
Abstract
Threatening stimuli capture our attention more rapidly than benign stimuli, and threatening experiences lead to longer lasting and more vivid episodic memories. The common interpretation of these findings is that humans share an evolved fear response that enables prioritized processing of threats, providing a survival advantage. This response is assumed to be universal; however, these findings have been documented almost entirely in WEIRD (western, educated, industrialized, rich, and democratic) populations. Here, we address this gap by examining threat detection and fear memories in a remote African culture, the Himba. We found that threats captured attention more rapidly than benign stimuli, and that fear memories, despite differing in content, were shaped by threat in a similar manner to that reported in WEIRD populations.
Fear, thought to be a universal response to threat, shapes how we perceive and remember the world. Threatening stimuli capture attention more rapidly than benign stimuli (Blanchette, 2006; Brown et al., 2010; Fox, 1996; Fox et al., 2000, 2007; LoBue, 2014; Mogg & Bradley, 1999; Öhman & Mineka, 2003; Öhman et al., 2001a, 2001b), and encounters with threatening stimuli form more vivid and long-lasting memories (Bowen et al., 2018; Clewett et al., 2020; Holland & Kensinger, 2010; Kensinger, 2009; Sheldon & Donahue, 2017). This impact of fear on attention and memory is thought to confer a survival advantage—rapid detection of threat enables rapid responding, and enhanced memories can provide useful knowledge for the next threatening encounter. The adaptive benefit of these responses supports the notion that they are universal or shared cross-culturally. However, this has been relatively unexplored. The classical findings that fear shapes core aspects of cognition, such as attention and memory, come almost entirely from WEIRD (Western, educated, industrialized, rich, and democratic, Henrich et al., 2010) populations. Here, we begin to address this gap by asking how threat shapes attention and memory in a remote African culture, the Himba. The Himba live in a context radically different from the majority of WEIRD participants, offering a unique opportunity to examine whether fear, and its cognitive consequences still hold despite these differences. We focused on rapid attentional capture and episodic memory because these are two of the most commonly studied areas with respect to fear, and both are widely assumed to be the result of evolutionary-beneficial adaptations. To begin, we review key findings from the WEIRD literature, on the impact of fear on visual attention and on memory, and then briefly introduce how the Himba and their environment, motivating our current study.
The Threat Superiority Effect
The impact of threat on attention is captured by the threat superiority effect (TSE), the well-replicated finding that images of threatening stimuli are detected more rapidly than benign stimuli in a visual search task. Specifically, when participants are presented with an array of images and have to find the odd one out (e.g., 8 ropes and 1 snake) they are faster to detect the target when it is threatening than when it is neutral (e.g., 8 ropes and 1 spaghetti). The TSE has been predominantly accounted for by the classical or evolutionary view—that a specialized fear circuit, or neural module evolved for the rapid detection of threats present at the time of mammalian evolution, such as snakes and spiders (Öhman & Mineka, 2001, 2003). In support of this view, infants orient their attention to images of snakes and spiders more rapidly than to images of other threat types (LoBue & Deloache, 2010). Interestingly, the TSE is also present for modern threats, such as guns, knives, and syringes (Blanchette, 2006; Brosch & Sharma, 2005; Brown et al., 2010; Cinq-Mars et al., 2022; Subra et al., 2017). Besides the few studies with infants, there is not much work testing the universality assumption underlying the classical view of the TSE. Here, we use a straightforward approach to testing this assumption, we ask whether the effect replicates for the commonly tested stimuli (snakes, spiders, guns, and knives) in a non-WEIRD population, the Himba. In addition to rapid attention to relevant threats, we also explore fear memories. While rapid attention is crucial for responding in the moment, forming detailed long-lasting memories of those encounters is also crucial to adaptively respond in the future
Enhanced Vividness and Episodic Richness of Threat Memories
Studies with Western populations show that the memory system treats threatening stimuli and encounters differently from non-threatening events. For example, both humans and non-human primates create more rapid and long-lasting memories for snakes compared to non-threatening animals (Mineka et al., 1980; Öhman et al., 1976). Emotionally charged events that involve real or perceived threat of many kinds are longer-lasting, recalled more frequently (see Dolan, 2002; Hamann, 2001, for reviews), and are more vividly recalled than more mundane events (Bahrick et al., 1998; Brown & Kulik, 1977; Budson et al., 2004; Paradis et al., 2004; Peterson & Bell, 1996; Peterson & Whalen, 2001; Pezdek, 2003; Wolters & Goudsmit, 2005). It is not only the subjective sense of vividness that is impacted, but fear-related memories are actually more detailed. For example, individuals remember more sensory details, such as the shape of the gun, or the color of gloves, from threatening encounters than neutral encounters, although this tends to be for central elements of the threatening stimulus, as opposed to the broader context (e.g., Easterbrook, 1959; Hurlemann et al., 2005; Kensinger & Corkin, 2003; Laney et al., 2004; Mather, 2007). Thus, threatening encounters appear to enhance subjective vividness, and some aspects of the memory, such as sensory details for central details.
Both vividness and sensory details are associated with episodic memory, the ability to recall an episode from one’s past that is distinct in place and time and dependent on the hippocampal memory system (Danker et al., 2017; Staresina et al., 2012; Tompary et al., 2016). One approach to quantifying how episodic a narrative memory is, is the autobiographical interview method (AIM). In this method, participants are cued to describe an event memory from their personal life in as much detail as possible. The memory is then transcribed and the different types of details in the memory are coded into “internal” (i.e., episodic), which includes event details, perceptual (multisensory) details, descriptions of emotions and thoughts at the time of the event, and details about place and time, and “external” details, which includes non-episodic aspects of the narrative, such as general semantic knowledge or reference to other events (Levine et al., 2002; for a methodological review, see Wardell, Esposito, et al., 2021).
Using this method, McKinnon et al. (2015) found that individuals’ memories from a near-death experience on transatlantic flight AT236 were richer in internal detail than more neutral memories. The same pattern was reported in adults who were cued to recall their most frightening memory (Hilverman et al., 2016), and for negative emotional memories (Wardell, Madan, et al., 2021). These studies suggest, in line with lab-based studies, that intense real-life threat encounters enhance hippocampal-dependent episodic memory.
Gesture rate is another marker of episodic memory reinstatement. Specifically, Hilverman et al. (2016) showed that the amount of gestures an individual made when verbally recalling a memory was correlated with the number of internal details recalled, in healthy older adults but not in adults with hippocampal damage. Taken together, these results suggest that threat enhances episodic memory, and that these enhancements in episodic memory can be studied in naturalistic narratives by quantifying memory details and gesture rate.
Like the TSE, these effects are presumed to be universal, but little is actually known about the nature of autobiographical fear memories cross-culturally. Interestingly, there are cross-cultural differences in the types of details that compose neutral episodic memories. East Asian participants’ memories are less rich with episodic detail and are more generalized or schematic than Westerns (Millar et al., 2013). Despite these baseline differences in memory specificity, there is also some evidence that certain experiences shape autobiographical memory similarly across cultures. For example, although Asians had less baseline memory specificity, exposure to traumatic events similarly decreased memory specificity across Western and Asian participants (Humphries & Jobson, 2012). Like traumatic exposure, fear might shape memories in a similar manner, even if the types of threatening encounters differ substantially.
The Himba
The Himba are a semi-nomadic group of approximately 50,000 living in the Kunene region in Northern Namibia. They are pastoralists (tending to herds of goats and cattle) and relocate within the year based on water access. They live in remote villages and have limited formal schooling. Psychologists have studied the Himba for 20 years, but there have been no studies on threat processing in the Himba. One study on emotional perception included fearful faces. Specifically, United States and Himba participants were asked to freely sort images of posed facial expressions of the six “universal” emotion categories. U.S. participants grouped smiling (happy), scowling (angry), wide-eyed (fearful), and neutral faces into distinct piles, while the Himba only distinctly grouped wide-eyed (fearful) and smiling (happy) faces. This was interpreted as evidence against the universality of emotion-recognition in facial expressions (Gendron et al., 2014a). However, it is interesting that of the six emotional categories tested, expressions of happiness and fear were the two categories sorted distinctly. A later study showed the same pattern with auditory emotional utterances (Gendron et al., 2014b). This hints at some universality in the category but doesn’t explore the impact of fear on cognition.
The Current Study
The goal of this study was to examine how fear shapes attention and memory in a remote population, the Himba. To examine attention, we asked whether the classic TSE effect established in WEIRD populations is also present in the Himba. We explored this using both evolutionary-relevant and modern threats and examined whether it was enhanced for a particularly relevant threat, snakes. To gain more knowledge about the threat landscape and examine structure of fear memories, we asked participants to recall their most fearful memory. We utilized coder ratings of episodic, perceptual, and emotion/thought richness, as well as categorization of internal and external memory details, where internal details are associated with episodic memory (Wardell, Madan, et al., 2021) While there is currently no meta-analysis on relative proportion of internal and external details for different valence memories, previous studies have reported a higher proportion of internal details for memories of a threatening or fearful experiences relative to more neutral memories (Hilverman et al., 2016; McKinnon et al., 2015). The number of internal details has also been linked to gesture rate during recall in the West (Hilverman et al., 2016). Thus, if threat shapes memory in a similar manner in the Himba, we expected memories to be rated as vivid, to be composed primarily of internal details, and for these metrics to positively relate to gesture rate.
Methods
Participants
Participants were 74 (38 female and 36 male) Himba individuals, aged 15-70 (M = 33, median = 30). The TSE is a robust effect, our lab and others have found the effect with an N of 20 to 30 participants (Blumenthal et al., in preparation; Brown et al., 2010; Cinq-Mars et al., 2022; Flykt, 2005; Fox et al., 2007; Öhman et al., 2001a; Subra et al., 2017). Given that these studies have been conducted in highly controlled laboratory settings in the West, we aimed for a larger N with the Himba. Seventy-four participants was the largest N we could achieve across a 3-week research trip. Testing took place in a remote village, Ogongo, as well as in Opuwo, the main regional town (population = 20,000). Most participants (57) were living in and tested in Ogongo, but 18 individuals in Opuwo participated. To receive local permission in Ogongo, the experimenters and guides first introduced themselves to the chief and explained the research project, and then and received permission to set up a testing site. Participants often came by of their own accord from seeing the site, or via word of mouth. The procedure was identical in Opuwo, except that there was no local authority. All participants were native speakers of Otjihimba, and when they came by the site introductions and the general procedure and compensation was explained orally in Otjihimba by the local guides, and oral consent was obtained. Compensation consisted of sugar and soap, which was chosen based on what participants reported liking most during past fieldwork. Following oral consent, the participant was then invited into the testing tent and sat next to the experimenter and translator. The specific experimental procedure and task instructions were provided orally oral consent for the specific task was obtained again. Experiment 1 was always done first, it included recording a threat-evoked heart rate, followed by the visual search task. Experiment 2, the fear-memory task, was done second based on time available and a separate oral consent. Specifically, after completing Experiment 1, participants were given the option to participate in Experiment 2. The translator explained the procedure and asked if they agreed to be filmed while describing their memory. Oral consent was used given low levels of reading ability in the Himba. The local ethics committee of the Université du Québec à Trois-Rivières approved the study.
Experiment 1: Attention to Threat
Threat-Evoked Heart Rate
To establish whether the threat stimuli evoked the common autonomic arousal response in the Himba, we collected heart rate data. Specifically, prior to completing the visual search task, we measured participants heart rate while they passively viewed the matrices from the visual search task, featuring threatening or neutral targets (see Figure 1). Heart rate was recorded using the Cardio app on the experimenters iPhone. Participants placed their pointer finger over the camera lens, the app works by detecting the color change resulting from changes in blood flow at the fingertip with each heartbeat. We extracted beats per minute (BPM) to examine whether BPM was elevated for threatening relative to neutral stimuli. Contact apps that use a phones’ built-in camera to detect blood flow changes have been compared to the gold standard (ECG) and shown to be accurate and reliable measures of BPM in healthy participants (Coppetti et al., 2018; Mitchell et al., 2016). This, combined with the block design which enabled us to have a large number of trials and examine average differences across blocks, gives us confidence that this tool provided a valid measure of stimuli differences.
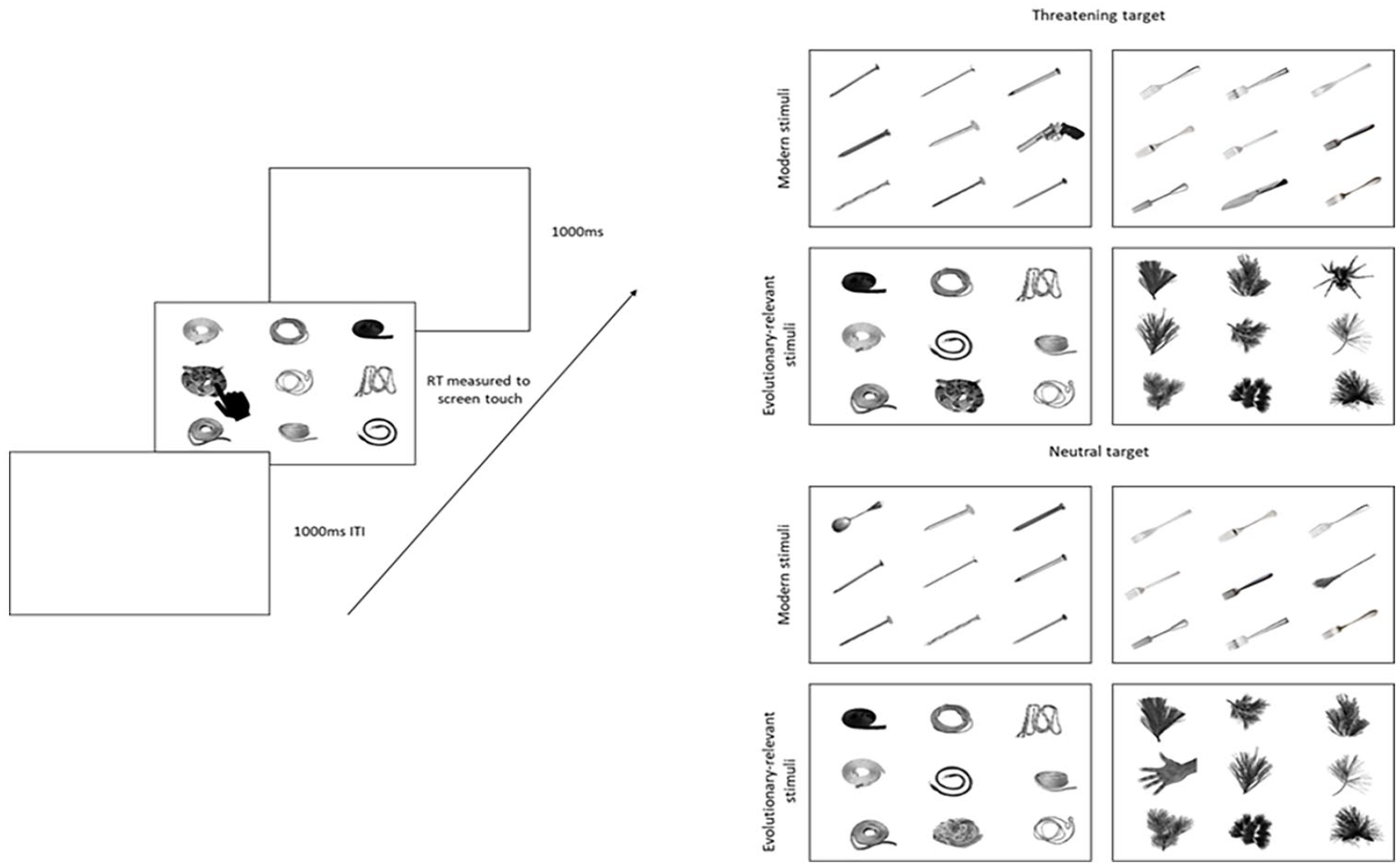
Procedure and Stimuli for the Visual Search Task.
Threat Detection Visual Search Task
Participants completed a visual search task on a touch screen (Figure 1). Before starting the task, they familiarized themselves with the stimuli, touch screen, and reaction time procedure by completing 12 trials in which a single image was presented that they had to reach out and touch as quickly as possible. Specifically, they were instructed to place their hands in a marked spot on the table and reach out to touch the image with their pointer finger as quickly as possible, and then return their hands to the same position before the start of the next trial. Images consisted of an example image from each threat target, neutral target, and distractor category presented at random in one any of the 9 positions on an equally spaced 3X3 grid. Following the familiarization, the experimental task trials began. Participants completed a set of 8 practice trials, 1 for each target type, to ensure they were comfortable with the task before beginning. The procedure was identical, except that the single target image was presented with 8 distractor images from the same category. Participants were encouraged to respond as quickly as possible and place their hands back in the marked position following the trial to ensure reliability in reaction time measures across trials. Targets were either threatening or neutral. Threatening targets included evolutionary-relevant threats (snakes and spiders) and modern threats (guns and knives). Neutral targets matched in shape to the threats included ropes, hands, spoons, and brooms (respectively). On threat target trials, a single target image (snake) was surrounded by 8 distractor images matched in shape (spaghetti). On neutral target trials, a single neutral target (rope) was surrounded by the same 8 distractors (spaghetti). Distractor categories were spaghetti, pines, nails, and forks (Figure 1) and were the same for the threat and neutral targets. This allowed us to examine the reaction time differences between threatening and neutral targets matched in shape (snake and rope) surrounded by the same distractor images (spaghetti). Trials terminated when participants touched the target image. The task consisted of 36 trials, with the 9 items (1 target and 8 distractor images) presented in randomly selected positions. Items were randomly selected from a pool of 9 items. Trial type (threatening or neutral) was also randomized. The dependent measure was reaction time.
Experiment 2: Memory of a Threatening Experience
After completing the visual search task, participants who participated in the second experiment were asked to share their most fearful memory. Specifically, the translator asked them, if they were comfortable, to describe the most fearful experience in their life. Videos of the participant describing their memory, as well as the translator translating were recorded on the experimenters iPhone. There were 30 participants (13 male and 17 female) aged 15 to 60 (M = 29, standard deviation [SD] = 9.4). Almost all participants described their fearful memory in Otjihimba, and the translator translated their description to English out loud to the experimenter. A few participants from Opuwo described their experiences in a mix of English with Otjihimba, which was then translated on the fly. There were 10 individuals from Opuwo (3 male, 7 female), and 20 (14 female, 6 male) from the remote villages Ogongo with no difference in age (Opuwo M = 30.1, Ogongo M = 27, t (29) =0.84, p = .41).
Two approaches were used for data analysis: coder ratings of memory characteristics, and categorization of memory details. Both approaches were adopted from Levine et al. (2002). For coder ratings, research assistants blind to the study hypothesis listened to the translations and provided 3 subjective ratings: episodic richness, perceptual richness, and emotional/thought richness (Levine et al., 2002; Wardell, Esposito, et al., 2021). Episodic richness was defined as the extent to which a subject evokes the impression of true re-experiencing by taking a listener back to a specific moment in time and place in which they are able to recreate the perceptual, emotional and cognitive contextual detail of an event in the past.
Perceptual richness was defined as “the vividness or intensity of the percept” which could be evoked by auditory, olfactory, tactile, taste, visual, or spatial-temporal details. Emotion/thought richness is defined as “the extent to which a person is able to re-create what they were thinking and/or feeling at the time the event occurred.”
For the categorization of memory details, a Namibian colleague who is a native Otjihimba speaker translated and transcribed participants’ memories word for word into English. A research assistant trained in the Levine et al. method then coded memory transcripts into internal (episodic details) and external categories. Internal categories included event details, place details, time details, perceptual details, and emotion/thought details, while external categories included general semantic details, personal semantic details, self-knowledge, external specific events, repeated events, extended events, repetitions, and other. A higher proportion of internal details is associated with specific episodic memory.
Finally, to examine the potential relationship between gesture rate and episodic richness, research assistants computed gesture rate (number of gestures / total narrative time). Specifically, prior to listening to the translation (to remain unbiased), research assistants counted the number of gestures made by each participant when recalling their fear memory. Mean gesture rate across three research assistants was used as the dependent measure. Gesture rates were significantly correlated at p < .001 between each pair of raters (rater a and b: r =.968; rater b and c: r = .911; rater a and c: r = .936).
Results
Experiment 1: Attention to Threat
Threat-Evoked Heart Rate
Participants had an increased heart rate when viewing threatening stimuli (M = 96 bpm. SD = 17) relative to neutral stimuli (M = 91 bpm, SD = 13), t(56) = 2.53, p < .01, d = .23, CI = 0.40, 7.7.
Threat Detection Visual Search
Participants were highly accurate at selecting the target image (Mean accuracy = 96%; SD = 0.14, Mean RT = 2214 ms, SD = 675ms), showing that familiarity with computers or touch screens did not impair overall performance on the task. Our main question was whether the Himba detect threatening targets faster than neutral targets. We examined this in evolutionary old threats (snakes and spiders) and modern threats (guns and knives). To do this, we conducted a 2 X 2 analysis of variance (ANOVA) on RTs with two within-subject variables: target type (threatening vs. neutral) and evolutionary relevance (evolutionary-relevant vs. modern). These analyses were conducted on the entire sample (N = 74), using correct trials only, trials with RT >= 10,000 or <= 400 ms were excluded.
There was an interaction between target type and evolutionary relevance, F(1, 73) = 19.1, p = .00004, ηp2 = .21. Post hoc comparisons showed participants were faster at detecting evolutionary-relevant threat targets compared to their matched neutral targets, mean difference = - 146.6, SE = 61.6, t(-2.380), p = .090, d = -0.277, CI = -0.51, -0.04, and were also faster at detecting modern threat targets compared to their matched neutral targets, mean difference = -490.9, SE = 43.8, t(- 11.200), p = .0001, d = -1.30, CI = -1.61, -0.99 (Figure 2), with a larger effect for modern targets, with a mean difference in magnitude of 344.3. When included as a covariate, there was no impact of age on target type, F(1, 73) = 0.12, p = .727, evolutionary-relevance, F(1, 73) = 1.42, p = .237, or the interaction, F(1, 73) = 0.83, p = .366. Separate T tests were conducted to compare the effect for snakes relative to other stimuli types. There was no difference in the magnitude of the effect for snakes compared to guns, mean snake difference = -598.7, SE = 95.7, mean gun difference = -581.4, SE = 63.5, t(0.156), p = .876, CI = -204, 237, or knives, mean knife difference = 404.2, SE = 62.8, t(1.63), p = .108, CI = -43.9, 433, but the effect for snakes was larger than that of spiders, mean spider difference = -293.5, SE = 67.3, t(7.65), p < .001, CI = 660, 1125.
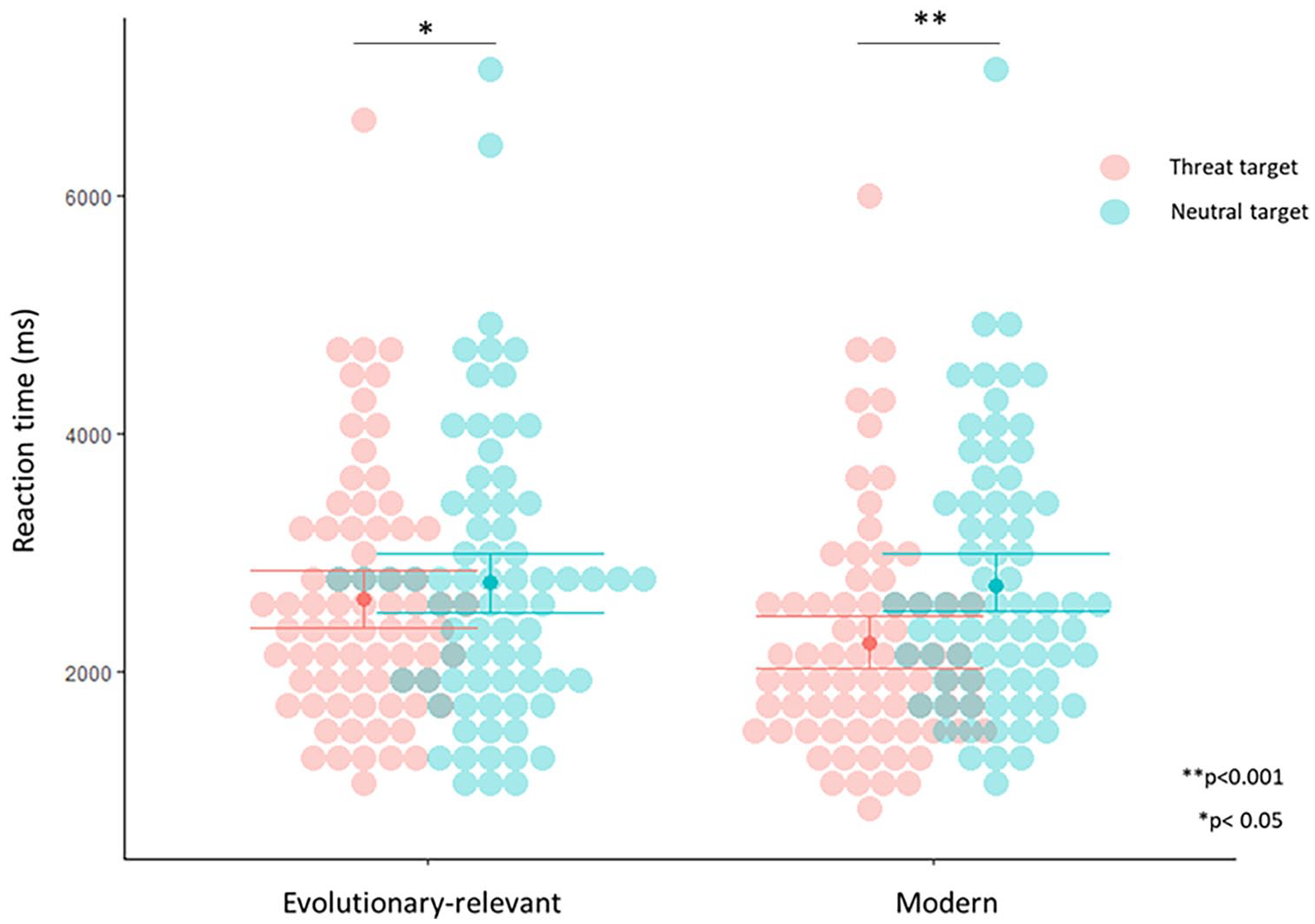
Threat Superiority Effect (TSE).
Experiment 2: Memory of a Threatening Experience
Coder Ratings of Memory Richness
The Himba recalled episodically rich (episodic richness = 3.93, SD = 1.79), and detailed (perceptual richness = 2.1, SD = 0.99) fear memories that included mentions of emotion (emotions/thought richness = 1.77, SD = 1.00). There was variability within the sample, with memories ranging from the lowest (1 for episodic richness, 0 for perceptual richness and emotion/thoughts richness) to highest (6 for episodic richness, 3 for perceptual richness and emotion/thought richness) (Figure 3A). Reported scores are averaged across three raters, ratings were significantly correlated across each pair of raters all at p < .001 (rater a and b: r = .83; rater b and c: r = .68; rater a and c: r = 0.71). Many of the most fearful memories reported involved encounters with either a snake or an elephant, with a smaller number including more modern threats (e.g., guns, car accidents). A summary of the memories can be seen in Figure 5.
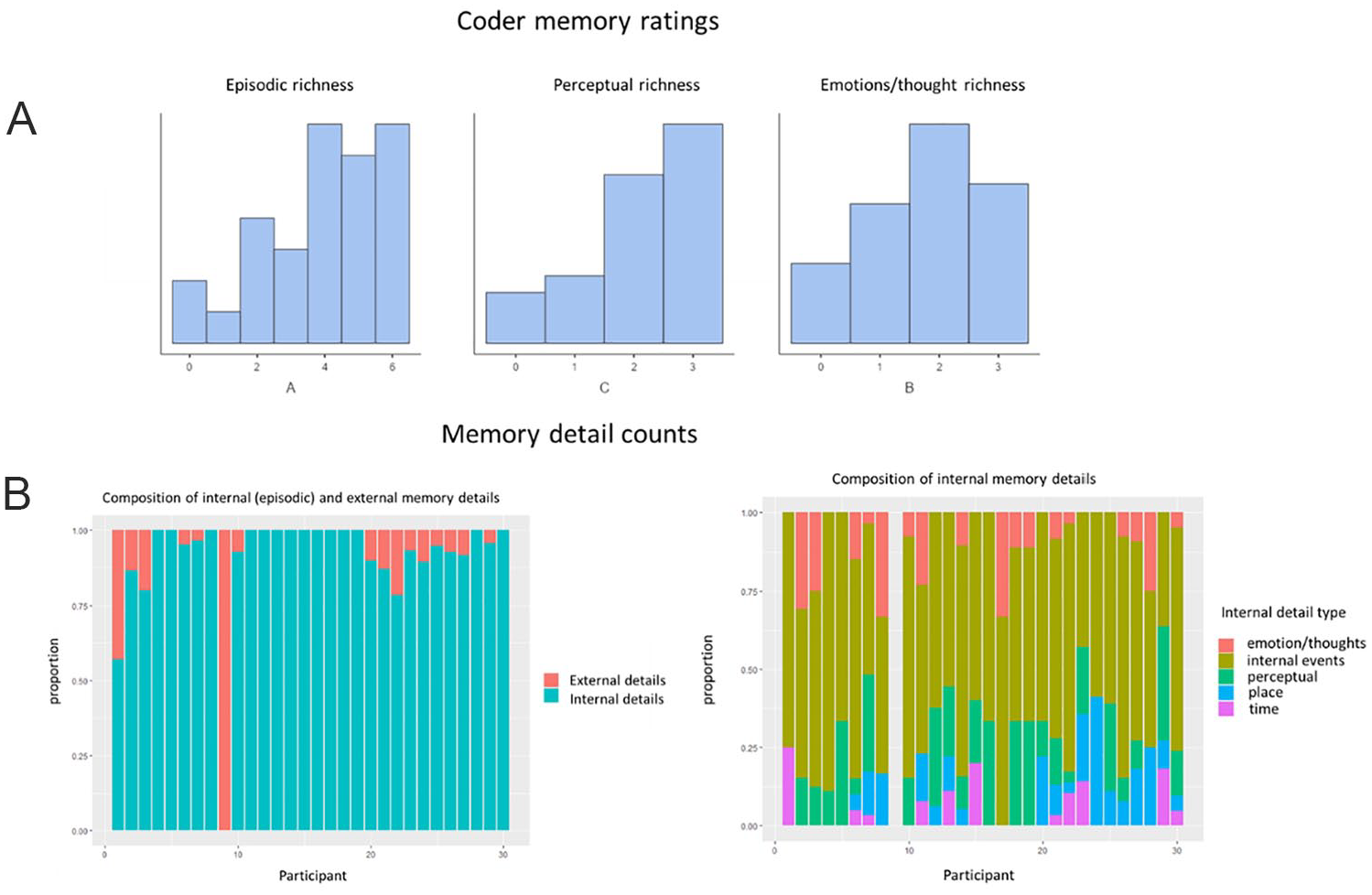
Coder Memory Ratings and Memory Detail Categorization (N = 30). (A) Histograms of Coder Memory Ratings on 3 Dimensions (Left to Right) Episodic Richness, Perceptual Richness and Emotion/Thought Richness. Higher Ratings Indicate Increasing Richness in That Dimension. Memories Are Skewed to Higher Ratings on All Accounts, Indicating That They Are Episodic in Nature and Vivid (Evoke a Sense of Reliving) (B) Composition of Memory Details for Each Participant
Categorization of Memory Details
In line with the coder ratings, the categorizing of memory details reflected the episodic nature of the memories, with within-group variability. Memories were overwhelmingly composed of internal details (M = 91%). Of the internal details, most were internal events (M = 61%), followed by perceptual details (M = 15%), place details (M = 8%), emotion/thought details (M = 8%), and time details (M = 4%). The external details were composed primarily of semantic autobiographical knowledge (M = 46%) and semantic general knowledge (M = 27%) (Figure 3B). The number of internal details was positively correlated with the coder rating of episodic richness (r = 0.64, p =.00014), as was the number of perceptual details with the coder ratings of perceptual richness (r = 0.40, p = .029). The relationship between coder rating of emotion/thought richness and the objective count of emotion/thought details was not significant (r = 0.19, p = .315).
Relationship to Gesture
Gesture rate positively correlated with subjective measures of episodic richness. Specifically, gesture rate correlated with subjective ratings of episodic richness (r = 0.39, p = .032) and perceptual details (r = 0.38, p = .038) but not with emotions/thoughts (r = 0.11, p = .561) (Figure 4). The proportion of internal details did not reach a significant positive correlation with gesture rate (r = .24, p = .201).
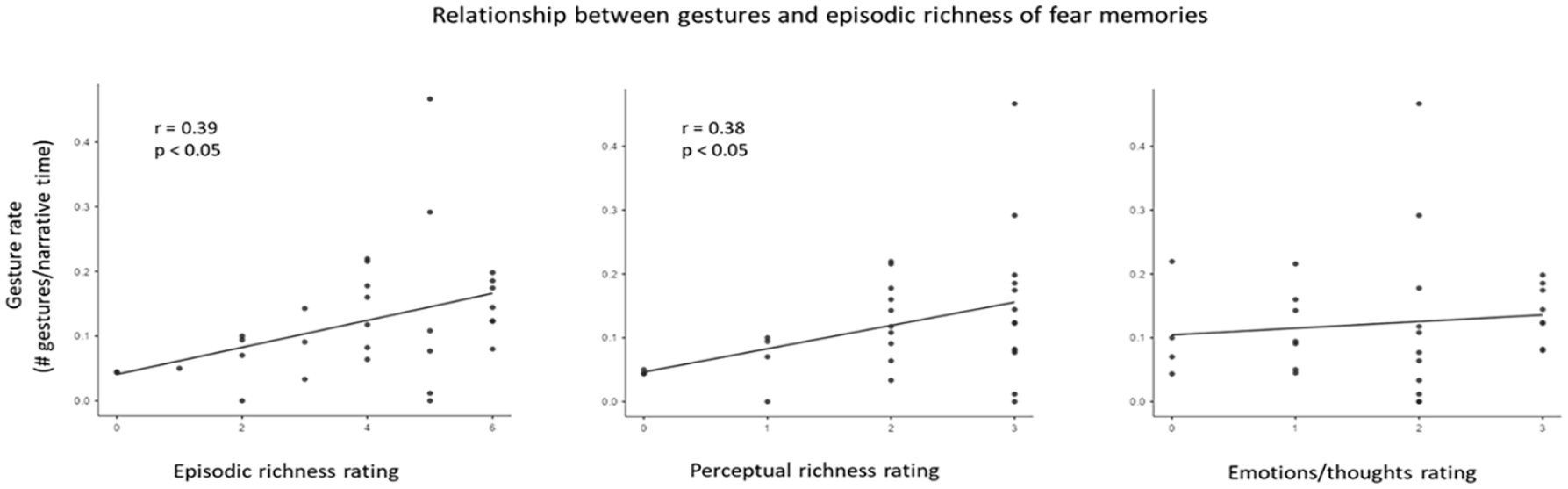
Correlations Between Subjective Ratings of Memories and Gesture Rate.
Discussion
It is widely assumed that fear shapes cognition in adaptive, and therefore universal, ways. However, almost all of the research examining this assumption comes from the West. Here, we addressed this gap across two experiments with a remote African population, the Himba. In Experiment 1, we replicate the classic TSE effect in the Himba. In experiment 2, we show that the Himba have episodic and detailed fear memories.
Experiment 1: Attention to Threat
Here, we find that the Himba show the TSE response for all threat stimuli tested, evolutionary-relevant (snakes and spiders), as well as modern (guns and knives). The speeded response to snakes and spiders could be interpreted as consistent with the fear-module view (rapid responding to these stimulus types is universal due to evolutionary-pressure). We did not find any evidence that this response was enhanced by experience. Specifically, the effect was not larger for snakes relative to the modern threats (guns and knives) though it was larger than the effect for spiders. This is despite the fact that dangerous snakes are fairly common in this region, and many individuals have had fearful encounters with them, as seen in the fear narratives (Figure 5). In fact, somewhat surprisingly the TSE response for modern threats was larger than the effect for evolutionary-relevant threats. One possibility is that reaching out to touch the threats included an approach versus avoidance effect. Specifically, if evolutionary-relevant threats evoke avoidance, but modern threats evoke approach (grabbing that item before your enemy) this could account for the larger effect for modern threats. We think this is unlikely to be the main explanation, because a larger response to modern threats has been documented across attentional bias paradigms, including many that do not employ a touch screen (or a review see Shapouri & Martin, 2022). Specifically a number of studies show, modern threats show just as efficient search as evolutionary-relevant threats, namely, are not sensitive to position effects or increasing grid size (Blanchette, 2006; Brosch & Sharma, 2005; Fox et al., 2007) and the effect has also been found to be larger than evolutionary-relevant threats in paradigms that include target-absent trials and require a same versus different button press response (Blanchette, 2006; Cinq-Mars et al., 2022). Finally, an increased effect size for modern threats has also been found in attention cueing paradigms. For example, Subra et al. (2017) found that participants were faster at selecting a neutral target after it was preceded by a gun than a snake, and Brown et al., (2010) found that using a dot-cueing probe the presentation of modern threats enhanced the P1, a rapid (80–100 ms) visually evoked attentional response more than evolutionary-relevant relevant threats. This finding in multiple paradigms and measures makes it unlikely to be due to an approach avoidance or stimuli similarity confound (unlike the visual search the dot paradigms do not involve distractor-target similarity). Thus, while we do not think this finding is likely due to these confounds, it does require a deeper exploration of stimuli-relevance when conducting these types of studies.
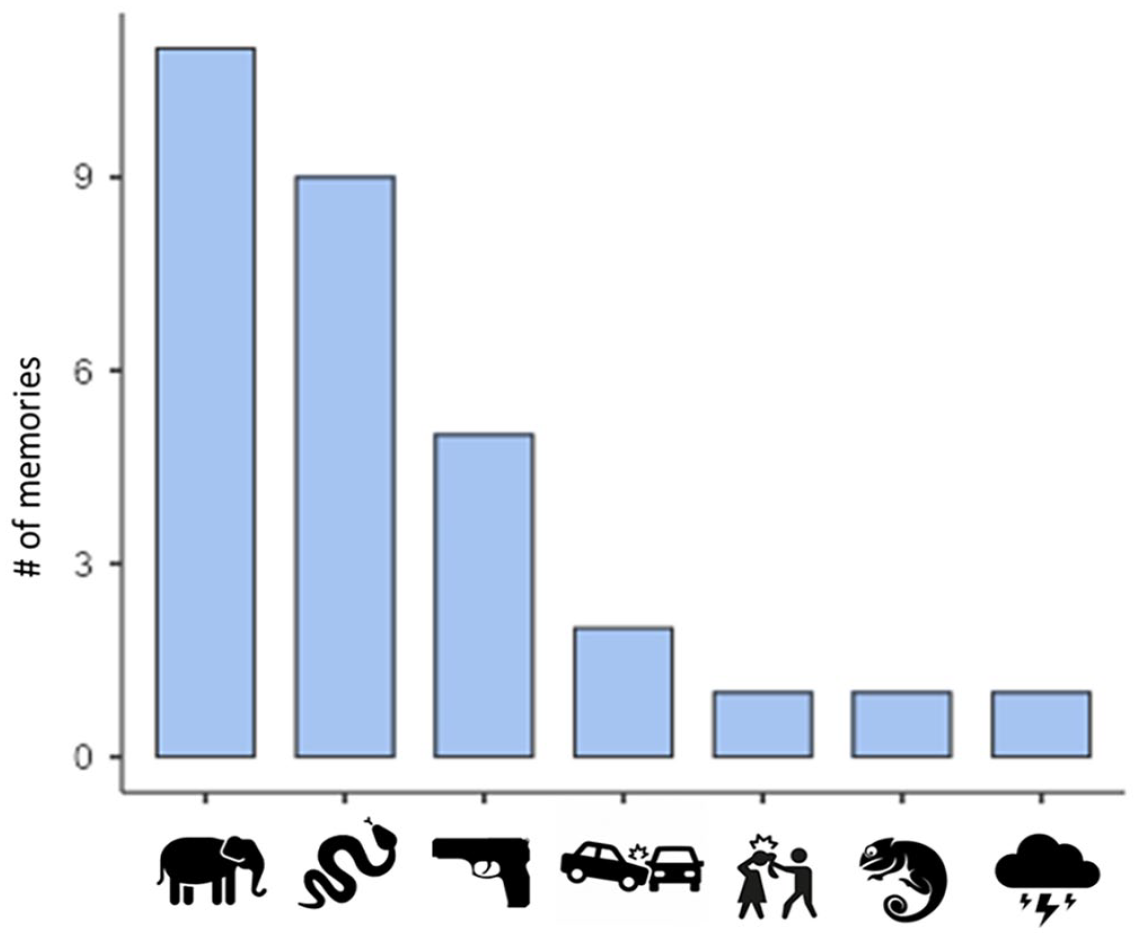
Fear Memory Counts by Topic.
In Western participants, one potential explanation for the enhanced effect is the role of experience, given in-person or media exposure with these stimuli as weapons is likely common. However, how familiar participants are with these items, and how familiar they are with them as threats, remains an open question. Presumably, the Himba are less exposed to guns in general, and knives as violent weapons, but we don’t have data on the frequency of exposure. A small number of fear memories, however, were of episodes of violence involving guns or knives, indicating that exposure might be higher, potentially in Himba with more urban exposure. On the other hand, exposure to snakes in the Himba in our sample was fairly common, 9 out of 30 memories reported were of snake encounters, and Namibia is home to 81 species of snakes, 25 of which are poisonous (Ohorongo, 2021). The Black Mamba, a particularly infamous highly venomous snake is common there. Nevertheless, we did not find evidence of a faster reaction time for snakes in particular, relative to the modern threats tested. Importantly, this does not detract from establishing the effect, but will be important in future studies aimed at understanding the relationship between exposure and threat response. Furthermore, future studies in the West and remote populations will benefit from characterizing exposure to different threat types and examining how experience with and knowledge of threats shapes capture of visual attention.
Experiment 2: Memory of a Threatening Experience
In addition to finding evidence that threats rapidly capture visual attention in the Himba, we find that threatening encounters lead to vivid detail-rich episodic memories. Specifically, each participant was able to generate a distinct episode, and beyond just generating a specific episode, many were rich in episodic, perceptual, and emotional details. Further, these details correlated with ratings of perceptual and episodic richness, and richness was associated with an increased gesture rate. Overall, this pattern is similar to what has been reported for frightening or emotionally negative memories in the West, despite the fact that the content is fairly different. Himba fear memories were most often of encounters in the bush with snakes or elephants. WEIRD memories examined in a similar manner include threat of an impending plane crash (McKinnon et al., 2015), or negative events related to academics, health, or arguments (the most common topics reported in Wardell, Esposito, et al., 2021 fear memory topics were not reported in Hilverman et al., 2016). While there is not yet a comprehensive meta-analysis available for studies in the West employing the AIM method, the proportion of internal details in the Himba fear memories is similar to or higher to what has been reported in the reviewed literature (on average 90% internal details). For example, memories of the near-death experience on the Air Transat flight (McKinnon et al., 2015) and in self-selected most fearful encounters (Hilverman et al., 2016) were composed of about 70% internal details. This suggests that fear may shape memories in similar ways, across cultures and context.
It is not clear whether the episodic vividness reported here is specific to fearful memories or extends to other types of autobiographical memory. Cross-cultural differences for neutral memories have been found between the West and East Asia, with the pattern showing that narratives of neutral memories in Western populations are higher in episodic details. The Himba fear memories appear to be just as episodically detailed to those reported in Western samples, perhaps even more so. One limitation of the current study is that we do not have neutral event memories to compare to the fear memories in the Himba. This will be an important future step, as it will allow us to determine if the structure of episodic memories in general is similar in the Himba, or if the similarities in structure are specific to the way that fear shapes memories. Our working hypothesis is that neutral memories will still be episodic in nature and distinct, but less detailed and vivid than fear memories, as has been reported in the West. Furthermore, having neutral or positive memories to compare will help illuminate reasons for the reported cross-cultural differences between Western and East-Asian autobiographical memory. Specifically, a common explanation of why Western memory narratives is more episodically detailed, is that it is due to the more individualistic culture. If this is true then the Himba memory narratives for neutral events should also be less episodically detailed, given that they are collectivist (Trémolière et al., 2021).
There are few studies using the AIM method to examine the structure of autobiographical memories outside of the West (much less the impact of emotion on autobiographical memory), and none from Africa. Most existing research compares Western healthy adult controls to individuals with different pathologies. The underlying assumption is that episodic memories high in internal details are the default or healthy model. However, the reported cross-cultural differences between healthy East-Asian adults call this into question. Characterizing the structure of AM, and how emotion and pathology shape AM cross-culturally, is therefore a key step forward.
Conclusion
The field of psychology has made impressive advancements in understanding the human mind. However, a serious challenge is the assumption that these advances in understanding are about fundamental aspects of the human mind, and not the WEIRD minds of the participants in these studies. The idea of a “fear system,” or a consistent and distinct way our brain responds to threat, is perhaps one of the most widely assumed universal aspects of cognition. Indeed, the rapid capture of visual attention by threat, as well as enhanced long-term memory for threat, are highly adaptive responses assumed to exist cross-culturally. However, this has not been well-tested outside of the West. Our study makes an important contribution by putting these assumptions to the test, through testing these aspects of cognition in a unique population with different threat experience. We find that the same types of threat stimuli that rapidly capture visual attention in the West also rapidly capture visual attention in the Himba and real-life encounters with understudied threats (snakes and elephants) form rich episodic memories. Clearly, there is more work to be done to understand the relationship between threatening experiences, episodic memories of those experiences, and visual attention to threat. Our study is an important first step toward understanding these relationships and illuminates the benefits of expanding cognition research to remote populations with diverse experiences.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by The Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery grant awarded to Isabelle Blanchette.