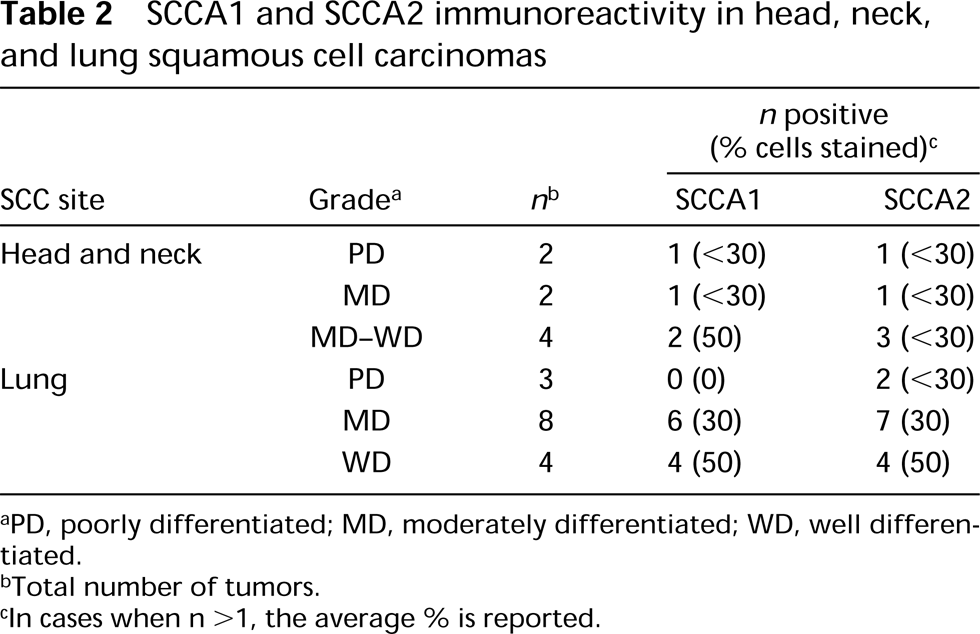
aPD, poorly differentiated; MD, moderately differentiated; WD, well differentiated.
bTotal number of tumors.
cIn cases when n >1, the average % is reported.
Immunostaining was performed at room temperature in a humidified chamber. The standard incubation volume for each reagent was 200 μl. Sections were circled with a PAP pen (Zymed Laboratories; San Francisco, CA) and blocked by incubating with 10% normal goat serum diluted in PBS with 1% (w/v) bovine serum albumin (Sigma Chemical; St Louis, MO) for 30 min. After washing in PBS, the sections were incubated with the primary antibody (purified mouse monoclonal ascites) for 1 hr at the following dilutions: anti-SCCA1 (clone 8H11) at 1:1000 (final concentration 3.4 μg/ ml); anti-SCCA2 (clone 10C12) at 1:500 (6.0 μg/ml). Optimal hair follicle staining in skin required a 1:500 dilution of anti-SCCA1 MAb (6.8 μg/ml). For certain normal tissues and for all neoplastic tissues, it was necessary to block endogenous biotin activity, especially in antigen-retrieved sections (e.g., salivary gland ducts, bronchial mucous glands, liver, kidney, testis, and endocrine tissues). A biotin blocking kit (SP-2001: Vector Laboratories; Burlingame, CA) was used as follows: 4 drops of Avidin D solution (70 μg/ml) was added per ml normal goat serum before incubation on sections. Sections were washed briefly in PBS, then incubated with 4 drops of biotin solution (20 μg/ml) per ml diluted primary antibody. For all tissues tested, the primary antibody was replaced with mouse monoclonal anti-glutathione-S-transferase IgG (Sigma) diluted 1:1000 (10 μg/ml) as a negative control. As a positive control, each tissue type was stained with a rabbit anti-SCCA heteroantiserum that does not discriminate between SCCA1 and SCCA2 (Schick et al. 1997).
After the primary antibody incubation, sections were washed for 10 min in PBS, then incubated with a biotinylated goat anti-mouse IgG (Jackson Immunoresearch Laboratories) diluted 1:1000-1:2000 (1.0 −0.5 μg/ml) as the secondary antibody. After a 10-min PBS wash, the secondary antibodies were detected by incubation for 30 min with an avidin-biotin-peroxidase complex (Vectastain Elite ABC horseradish peroxidase kit; Vector Laboratories), followed by a 10-min PBS wash, then a 5-min incubation with a 3,3'-diaminobenzidine substrate/chromogen solution (Liquid DAB-Plus substrate kit; Zymed Laboratories). Sections were washed in several changes of distilled water and counterstained with methyl green.
To control for fixation artifacts, immunostaining was also performed on fresh frozen vagina (positive control), skin, breast, fetal membranes, and testis. These tissues were embedded in Tissue-Tek OCT Compound (Miles; Elkhart, IN), sectioned to 4 μm using a cryostat and fixed by dipping in 25C acetone. Slides were stored at −80C until use. Before staining, sections were equilibrated to −20C in a cryostat, fixed in acetone for 10 min at −20C, washed with cold (4C) PBS, and blocked for endogenous peroxidase as above. Immunohistochemistry was performed as above with 1:100 dilutions of anti-SCCA MAb 8H11 (34 μg/ml) and anti-SCCA2 MAb 10C12 (30 μg/ml).
Immunostaining results were assessed in a blinded fashion by two independent observers. In normal tissues, immunoreactivity was reported as present or absent with qualitative comments about the intensity of staining, such as weak, moderate, or strong. In addition, immunostaining in tumor tissues was assessed as the percentage of positively stained cells.
Results
Survey of SCCA1 and SCCA2 Expression by PCR
First-strand cDNA libraries were assayed to determine whether SCCA1 and SCCA2 were expressed in different tissues (Figures 2A and 2B). SCCA1- and SCCA2-specific primers amplified the correct-sized fragments from lung, lymph node, thymus, tonsil, placenta, prostate, and testis cDNAs. Neither SCCA1 nor SCCA2 was detected in samples from brain, heart, kidney, liver, pancreas, skeletal muscle, bone marrow, fetal liver, peripheral blood leukocytes, spleen, colon, ovary, and small intestine. All PCR assays were performed at least three times and the positive reactions for SCCA1 and SCCA2 were repeated by using the primer set from exons 6 and 8 (not shown). This result confirmed that the positive signals were not derived from contaminating genomic DNA sequences. The control β-actin fragment was amplified from all of the cDNA samples tested (Figure 2C).
Specificity of SCCA1 and SCCA2 MAbs Using Immunocytochemistry on Transfected Cell Lines
To confirm the specificity of these antibodies as an immunohistochemical reagent, we studied their reactivity against transfected MDA-MB-435 cell lines expressing either SCCA1 or SCCA2. A mock-transfected (non-SCCA-expressing) MDA-MB-435 cell line served as a negative control. Anti-SCCA1 MAb 8H11 showed immunoreactivity only with SCCA1-transfected MDA-MB-435 cells and not with the SCCA2 or mock-transfected MDA-MB-435 cells (Figures 3B, 3E, and 3H). Similarly, anti-SCCA2 MAb 10C12 reacted against only the SCCA2-transfected MDA-MB-435 cells (Figures 3C, 3F, and 3I). These studies confirmed that the MAbs 8H11 and 10C12 could discriminate between SCCA1 and SCCA2 when used as an immunohistochemical reagent.
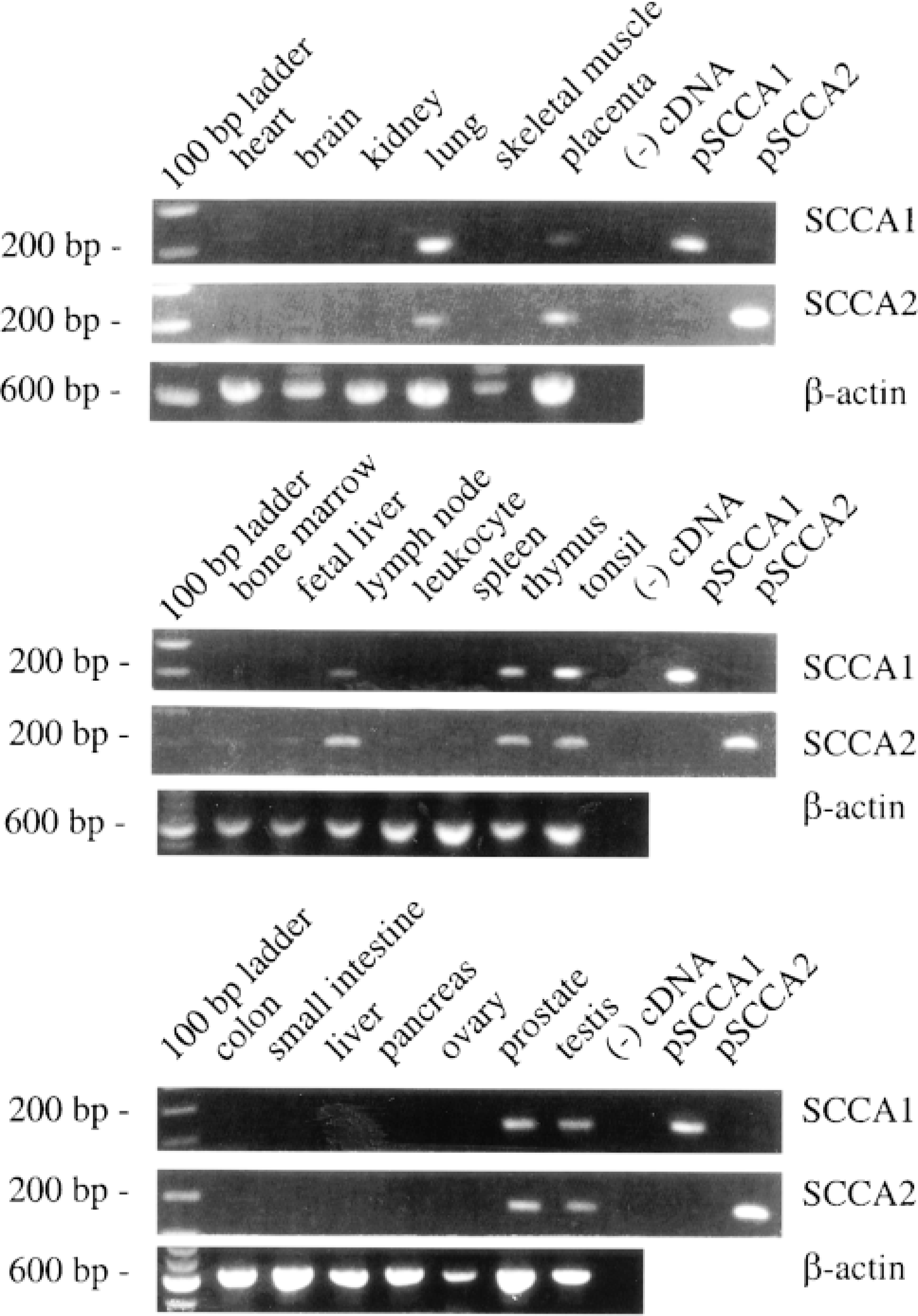
RNA expression of SCCA1 and SCCA2. cDNAs generated by reverse transcription of polyA+-selected RNA from various human tissues served as templates in PCR assays with SCCA1, SCCA2, and β-actin primers. SCCA1 and SCCA2 primers amplified a 191-bp fragment; β-actin primers amplified a 616-bp fragment. Tissue sources are labeled above each lane of the ethidium bromide-stained 1% agarose gels. No cDNA served as a negative control. Plasmids containing SCCA1 or SCCA2 cDNA served as positive controls for SCCA1 and SCCA2 primers.
Immunohistochemical Analysis of Human Tissues Using SCCA1- and SCCA2-specific MAbs
The results are summarized in Tables 1 and 2 and representative sections are illustrated in Figures 3J–3U and 4.
Adipose Tissue. No SCCA1 or SCCA2 immunoreactivity was observed in adipocytes.
Musculoskeletal System. No SCCA1 or SCCA2 immunoreactivity was observed in skeletal muscle, myofibrils, chondrocytes, or osteocytes.
Skin. In normal adult skin, weak to moderate focal SCCA1 and SCCA2 cytoplasmic immunoreactivity was detected in the external root sheath of selected hair follicles. In some cases, the epidermis adjacent to positively stained hair follicles exhibited weak to moderate suprabasal staining for SCCA1 and SCCA2. However, normal epidermis away from these structures was negative for SCCA1 and SCCA2 immunoreactivity (Figures 3K and 3L). Moderate to strong cytosolic staining was noted in the suprabasal epidermal keratinocytes (strata granulosum and spinosum) over areas of inflammation. Often these areas were marked by epidermal thickening (acanthosis).
Cardiovascular System. No SCCA1 or SCCA2 immunoreactivity was detected in cardiomyocytes, fibroblasts, endothelial cells, or arterial smooth muscle cells.
Respiratory System. The pseudostratified columnar epithelial cells lining the nasopharynx, trachea, bronchi, and bronchioles exhibited strong primarily cytosolic immunoreactivity for SCCA1. SCCA2 staining was moderate to strong (Figures 3N and 3O). In the alveoli, both Type I and Type II pneumocytes were nonreactive.
Digestive System. Strong cytosolic SCCA1 immunoreactivity was observed in the suprabasal squamous epithelium of the tongue, and SCCA2 staining was moderate to strong. Weak to moderate cytosolic immunoreactivity for both SCCA1 and SCCA2 was detected in the suprabasal squamous epithelium lining the tonsils. Parotid and minor salivary glands were negative for SCCA1 and SCCA2, and submandibular gland ducts showed weak to moderate immunoreactivity only in areas of squamous metaplasia (not shown). Strong cytosolic immunoreactivity was detected for SCCA1 in the suprabasal keratinocytes of the stratified squamous epithelia in both esophageal and anal mucosa. SCCA2 staining was moderate to strong in these structures. No staining was detected in any cell type of the stomach, small or large intestine, pancreas, liver, or gallbladder.
Hematolymphoid System. Moderate to strong staining was observed for both SCCA1 and SCCA2 in the thymic epithelial cells in Hassall's corpuscles (Figures 3Q and 3R). Hematopoietic cells of the bone marrow, medullary and cortical thymocytes, and lymphoid cells of lymph nodes, tonsils, and spleen were all nonreactive.
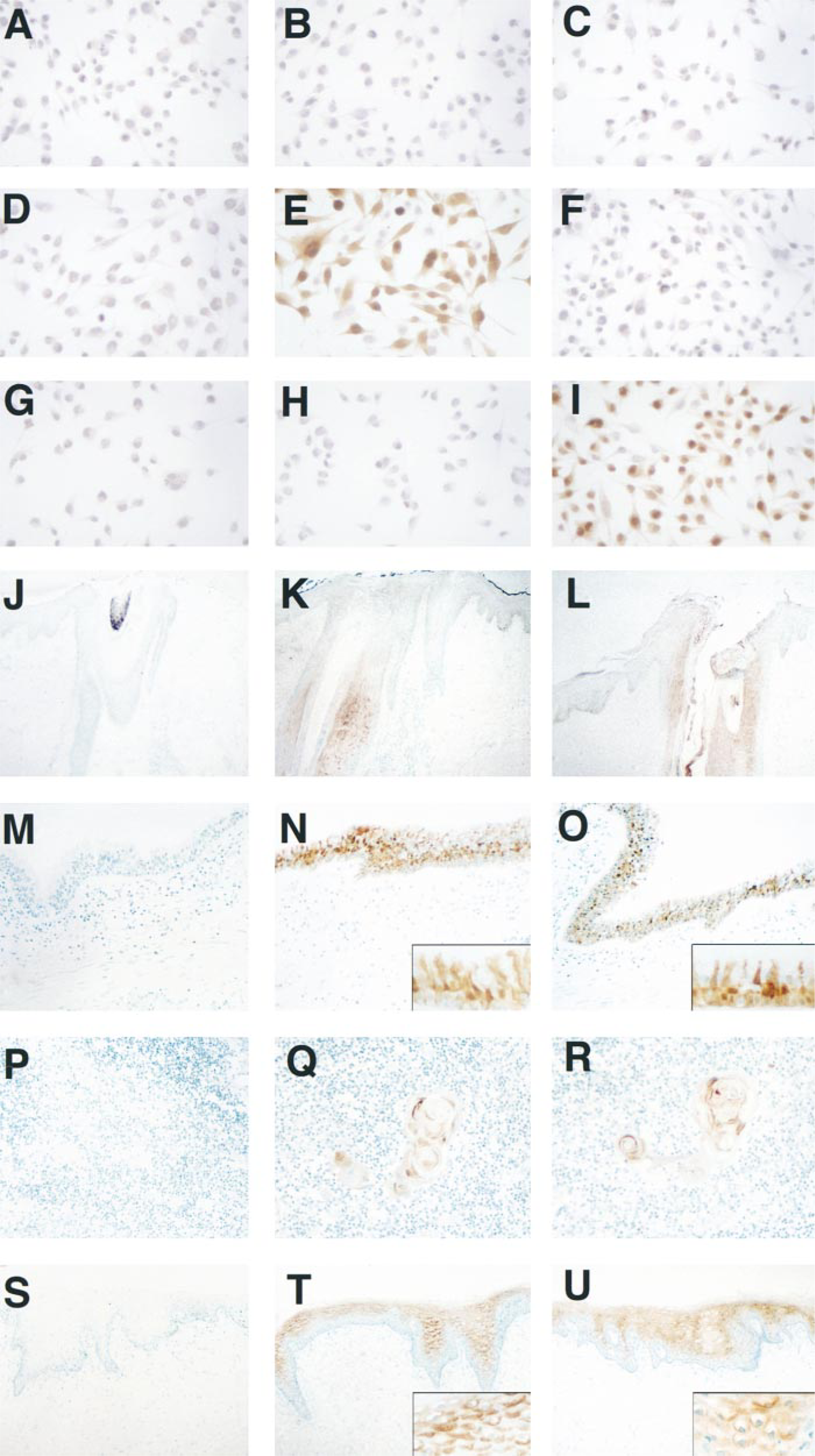
Specificity of SCCA1 and SCCA2 MAbs by immunocytochemistry (top) and normal adult human tissue distribution of SCCA1 and SCCA2 by immunohistochemistry (bottom). (
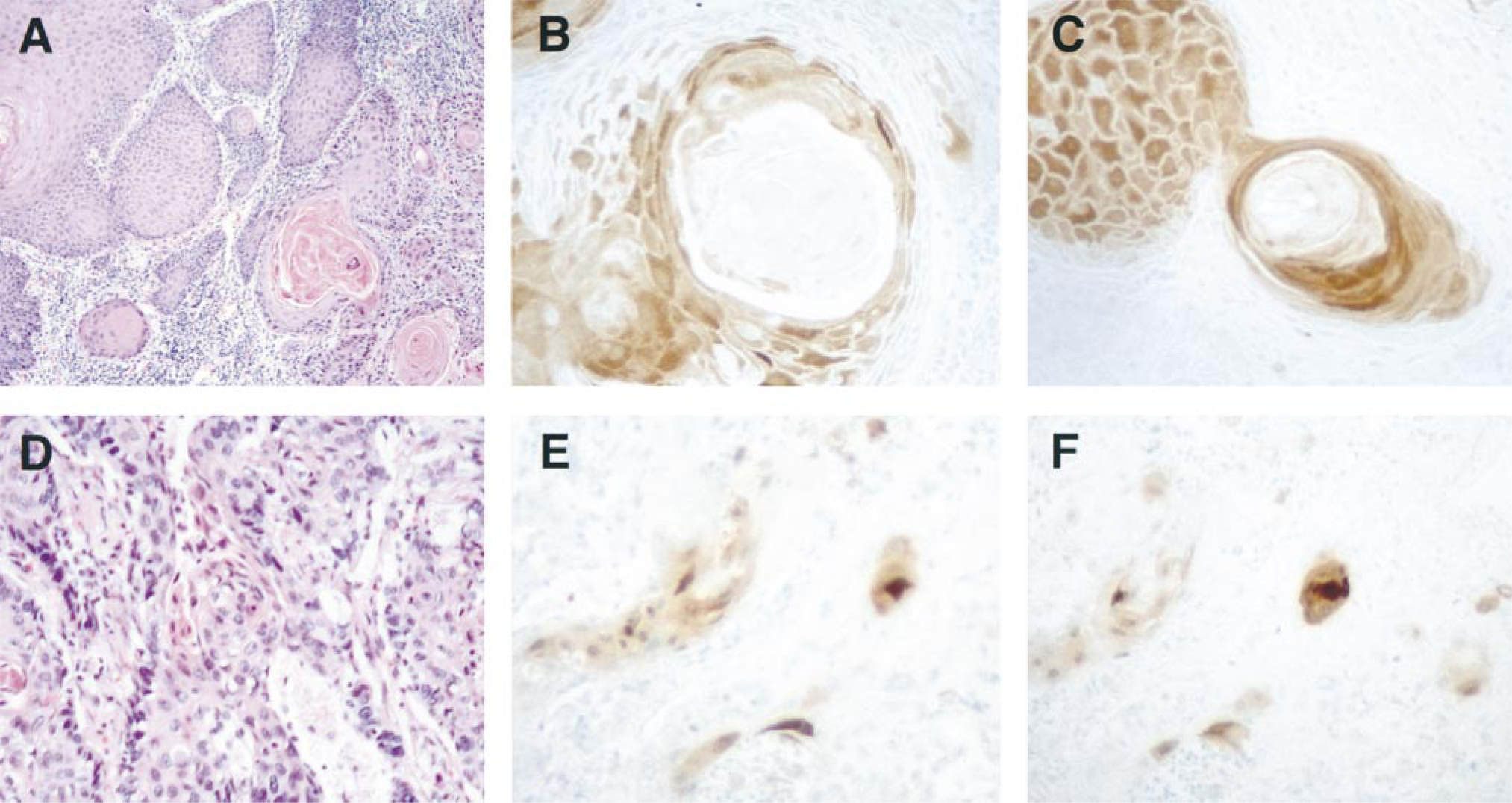
Immunohistochemistry of head, neck, and lung SCCs with SCCA1 and SCCA2 MAbs. Columns represent sections stained with (left) hematoxylin and eosin, (center) SCCA1 MAb, and (right) SCCA2 MAb. Panels depict representative moderately to well-differentiated tongue SCC (
Urinary System. No SCCA1 or SCCA2 immunoreactivity was detected in kidney, ureter, urethra, or bladder.
Male Reproductive System. No SCCA1 or SCCA2 immunoreactivity was observed in normal prostate ducts or acinar tissue. However, selected areas of prostate showed expression of SCCA1 and SCCA2. These areas were focal and were marked by atrophy and/or chronic inflammation as well as squamous metaplasia (not shown). No SCCA1 or SCCA2 immunoreactivity was observed in testis, epididymis, vas deferens, seminal vesicles, or rete testis.
Female Reproductive System/Mammary Gland. Strong cytosolic SCCA1 immunoreactivity was detected in the intermediate, suprabasal stratified squamous epithelia of the uterine cervix and vagina. SCCA2 staining was moderate to strong in the former and strong in the latter (Figures 3T and 3U). All cells in the uterus, Fallopian tubes, ovaries, and pre-/postmenopausal mammary gland were nonreactive for SCCA1 and SCCA2.
Female Reproductive System/Placenta. No SCCA1 or SCCA2 immunoreactivity was observed in any cell type within the fetal membranes, placental disk, or umbilical cord.
Endocrine System. In addition to negative results observed in the reproductive endocrine tissues, neither antigen was detected in thyroid, parathyroid, adrenal gland, or pancreatic islets.
Head and Neck SCCs. Weak to moderate cytosolic SCCA1 immunoreactivity was observed in four of the seven primary tongue SCC cases (Table 2). In general, SCCA1 immunoreactivity was observed in the more keratinized and better-differentiated cells within the tumors and was confined to less than 50% of the population (Figure 4B). Similarly, cytosolic SCCA2 immunoreactivity was detected in five of the seven primary tongue SCCs, and the intensity was moderate to strong in well-differentiated tumors compared to the less intense staining of poorly differentiated tumors (Figure 4C). In almost all cases, the antigens co-localized to selected keratinizing tumor nests, with no appreciable difference in intensity of the staining between SCCA1 and SCCA2. In contrast to the homogeneous staining pattern in suprabasal layers of normal stratified squamous epithelia, SCCA1 and SCCA2 immunoreactivity in SCCs was heterogeneous and sparse, with some predilection for the superficial portion of the tumors.
Lung Carcinomas. Of the four major forms of lung carcinoma (squamous cell, adeno-, large-cell, and small-cell), SCCA1 and SCCA2 immunoreactivity was detected only in SCCs (Table 2). In the moderately and well-differentiated tumors, the intensity and extent of SCCA1 and SCCA2 staining were comparable and were correlated with their state of differentiation of the SCCs. Focal moderate to intense cytoplasmic staining was observed in selected nonbasal keratinizing cells (Figures 4E and 4F). In two of the three poorly differentiated lung SCCs, weak SCCA2 immunoreactivity was observed in a small percentage of cells. None of the poorly differentiated cases showed SCCA1 staining.
Discussion
In this study we showed that SCCA1 and SCCA2 co-localized to several normal adult human tissues at both the mRNA and the protein level. This co-localization pattern was also observed in most cases of head and neck SCCs and in lung SCCs.
The RNA transcripts of both SCCA1 and SCCA2 genes were detected by RT-PCR (reverse transcription-PCR) in human lung, tonsil, thymus, lymph node, placenta, prostate, and testis. Using discriminatory MAbs whose specificities were confirmed by immunocytochemistry on transfected cell lines, SCCA1 and SCCA2 proteins co-localized to the skin, esophagus, tonsil, tongue, thymus, trachea, bronchi, bronchiole, vagina, and uterine cervix. Overall, SCCA1 and SCCA2 showed a restricted expression pattern in normal tissues, implying tissue-specific control of coordinated gene expression for these serpins. In four tissues (placenta, lymph node, prostate, and testis) in which SCCA1 and SCCA2 transcripts were detected by RT-PCR, immunohistochemistry failed to localize the corresponding proteins. This discrepancy probably reflects the greater sensitivity of the former technique. Consistent with the RT-PCR data, dbEST also contains SCCA cDNA sequences that were cloned from normal placenta and testis libraries.
In normal adult skin, we detected weak and focal SCCA1 and SCCA2 immunoreactivity only in the outer root sheath of selected vellus and terminal hair follicles and, in some of these cases, in the epidermis immediately adjacent to these structures. SCCA immunoreactivity in normal squamous epithelium of the skin was reported in several studies (Mino-Miyagawa et al. 1990; Horiuchi et al. 1994). By radioimmunoassay, Crombach et al. (1989) and Mino-Miyagawa et al. (1990) showed that SCCA concentration in normal skin epithelium is comparable to that in the squamous epithelia of vagina, cervix, esophagus, and SCCs of the cervix. In contrast to these reports, Morioka (1980) and Duk et al. (1989) found either low or absent SCCA expression in the skin. The lack of consensus in the literature regarding SCCA expression in the normal skin is related to at least two factors: antibody specificity and the tissue source. First, the polyclonal antibodies against SCCA are likely to react with the 14 different subfractions of the complex and to give a different staining pattern than that provided by a monospecific reagent. Second, the variability of the SCCA expression in the skin could be related to the characteristics of the “normal” skin tissue used in immunohistochemistry. During our survey, we noted strong SCCA1 and SCCA2 immunoreactivity in the suprabasal layers of the normal epidermis overlying inflamed dermis or in areas adjacent to tumor tissue. These observations suggested that SCCA1 and SCCA2 expression in the skin was inducible and that previous determinations of the baseline expression may have been influenced by local inflammatory or infiltrative processes. Indeed, elevated levels of serum SCCA and increased SCCA content of the skin are associated with inflammatory skin disorders such as psoriasis and atopic dermatitis (Duk et al. 1989; Campbell and De'Ambrosis 1990).
In a previous study, TA-4 immunoreactivity was detected in the intermediate layer of the squamous epithelium in the tongue, pharynx, and esophagus and in the ductal epithelia of the submandibular glands (Mino-Miyagawa et al. 1990). Kato (1996) reports that the acidic form of SCCA is normally found in the saliva as a result of active secretion from the salivary glands. We showed that both SCCA1 and SCCA2 were present in the stratified squamous epithelium of the digestive system lining the mucosal surfaces of the tongue, tonsils, and esophagus, but we did not detect SCCA1 or SCCA2 immunoreactivity in submandibular, parotid, or minor salivary glands. Our results suggested that SCCA1 and SCCA2 in the saliva was derived from the squamous epithelial cells lining mucosal surfaces of the upper digestive tract rather than from the salivary glands.
Interestingly, we noted that both SCCA1 and SCCA2 were present in areas of squamous metaplasia in the ductal epithelium of the submandibular glands and the prostate gland. Other investigators report the presence of TA-4 in areas of squamous metaplasia within the lung (Mino-Miyagawa et al. 1990). Therefore, it appears that SCCA1 and SCCA2 may be co-expressed in non-squamous epithelia undergoing squamous metaplastic changes in response to inflammation or infection.
SCCA1 and SCCA2 expression was not restricted to squamous epithelial cells. SCCA1 and SCCA2 were detected in the pseudo-stratified columnar epithelium of the conducting airways. Although previous reports indicated the presence of SCCA in respiratory secretions, its source was not identified (Kato 1992; Ashitani et al. 1996). Diseases such as asthma, cystic fibrosis, and emphysema are characterized by an influx of inflammatory cells and the release of proteinases that degrade the structural components of the lung. The degree of the damage caused by proteinases in the lung, to a major extent, is determined by the degree of protection afforded by local (e.g., secretory leukocyte proteinase inhibitor and elafin) and systemic proteinase inhibitors (e.g., α1-proteinase inhibitor). Because SCCA1 and SCCA2 are inhibitors of serine and cysteine proteinases, their location in the bronchial mucosa would place them in an ideal position to protect the airways from proteinases derived from inflammatory cells, epithelial cells, and microorganisms.
Another novel finding of this survey was the detection of both SCCA1 and SCCA2 in Hassall's corpuscles of the thymus. Hassall's corpuscles are composed of terminally differentiated keratinized epithelial cells and resemble the suprabasal squamous epithelium in their cytokeratin contents (Shezen et al. 1995). On the other hand, the subcapsular-perivascular epithelium of the thymus, which gives rise to the epithelial cells of the Hassall's corpuscles, resembles the basal layer of squamous epithelium. During their migration to the medulla and ultimately to the Hassall's corpuscles, the cells that originate from the subcapsular-perivascular epithelium gradually lose the cytokeratin markers of basal cells and acquire those of mature keratinocytes. The finding of SCCA1 and SCCA2 expression by the keratinized epithelial cells of the Hassall's corpuscles, but not by their precursor cells, suggests that these serpins are related to epithelial cell differentiation in the thymus gland.
In the female genitourinary system, we detected comparable SCCA1 and SCCA2 staining in the suprabasal layers of the stratified squamous epithelium of the uterine cervix and vagina. Kato et al. (1987) reported that an MAb that binds both the acidic and neutral isoforms of SCCA stained the intermediate layers of the normal squamous epithelium of the cervix. In contrast, a different MAb that detected only the acidic fractions of SCCA failed to stain the normal squamous epithelium. The discrepancy between the results of Kato et al. (1987) and those of this report may be due to the lack of use of antigen retrieval methods in the former study as well as differences in the epitopes recognized by the MAbs.
In head, neck, and lung SCCs, we detected SCCA1 and SCCA2 immunoreactivity in well-differentiated squamous cell components of the tumor tissues. This pattern of expression was similar to that observed in several studies using nondiscriminatory antibodies raised against the TA-4 complex (Tsuji et al. 1989; Kearsley et al. 1990; Mino-Miyagawa et al. 1990). Collectively, these data suggest that SCCA1 and SCCA2 MAbs are useful in assessing the differentiation states of SCCs. In contrast to a previous study (Kato et al. 1987), however, we are unable to confirm a unique distribution pattern of SCCA1 and SCCA2 in SCCs. In a previous study, the neutral isoform (SCCA1) was found throughout the tumor, whereas the acidic fraction (SCCA2) was found only at the invading edge. Once again, this discrepancy may be due to differences between immunohistochemical techniques and antibody specificities.
The co-expression of SCCA1 and SCCA2 in the squamous epithelium of mucous membranes, some areas of the skin, lung, thymus, and in moderately and well-differentiated SCCs suggests an important role for these serpins in the coordinate regulation of certain serine and cysteine proteinases associated with both normal and transformed cells. Further analysis of SCC with the discriminatory MAbs described in this report should determine whether perturbations of the coordinate expression of SCCA1 and SCCA2 alter the proteinase-inhibitor balance and contribute to the transformed phenotype associated with different types of squamous cell carcinomas.
Footnotes
Acknowledgements
Supported by grants from the Smokeless Tobacco Research Council, Inc. (GAS), the Nell and Nancy Fund, a Cancer Prevention Fund supported by AFLAC, Inc. (MPU), the Pine Mountain Benevolent Association (MPU), the Beth Israel Pathology Foundation, Inc., Boston, MA (MPU), and the National Institutes of Health grants HD28475 (GAS), CA69331 (GAS), and CA73031 (CS).
We wish to thank Drs Theodore Kwan, Larry Brown, Stephen Pak, and Cliff Luke for helpful discussions, and Ellie Manseau, Doina Mandrila, and John Prioleau for technical advice.