
Abstract
The underlying basement membrane is degraded during progression of breast and colon carcinoma. Thus, we imaged degradation of a quenched fluorescent derivative of basement membrane type IV collagen (DQ-collagen IV) by living human breast and colon tumor spheroids. Proteolysis of DQ-collagen IV by HCT 116 and HKh-2 human colon tumor spheroids was both intracellular and pericellular. In contrast, proteolysis of DQ-collagen IV by BT20 human breast tumor spheroids was pericellular. As stromal elements can contribute to proteolytic activities associated with tumors, we also examined degradation of DQ-collagen IV by human monocytes/macrophages and colon and breast fibroblasts. Fibroblasts themselves exhibited a modest amount of pericellular degradation. Degradation was increased 4–17-fold in cocultures of fibroblasts and tumor cells as compared to either cell type alone. Inhibitors of matrix metalloproteinases, plasmin, and the cysteine protease, cathepsin B, all reduced degradation in the cocultures. Monocytes did not degrade DQ-collagen IV; however, macrophages degraded DQ-collagen IV intracellularly. In coculture of tumor cells, fibroblasts, and macrophages, degradation of DQ-collagen IV was further increased. Imaging of living tumor and stromal cells has, thus, allowed us to establish that tumor proteolysis occurs pericellularly and intracellularly and that tumor, stromal, and inflammatory cells all contribute to degradative processes.
Keywords
Introduction
Invasion of colon and breast tumor cells has been linked to degradation of basement membrane proteins by proteases of the matrix metallo, serine, aspartic, and cysteine classes [1]–[9]. Consistent with this is that decreases in staining for the basement membrane proteins laminin and type IV collagen occur at the invasive edges of tumors [10]–[14], suggesting that the basement membrane is being degraded in this region. In order to study degradation in real time and to localize the sites of proteolytic degradation, we established an assay in which we can observe cleavage of quenched fluorescent type IV collagen (DQ-collagen IV) by living tumor cells [15],[16]. Accumulation of fluorescent degradation products is imaged with a confocal microscope and is localized by optically sectioning both the tumor cells and their surrounding matrices. Using this assay, we determined that monolayers of BT20 human breast carcinoma cells degrade DQ-collagen IV pericellularly using matrix metalloproteinases (MMPs), serine proteases, and cysteine proteases [15]. Monolayers of BT549 human breast carcinoma cells degrade DQ-collagen IV intracellularly in an endosomal-lysosomal compartment using cysteine proteases [15], yet, when the BT549 cells were grown as spheroids, they degraded DQ-collagen IV both intracellularly and pericellularly (unpublished observations). We may have underestimated pericellular proteolysis in our previous studies, as they did not include any of the stromal and inflammatory cells associated with tumors in vivo.
Host cells including fibroblasts and macrophages are often the source of proteases in human tumors [5],[17]–[19]. This is true for the three classes of proteases (MMPs, serine proteases, and cysteine proteases) shown to contribute to pericellular degradation by BT20 breast carcinoma cells. For breast carcinoma cells, expression of a number of MMPs has been shown to be a response of host cells to interactions with the tumor cells [18]. The same appears to be true for colon cancer where expression of MMP-2 is induced when stromal fibroblasts interact with colon carcinoma cell lines [20]. Tumor-stromal fibroblast interactions have not been shown to modulate expression of cysteine proteases. On the other hand, tumor-macrophage interactions are associated with increased expression of the cysteine protease cathepsin B, for example, in human colon carcinomas [5],[17]. Such observations suggest that expression of more than one class of proteases in host cells may be modulated by their interactions with tumor cells.
To assess the contribution of host cells to proteolysis in human colon and breast tumors, we employed our confocal assay to image degradation of DQ-collagen IV by spheroids of living human colon (HCT 116 and HKh-2) and breast (BT20) carcinoma cells alone and when cocultured with colon (CCD-112CoN) and breast (12TT) fibroblasts, respectively; monocytes/macrophages (U937); or fibroblasts plus monocytes/macrophages. The colon tumor spheroids degraded DQ-collagen IV pericellularly and intracellularly, and the breast tumor spheroids degraded DQ-collagen IV pericellularly. Pericellular and intracellular proteolysis was markedly enhanced in the cocultures of tumor spheroids with stromal and inflammatory cells. Thus, the ability to image proteolysis of living cells has allowed us to demonstrate that pericellular degradation, intracellular degradation, and multiple cell types all contribute to tumor proteolysis.
Materials and Methods
Materials
DQ-collagen IV, Cell Tracker Orange, Cell Tracker Blue CMAC, Slow Fade Reagent, Live-Dead Kits, and Propidium Iodide Nucleic Acid Stain were purchased from Molecular Probes (Eugene, OR), fluorescein-conjugated affinity-purified donkey antirabbit IgG and normal donkey serum from Jackson ImmunoResearch (West Grove, PA), horseradish peroxidase-labeled goat-antirabbit IgG from Pierce (Rockford, IL), Matrigel from BD Biosciences (Bedford, MA), Agar Noble from DIFCO (Detroit, MI), fetal bovine serum (FBS) from Invitrogen Life Technologies (Carlsbad, CA), the Diff-Quik Stain Set from Dade Behring (Newark, DE), polycarbonate filters (polyvinyl pyrrolidone-free, 13 mm diameter) from Poretics (Livermore, CA), and finder grids from Ted Pella (Redding, CA). The rabbit anti-cathepsin B antibody was produced, affinity-purified, and characterized in our laboratory [21]; the cathepsin B substrate Z-Arg-Arg Cresyl violet 2-Hbr was a kind gift from Dr. Cornelius J.F. Van Noorden (University of Amsterdam, The Netherlands); the cathepsin B inhibitor CA074 [22] and its membrane-permeable derivative CA074Me [23] were purchased from Peptides International (Louisville, KY); and the broad spectrum MMP inhibitors, BB3103 and marimastat, were a kind gift of British Biotech (Oxford, England). Dulbecco's minimal essential medium (DMEM), bovine serum albumin, antibiotics, MEM, DMEM/F12, RPMI 1640, saponin, Nonidet P-40, aprotinin, and all other chemicals unless otherwise stated were purchased from Sigma (St. Louis, MO).
Cell Culture
The HCT 116 and HKh-2 human colon colorectal carcinoma cell lines, a kind gift from Drs. Robert Kerbel (Sunnybrook Health Science Center, Toronto, Canada) and Takehiko Sasazuki (Kyushu University, Higashi, Japan), were cultured in DMEM with 10% FBS. The BT20 human breast carcinoma, CCD-112CoN human colon fibroblast, and U937 human monocyte cell lines were purchased from American Type Culture Collection (Rockville, MD) and cultured according to their recommendations. BT20 cells were cultured in MEM with 10% FBS; CCD-112CoN fibroblasts were cultured in MEM with 0.1 mM nonessential amino acids, 1.5 g/L NaHCO3, 1.0 mM Na pyruvate, Earle's balanced salt solution, and 10% FBS; and U937 monocytes were cultured in RPMI with 10% FBS, 2 mM
Assays for Proteolysis
Cultures of single cell types. Glass coverslips were washed with diluted sulfuric acid (one part sulfuric acid to three parts water) and placed in an oven to dry. Precooled glass coverslips were coated with 30 μL of Matrigel containing 25 μg/mL of DQ-collagen IV and left in a 37°C incubator for 15 min to solidify. Cells (3–4 × 10) were plated on the top of Matrigel and cultured in the media indicated above, including serum, for 40–44 hr. Broad spectrum MMP inhibitors [BB3103 (10 μM final concentration) or marimastat (10 μM final concentration)], a plasmin inhibitor (aprotinin, 1 μM final concentration), and cathepsin B inhibitors [CA074 (10 μM final concentration) and CA074Me (10 μM final concentration)] were added when specified to identify the proteases responsible for degradation of DQ-collagen IV. At the concentrations used and over the entire time course of the assay, neither the inhibitors nor the diluent (DMSO, 0.1% final concentration) affected cell viability as determined using a Live-Dead Kit and by an absence of staining with propidium iodide. Cultures of living cells were observed at intervals during the 44-hr period for fluorescent degradation products on a Zeiss LSM 310 using a 40 × water immersion objective.
Preformed spheroids. BT20 human breast carcinoma cells (3–5 × 10) were collected from a T-25 flask by trypsinization, suspended in 15 mL of complete growth medium and seeded into a 100-mm culture dish precoated with 0.75% Agar Noble in serum-free medium [24]. The cells aggregated into spheroids reaching a size of 100–200 μm after a period of 2–3 days.
Cocultures. Fibroblasts (colon or breast), monocytes, and macrophages were grown in T-25 flasks to ˜80% confluency, washed with phosphate-buffered saline (PBS), and stained with 5 μM of Cell Tracker Orange or Blue in serum-free medium for 45 min in a 37°C incubator. The prelabeled cells were washed with PBS and incubated with prewarmed medium containing 10% FBS for 1 hr in a 37°C CO2 incubator. Single cell suspensions of HCT 116, HKh-2, or BT20 tumor cells (3–4 × 104/coverslip) were seeded in 35-mm Petri dishes on two or three coverslips coated with Matrigel containing DQ-collagen IV. The dishes were placed in a 37°C CO2 incubator for 30–60 min until the cells attached. A 20-μL aliquot of prelabeled fibroblasts (3–4 × 103) and/or of prelabeled monocytes/macrophages (7–8 × 103) was added and allowed to attach. Then, normal culture medium was added and the cocultures were observed for fluorescent degradation products as described above. Cocultures were incubated in the absence or presence of protease inhibitors as described above for cultures of single cell types.
Viability Tests
Cells were grown on Matrigel-coated coverslips for 44 hr, washed 3 times with 37°C Tris-buffer and stained with 3 μM of propidium iodide nucleic acid stain in Tris-buffer (100 mM Tris, pH 7.4, 150 mM NaCl, 1 mM CaCl2, 0.5 mM MgCl2, 0.1% Nonidet P-40) for 15 min. Cells were washed 3 times with Tris-buffer, then serum-free medium was added, and the cells were observed with a Zeiss LSM 310 using a 40 × water immersion objective.
Invasion Assays
Polycarbonate filters (12-μm pores) were coated with 1% gelatin and dried overnight. Matrigel (50 μg/filter, i.e., 0.23 μg/mm2) was spread over the entire surface of the gelatin-coated filters and dried to a thin film in a laminar flow hood at room temperature. The Matrigel-coated filters were placed in Boyden chambers, and media, conditioned for 24 hr by incubating log-phase NIH-3T3 cells in DMEM supplemented with 1% FBS, was added to the lower compartment as a chemoattractant. Tumor cells (5 × 104) in 200 μL of DMEM with 0.1% BSA were placed on the filters and incubated overnight at 37°C in a CO2 incubator. In order to identify the proteases responsible for invasion, tumor cells were preincubated for 15 min with inhibitors before placing them on the filters in the presence of medium containing 0.1% BSA and the inhibitor. For studies with aprotinin, the tumor cells were in medium with 1% FBS in order to provide a source of plasminogen. The filters were removed, air dried, and stained with Diff-Quik prior to removing Matrigel and the cells from the top surface with a cotton swab. This left on the filter only those cells that had traversed the matrix barrier [24]. Invasion was quantified by counting cells on a Nikon TMS with a 10 × objective using finder grids. Experiments were done in triplicate and each filter was counted three times.
Immunocytochemistry
Spheroids of BT20 cells were grown on Matrigel-coated coverslips containing 20 μg/mL of DQ-collagen IV for 40–44 hr and then stained for surface cathepsin B according to our published procedures [15]. Fixation and subsequent steps were performed at 4°C and in the absence of detergents in order to visualize only surface cathepsin B. Spheroids were fixed for 10 min in 3.7% formaldehyde in PBS. After washing with PBS, nonspecific binding was blocked for 45 min by incubating with PBS-2 mg/mL bovine serum albumin. The spheroids were then incubated with primary antibody (230 μg/mL of rabbit antihuman liver cathepsin B IgG) or preimmune rabbit IgG (225 μg/mL) for 2 hr. After washing with PBS, the spheroids were incubated for 1 hr with fluorescein-conjugated affinity-purified donkey antirabbit IgG at 20 μg/mL and containing 5% normal donkey serum. Spheroids were then washed, mounted upside-down with SlowFade antifade reagent on glass slides, and observed with a Zeiss LSM 310 confocal microscope.
Cytochemistry
HCT 116 and HKh-2 cells (3–4 × 10) were plated on DQ-collagen IV/Matrigel-coated coverslips and grown for 44 hr. The colon carcinoma spheroids were imaged for fluorescent degradation products of DQ-collagen IV as described above. Then, a highly selective cathepsin B substrate, Z-Arg-Arg Cresyl violet 2-HBr [25] in DMEM, was added in order to determine the localization of cathepsin B activity using our published protocol [15],[24]. Cresyl violet fluorescence and colocalization of the Cresyl violet fluorescence with the fluorescent degradation products of DQ-collagen IV were imaged on a Zeiss LSM 310 confocal microscope using a 40 × water immersion objective as described [15].
Quantification and Statistical Analyses
Fluorescence intensities and areas in the presence and absence of various cell types were quantified from confocal fluorescence images by three independent observers using Image J Software (National Institutes of Health, Bethesda, MD). SuperANOVA version 1.11 (Abacus) was used to determine significant differences.
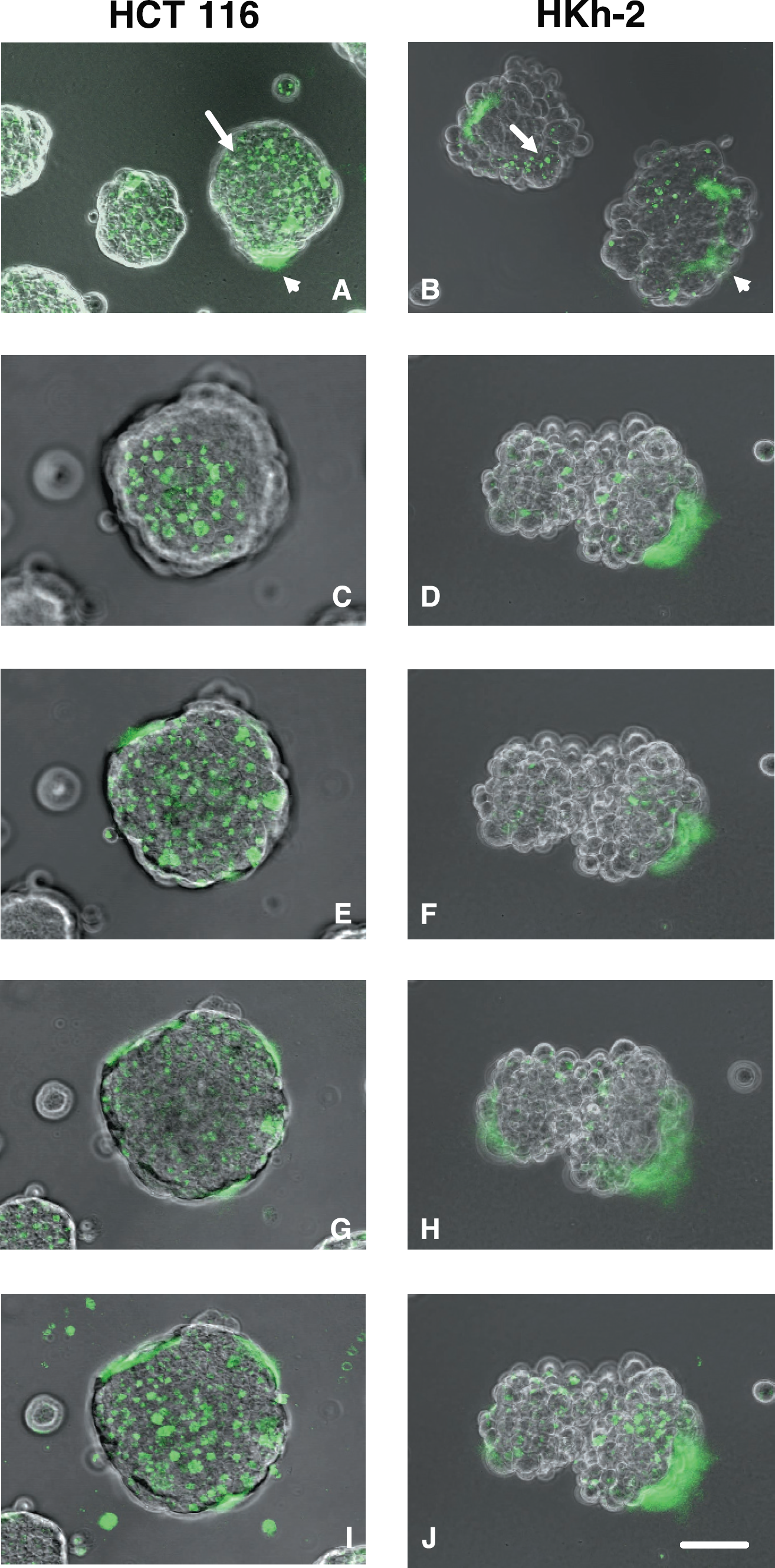
Spheroids of living colon carcinoma cells degrade a quenched fluorescent derivative of type IV collagen (DQ-collagen IV) both pericellularly (A and B, arrowheads) and intracellularly (A and B, arrows). Fluorescence images of DQ-collagen IV cleavage products (green) are superimposed on phase images of spheroids of parental HCT 116 cells (left panels) and of the less invasive daughter HKh-2 cells in which Ki-ras had been deleted (right panels) [26]. (A and B) Optical sections taken at an extended depth of focus of several representative HCT 116 and HKh-2 spheroids, respectively. Panels C-J are images of the same HCT 116 spheroid and HKh-2 spheroid taken at the top (C and D), middle (E and F) and bottom (G and H) of spheroids. H and I are images of the same HCT 116 and HKh-2 spheroids, respectively, taken using an extended depth of focus. Single cell suspensions of tumor cells were plated on DQ-collagen IV/Matrigel coated coverslips and imaged at 40–44 hr. Bar = 10 μm.
Results
Imaging Proteolysis by Living Colon Carcinoma Cells
We compared the proteolytic capabilities of two human colon carcinoma cell lines: The parental HCT 116 cell line that expresses a mutant Ki-ras allele and is malignant and invasive and the HKh-2 daughter cell line in which the Ki-ras allele was deleted by homologous recombination resulting in a less malignant and invasive cell line [26,27]. We confirmed these differences in invasiveness under the conditions of our invasion assay: HCT 116 cells invaded and HKh-2 cells did not (data not shown). We plated the two human colon carcinoma cell lines on glass coverslips coated with an intramolecularly quenched fluorescent basement membrane protein substrate, DQ-collagen IV, mixed with the reconstituted basement membrane Matrigel. Both cell lines migrated into spheroids with the HKh-2 cells forming somewhat less tightly packed spheroids (Figure 1, right panels) than did the parental HCT 116 cells (Figure 1, left panels). Degradation of the DQ-collagen IV by the living colon carcinoma cells and accumulation of green fluorescent cleavage products was visualized by confocal microscopy in optical sections taken at various levels through the spheroids. Discrete areas of intracellular degradation were abundant in HCT 116 spheroids. This was particularly evident in images taken at the top and middle of the HCT 116 spheroids (Figure 1C and E) and in images taken at an extended depth of focus (Figure 1A and I). Areas of pericellular proteolysis were present in images taken at the middle and bottom of the HCT 116 spheroids (Figure 1E and G) and images taken at an extended depth of focus (Figure 1A and I). The amount of intracellular degradation was considerably less in HKh-2 spheroids (Figure 1, right panels) than in HCT 116 spheroids (Figure 1, left panels). Pericellular degradation in the HKh-2 spheroids ranged from comparable to that in HCT 116 spheroids to more than in HCT 116 spheroids; pericellular degradation could be observed in optical sections of the HKh-2 spheroids taken at all levels (Figure 1B, D, F, H, and J). Thus, the less invasive HKh-2 cells differed from the HCT 116 cells primarily in regard to intracellular rather than pericellular proteolysis, suggesting that intracellular proteolysis may play a role in colon carcinoma invasion.
Using protease inhibitors, we have previously established that living BT20 human breast carcinoma cells in monolayer cultures use MMPs, serine proteases of the plasminogen cascade, and the cysteine protease cathepsin B to degrade DQ-collagen IV pericellularly, whereas living BT549 human breast carcinoma and U87 human glioma cells use the cysteine protease cathepsin B to degrade DQ-collagen IV intracellularly [15,16]. We, therefore, tested the ability of protease inhibitors to affect proteolysis of DQ-collagen IV by HCT 116 human colon carcinoma spheroids (Figure 2). BB3103, a broad spectrum MMP inhibitor, had no effect on intracellular proteolysis, but did reduce pericellular proteolysis (Figure 2B). The plasmin inhibitor aprotinin did not reduce either pericellular or intracellular proteolysis (Figure 2C). CA074Me, a highly selective and cell-permeable cathepsin B inhibitor [23], dramatically reduced intracellular proteolysis as might have been expected for an inhibitor that can enter the cells; however, it also reduced pericellular proteolysis (Figure 2D). For the HKh-2 daughter spheroids, similar reductions in degradation of DQ-collagen IV were observed with the three inhibitors (data not shown). Our results, thus, suggest that MMPs and cathepsin B were both involved in pericellular degradation of DQ-collagen IV by living HCT 116 and HKh-2 colon carcinoma spheroids, whereas cathepsin B appeared to be the primary enzyme involved in intracellular degradation of DQ-collagen IV.
In BT549 human breast carcinoma [15] and U87 human glioma [16] cells, fluorescent cleavage products due to the intracellular degradation of DQ-collagen IV accumulate within an endosomal-lysosomal compartment of the cells, a compartment that also stains for the lysosomal cysteine protease cathepsin B. In order to determine whether the intracellular compartment staining for degraded DQ-collagen IV in the two colon carcinoma cell lines was an endosomal-lysosomal compartment containing active cathepsin B, we incubated the cells with DQ-collagen IV and a cathepsin B-selective Cresyl violet substrate that fluoresces red upon cleavage by cathepsin B [25]. Cleavage products of DQ-collagen IV were present intracellularly in the HCT 116 (Figure 3A) and the HKh-2 (Figure 3B) cells as discrete spots of green fluorescence. Substantially fewer fluorescent spots were present in the HKh-2 cells. Staining for active cathepsin B was present in the two cell lines as discrete spots of red fluorescence (Figure 3C and D). When the degradation products and cathepsin B activity were merged (Figure 3E and F), we observed colocalization (yellow fluorescence), suggesting that DQ-collagen IV degradation took place in a vesicular compartment that contained active cathepsin B. Substantially less colocalization was observed in HKh-2 spheroids (Figure 3F) than in HCT 116 spheroids (Figure 3E), a finding consistent with less intracellular DQ-collagen IV degradation in the HKh-2 spheroids. Taken together, our results are consistent with cathepsin B participating in intracellular degradation of DQ-collagen IV in the living human colon carcinoma cells.

Inhibitors of MMPs (B) and cathepsin B (D) reduce degradation of DQ-collagen IV by living HCT 116 human colon carcinoma spheroids. Fluorescence images of DQ-collagen IV degradation products (green) taken at an extended depth of focus are superimposed on phase images of the living spheroids. (A) Control; (B) BB3103 (10 μM); (C) aprotinin (1 μM); and (D) CA074Me (10 μM). Single cell suspensions of tumor cells were mixed with inhibitors, plated on DQ-collagen IV/Matrigel coated coverslips and imaged at 40–44 hr. Bar = 10 μm.
Imaging Proteolysis by Living Breast Carcinoma Cells
We had previously established that in monolayer cultures living BT20 human breast carcinoma cells degrade DQ-collagen IV pericellularly, whereas living BT549 human breast carcinoma cells degrade DQ-collagen IV intracellularly [15]. Here, we examined the degradation of DQ-collagen IV by spheroids of BT20 cells (Figure 4). The BT20 spheroids were less cohesive than the colon carcinoma spheroids; groups of cells, as well as single cells, could be seen invading from the BT20 spheroids into the surrounding Matrigel (Figure 4A-F). Fluorescent cleavage products were present pericellularly around the spheroids (Figure 4B, C, H, and I) and around the invading groups of cells (Figure 4A, B, C, and F). Degradation of DQ-collagen IV could be visualized surrounding the extended process of a single cell invading through the Matrigel (Figure 4D-F). We had already established that BT20 breast carcinoma cells express a large quantity of cathepsin B on their surface when they are grown on monolayer culture [15]. We confirmed that cathepsin B was expressed on the outer surface of BT20 breast carcinoma spheroids (Figure 4G). Furthermore, the pattern of staining for cathepsin B resembled that of the DQ-collagen IV degradation products (Figure 4H). Indeed, the merged images exhibit areas of yellow fluorescence that indicate colocalization (Figure 4I) of cathepsin B and DQ-collagen IV degradation. These images suggest that cathepsin B was localized such that it could participate in the degradation of DQ-collagen IV by BT20 spheroids. We have already shown in BT20 monolayer cultures that cathepsin B is part of a proteolytic cascade also involving MMPs and the plasminogen cascade [15].
Imaging Proteolysis by Living Inflammatory and Stromal Cells
Both inflammatory and stromal cells may contribute to tumor proteolysis. In colon carcinoma, the serine protease urokinase plasminogen activator is expressed primarily by stromal cells, yet the receptors for urokinase are present on the surface of tumor, stromal, and inflammatory cells [28]–[30]. In breast carcinoma, Matrisian et al. [18] have shown that most MMPs are expressed by tumor-associated fibroblasts rather than by the tumor cells. The cysteine protease cathepsin B on the other hand is expressed by infiltrating macrophages in colon carcinoma [5,17] and breast carcinoma [31] and by breast stromal cells [31,32]. To evaluate the potential for host cells contributing to proteolysis, we assessed the ability of living human U937 monocytes and macrophages (phorbol ester-stimulated monocytes) and fibroblasts isolated from human colon (CCD-112CoN) and breast (12TT) tissues to degrade DQ-collagen IV. U937 monocytes did not degrade DQ-collagen IV (Figure 5A) and U937 macrophages degraded DQ-collagen IV intracellularly (Figure 5B). In contrast, fibroblasts from normal colon (Figure 5C) and breast (Figure 5D,E) tissues degraded DQ-collagen IV pericellularly. Inhibitors of MMPs (Figure 5F), the plasminogen cascade (Figure 5G) and cathepsin B (Figure 5H) all reduced to a comparable extent the pericellular degradation of DQ-collagen IV by spheroids of 12TT breast fibroblasts (Figure 5E). Thus, the fibroblasts like the carcinoma cells use more than one class of proteases to degrade DQ-collagen IV.

Fluorescent degradation products of DQ-collagen IV (A and B) colocalize with staining for cathepsin B activity (C and D) in living human colon carcinoma spheroids (HCT 116: left panels; HKh-2: right panels). In the bottom panels, the two fluorescent images are merged and superimposed on a phase contrast image of the tumor spheroids. Yellow fluorescence indicates colocalization of the green fluorescence due to cleavage of DQ-collagen IV and the red fluorescence due to cathepsin B activity (i.e., hydrolysis of the Z-Arg -Arg Cresyl violet substrate). Single cell suspensions of tumor cells were plated on DQ-collagen IV/Matrigel coated coverslips and imaged at 40–44 hr. Bar = 10 μm.

Spheroids of living BT20 human breast carcinoma cells degrade DQ-collagen IV pericellularly. Fluorescent cleavage products are concentrated in regions where groups of cells are invading from the spheroid into the surrounding Matrigel (A-C), surround individual cells and cell processes invading through the Matrigel (F), and surround spheroids (H and I). Panels A-C represent optical sections at the top, middle, and bottom of a single BT20 spheroid. Panels D-F represent optical sections of cells invading from a BT20 spheroid into the Matrigel with D, E, and F taken at successively further distances from the spheroid (for orientation, an arrow indicates the nucleus of a single invading cell in the three images). Panels G, H, and I are images of fixed cells and illustrate indirect immunofluorescent staining for pericellular cathepsin B (red) in an unpermeabilized BT20 spheroid (G); fluorescent degradation products (green) of DQ-collagen IV (H); and a merged image (I) in which the fluorescent images G and H (taken at an extended depth of focus) are superimposed on a phase image. The yellow fluorescence in Panel I indicates colocalization of DQ-collagen IV cleavage products and cathepsin B protein. BT20 spheroids were plated on DQ-collagen IV/Matrigel coated coverslips and imaged at 40–44 hr. Bar = 10 μm.
Imaging Proteolysis by Cocultures of Living Inflammatory and Stromal Cells and Carcinoma Cells
Interactions among the various cell types in solid tumors (tumor, stromal, and inflammatory cells) may enhance tumor proteolysis. Therefore, we determined whether proteolysis of DQ-collagen IV was altered when human HCT 116 and HKh-2 colon and BT20 breast carcinoma cells were cocultured with human monocytes, macrophages, or fibroblasts from normal colon and breast tissues. In cocultures of HCT 116 colon carcinoma cells and CCD-112CoN colon fibroblasts, proteolysis of DQ-collagen IV was increased pericellularly and intracellularly (Figure 6A), whereas in cocultures of HKh-2 colon carcinoma cells and CCD-112CoN colon fibroblasts, proteolysis of DQ-collagen IV was increased pericellularly (Figure 6B). Pericellular, but not intracellular, proteolysis also was greatly increased in cocultures of BT20 breast carcinoma cells and 12TT breast fibroblasts (Figure 7A). In the breast cocultures, proteolysis of DQ-collagen IV could be seen to be highly concentrated in regions where the fibroblasts were interacting with the tumor cells (Figure 7A and B). Pericellular proteolysis was not increased when either the HCT 116 or the HKh-2 colon carcinoma cells or the BT20 breast carcinoma cells were cocultured with U937 monocytes (Figure 6C and D and Figure 7C, respectively). Total proteolysis was increased when U937 macrophages were cocultured with HCT 116 or HKh-2 colon carcinoma or BT20 breast carcinoma cells (Figure 6E and F and Figure 7D, respectively). The increase in total proteolysis included intracellular proteolysis in the macrophages in the cocultures as indicated by the colocalization of the fluorescent label for the macrophages (red for cocultures with colon carcinoma and blue for cocultures with breast carcinoma) with the green fluorescent cleavage products of DQ-collagen IV. The colocalization is seen as yellow fluorescence in Figure 6E and F and aqua fluorescence in Figure 7D. Three independent people quantified (see Materials and Methods and Table 1) the increases in total proteolysis (pericellular + intracellular) in the cocultures of carcinoma cells and stromal or inflammatory cells as illustrated in Figures 6 and 7. The increase in mean or relative fluorescence (i.e., proteolysis) for cocultures of HCT 116 colon carcinoma cells with CCD-112CoN colon fibroblasts was fourfold and with U937 macrophages two- to threefold and for cocultures of BT20 breast carcinoma cells with 12TT breast fibroblasts was ˜15-fold and with U937 macrophages was two- to fourfold. In contrast, increases in relative fluorescence of cocultures of the HKh-2 daughter cells with fibroblasts and macrophages were only 10% and 40%, respectively. Inhibitors of MMPs (Figure 8C and D), the plasminogen cascade (Figure 8E and F), and cathepsin B (Figure 8G and H) reduced the increased pericellular proteolysis in the HCT 116 colon carcinoma/CCD-112CoN colon fibroblast (left panels) and in the BT20 breast carcinoma/12TT breast fibroblast (right panels) cocultures. Controls for the colon and breast cocultures are illustrated in Panels A and B, respectively, of Figure 8. The cathepsin B inhibitor also reduced the increased intracellular proteolysis in the HCT 116 colon carcinoma/CCD-112CoN colon fibroblast coculture (cf. Figure 8A and G). The cocultures of carcinoma cells and fibroblasts like the separate cultures of carcinoma and fibroblasts use more than one protease to degrade extracellular matrix proteins. For the pericellular degradation of DQ-collagen IV the proteases were MMPs, the plasminogen cascade and cathepsin B and for intracellular degradation of DQ-collagen IV the protease was cathepsin B. In contrast to the breast carcinoma cells alone, the colon carcinoma cells alone did not use proteases of the plasminogen cascade to degrade DQ-collagen IV pericellularly.

Living inflammatory and stromal cells degrade DQ-collagen IV. U937 monocytes are unable to degrade DQ-collagen IV (A). U937 human macrophages degrade DQ-collagen IV intracellularly (B), whereas CCD-112CoN human colon fibroblasts (C) and 12TT human breast fibroblasts (D) degrade DQ-collagen IV pericellularly. Inhibitors of MMPs (F), the plasminogen cascade (G), or cathepsin B (H) reduce degradation of DQ-collagen IV by 12TT human breast fibroblast spheroids [(E) control; (F) BB3103, 10 μM; (G) aprotinin, 1 μM; and (H) CA074 and CA074Me, 10 μM each]. Fluorescence images of DQ-collagen IV degradation products (green) taken at an extended depth of focus are superimposed on phase images of the living cells. Single cell suspensions of the monocytes, macrophages or fibroblasts (alone or mixed with inhibitors) were plated on DQ-collagen IV/Matrigel coated coverslips and imaged at 40–44 hr. Bar = 10 μm.
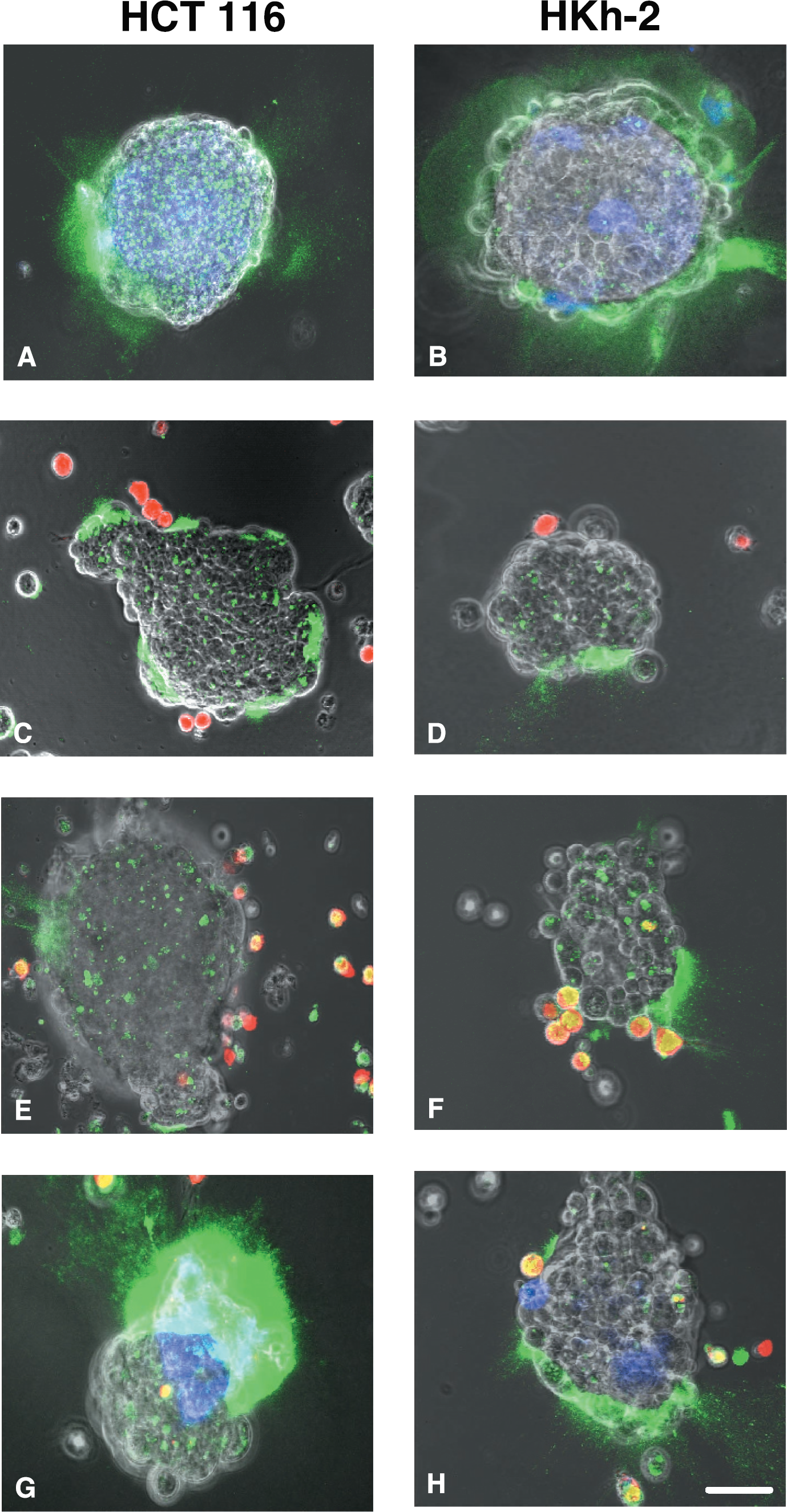
Degradation of DQ-collagen IV is increased in cocultures of living HCT 116 (left panels) human colon carcinoma spheroids and CCD-112CoN human colon fibroblasts (A) or fibroblasts plus U937 human macrophages (G). Intracellular degradation as well as pericellular degradation is increased in living HCT 116 spheroids cocultured with fibroblasts (cf. Panel A with Panels C and E). Coculturing HKh-2 (right panels) human colon carcinoma spheroids with CCD-112CoN human colon fibroblasts (B) increases pericellular degradation of DQ-collagen IV, yet, degradation is not further increased by the addition of U937 human macrophages (H). Although coculturing either HCT 116 or HKh-2 human colon carcinoma spheroids with U937 monocytes (C and D) or macrophages (E and F) does not affect pericellular degradation of DQ-collagen IV, the macrophages themselves degrade DQ-collagen IV intracellularly (E and F). Single cell suspensions of tumor cells were plated on DQ-collagen IV/Matrigel-coated coverslips and incubated until attached (see Materials and Methods for details). Single cell suspensions of stromal and/or inflammatory cells were then added and the living cocultures imaged at 40–44 hr. Fibroblasts and monocytes/macrophages were prelabeled with Cell Tracker Blue and Cell Tracker Orange so that they fluoresce blue and red, respectively. Fluorescence images were taken at an extended depth of focus and are superimposed on phase images of the living cocultures. Bar = 10 μm.
As both stromal and inflammatory cells are found in solid tumors, we also determined whether proteolysis of DQ-collagen IV was affected when human HCT 116 and HKh-2 colon and BT20 breast carcinoma cells were cocultured with human fibroblasts from normal colon and breast tissues, respectively, and in addition human U937 macrophages. Pericellular proteolysis was dramatically enhanced by adding macrophages to the HCT 116 colon carcinoma/CCD-112CoN colon fibroblast cocultures (Figure 6G) and the BT20 breast carcinoma/12TT breast fibroblast cocultures (Figure 7E and H). The increases in mean and relative fluorescence in the cocultures of carcinoma cells and host cells (Table 1) were 10-fold for HCT 116/CCD-112CoN/U937 cocultures and ˜20-fold for BT20/12TT/U937 cocultures. In the breast cocultures, proteolysis of DQ-collagen IV could be seen to be concentrated in regions of interaction between the fibroblasts and the tumor cells (Figure 7F and H). The presence of U937 macrophages in the cocultures of HKh-2 colon carcinoma/CCD-112CoN colon fibroblasts did not substantially increase pericellular proteolysis of DQ-collagen IV (Figure 6H and Table 1).
Parallel between Invasion through Matrigel and Proteolysis of DQ-Collagen IV
We have shown that the cysteine protease cathepsin B was one of the proteases responsible for degradation of DQ-collagen IV by living colon and breast carcinoma cells, stromal fibroblasts, and macrophages (Figures 2,5, and 8; data not shown; and Ref. [15]) by using two highly selective cathepsin B inhibitors, the impermeant form CA074 and the cell-permeable CA074Me, to abrogate pericellular and intracellular cathepsin B activity, respectively. The HKh-2 colon carcinoma cells did not invade through Matrigel (data not shown) and were less effective in degrading DQ-collagen IV than the HCT 116 colon carcinoma cells (cf. left and right panels of Figure 1). The number of HCT 116 cells invading through Matrigel was reduced from 161 ± 35 to 8 ± 7 (mean ± SD) by a cocktail of the two cathepsin B inhibitors (CA074 + CA074Me, each at a final concentration of 10 μM) that reduced proteolysis of DQ-collagen IV. We verified that the inhibitors totally abrogated cathepsin B activity under the conditions of our assays, including the 44-hr time course of the degradation assay, yet, did not affect cell viability. The cathepsin B activity in cell lysates and conditioned media was 0 ± 1% and 0 ± 2% (mean ± SD), respectively, of the activity in the absence of the inhibitors. Our studies, thus, support a role for cathepsin B in invasion of the HCT 116 human colon carcinoma cells through Matrigel and in the accompanying proteolysis of DQ-collagen IV.
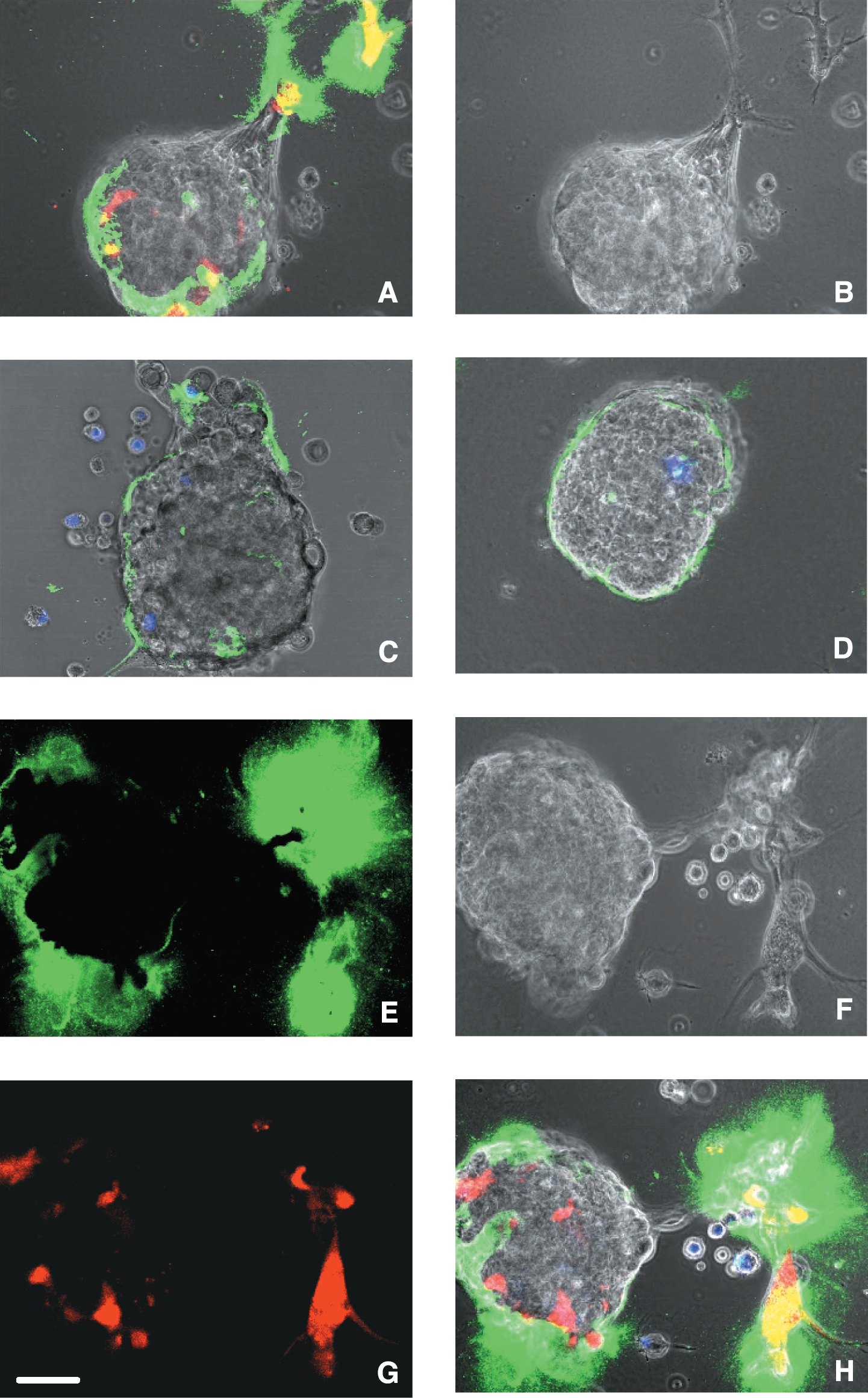
Degradation of DQ-collagen IV is increased in cocultures of living BT20 human breast carcinoma spheroids and 12TT human breast fibroblasts (A) or fibroblasts plus U937 human macrophages (E and H). Pericellular proteolysis (green fluorescence) is greatest in regions of interaction (yellow fluorescence) between tumor cells and fibroblasts (red fluorescence). For orientation, phase images of BT20/12TT (B) and BT20/12TT/U937 macrophage cocultures (F) are depicted. Coculturing BT20 human breast carcinoma spheroids with U937 monocytes (C) or macrophages (D) does not affect pericellular degradation of DQ-collagen IV. Some macrophages degrade DQ-collagen IV intracellularly (D). Single cell suspensions of tumor cells were plated on DQ-collagen IV/Matrigel-coated coverslips and incubated until attached (see Materials and Methods for details). Single cell suspensions of stromal and/or inflammatory cells were then added and the living cocultures imaged at 40–44 hr. Fibroblasts and monocytes/macrophages were prelabeled with Cell Tracker Orange and Cell Tracker Blue so that they fluoresce red (A and G) and blue (C, D, and H), respectively. Fluorescence images of the living cocultures were taken at an extended depth of focus. In Panels A, C, and H, fluorescence images are superimposed on phase images of the living cocultures. Bar = 10 μm.
Interaction of Carcinoma Cells with Stromal and/or Inflammatory Cells Increases Proteolysis of DQ-Collagen IV

a Mean fluorescence ± SD is in arbitrary units with each unit equal to the sum of the gray scale value for the pixels within an image using Image J Software.
b Area is in Px2 units and represents the total areas in which fluorescence was assessed, that is, area of spheroids and associated cells + areas of pericellular fluorescence in which there were not cells.
c Relative fluorescence is the mean fluorescence that has been normalized to the smallest area measured for each of the carcinoma cell lines.
d TC = tumor cells.
e The area used for normalization.
* Indicates p <.05 between TC/TC + monocytes and other combinations.
** Indicates p <.05 between TC + fibroblasts and TC + fibroblasts + macrophages.
*** Indicates p < .05 between TC + macrophages and TC + fibroblasts + macrophages.
Cathepsin B is not the only protease that plays a role in invasion and collagen IV degradation by the HCT 116 human colon carcinoma cells. A broad spectrum MMP inhibitor (marimastat, 10 μM final concentration) reduced the number of HCT 116 cells invading through Matrigel from 127 ± 15 to 1 ± 2 (mean ± SD). In contrast, the plasmin inhibitor aprotinin (1 μM final concentration) did not reduce the number of HCT 116 cells that invaded [control: 254 ± 6; aprotinin: 246 ± 5 (mean ± SD)], indicating that the plasminogen cascade does not participate in invasion of these cells. Note that the presence of 1% serum as a source of plasminogen increased the number of cells that invaded through Matrigel. The inability of aprotinin to inhibit HCT 116 invasion was consistent with the inability of aprotinin to reduce degradation of DQ-collagen IV. Thus, the proteases responsible for degradation of DQ-collagen IV by the HCT 116 cells seem to be the same as those responsible for invasion through Matrigel as broad spectrum MMP inhibitors and cathepsin B-selective inhibitors block both processes.
Discussion
Degradation of basement membrane appears to be an essential step for tumor invasion, but not for migration of tumor cells (for recent reviews, see Refs. [33]–[35]). To evaluate degradation and invasion, we have developed a confocal microscopy assay in which living cells are grown in extracellular matrices mixed with quenched fluorescent protein substrates. This assay images proteolysis via the appearance of fluorescent cleavage products and in addition localizes the site at which those cleavage products appear [15]. Using this assay, we have determined that: (1) living human BT20 and BT549 breast carcinoma cell lines degrade DQ-collagen IV pericellularly or intracellularly, respectively, when grown in monolayer culture [15], and (2) living human U87 glioma cells degrade DQ-collagen IV intracellularly when grown in monolayer culture or as three-dimensional spheroids [15]. Individual glioma cells in the process of invading through the surrounding matrix also contain fluorescent degradation products of DQ-collagen intracellularly. We have established that the intracellular degradation takes place in lysosomes and requires endocytosis of the quenched fluorescent protein substrate [16]. The intracellular degradation is mediated in part by the lysosomal cysteine protease cathepsin B as it is abrogated by cathepsin B inhibitors, whereas the pericellular degradation is mediated by multiple classes of proteases as inhibitors of MMPs, the plasminogen cascade and cathepsin B all reduce pericellular proteolysis [15].

Degradation of DQ-collagen IV by cocultures of living human carcinoma spheroids and stromal fibroblasts is reduced by inhibitors of MMPs (C and D), the plasminogen cascade (E and F), or cathepsin B (G and H). The left panels represent cocultures of HCT 116 colon carcinoma and CCD-112CoN colon fibroblasts and the right panels of BT-20 breast carcinoma and 12TT breast fibroblasts: (A and B) control; (C and D) BB3103 (10 μM); (E and F) aprotinin (1 μM); and (G and H) CA074 and CA074Me (10 μM each). Single cell suspensions of tumor cells (alone or mixed with inhibitors) were plated on DQ-collagen IV/Matrigel-coated coverslips and incubated until attached (see Materials and Methods for details). Single cell suspensions of stromal and/or inflammatory cells were then added and the living cocultures imaged at 40–44 hr. Colon and breast fibroblasts were prelabeled with Cell Tracker Blue and Cell Tracker Orange so that they fluoresce blue and red, respectively. Fluorescence images were taken at an extended depth of focus and are superimposed on phase images of the living cocultures. Bar = 10 μm.
Here, we have extended these observations to include spheroids of living human HCT 116 and HKh-2 colon carcinoma cells and have determined that these spheroids degraded DQ-collagen IV both pericellularly and intracellularly. A reduction in DQ-collagen IV degradation in the presence of broad-spectrum inhibitors of MMPs and highly selective inhibitors of cathepsin B indicated that these proteases were involved in the pericellular degradation and that cathepsin B was the primary enzyme involved in the intracellular degradation. As in the BT549 breast carcinoma cells [15], the cleavage products in the colon carcinoma spheroids colocalized with active cathepsin B, suggesting that the intracellular degradation occurred in lysosomes. We have demonstrated that tumor cells of multiple origins are able to degrade type IV collagen intracellularly and, therefore, we propose that this is an important pathway for matrix degradation. Furthermore, the fact that spheroids of the invasive and malignant HCT 116 cell line exhibited more intracellular proteolysis than spheroids of the HKh-2 cell line would be consistent with intracellular proteolysis playing a role in invasion of colon carcinoma cells. That the two cell lines exhibited comparable pericellular proteolysis further substantiates an important role for intracellular proteolysis in invasion as does the presence of intracellular degradation products in living glioma cells in the process of invading through Matrigel [16]. The confocal microscopy assay used in all of these studies evaluates proteolysis by living cells, that is, cells that are capable of taking up substrates by endocytosis, and thereby has allowed us to demonstrate that invasion of tumor cells can involve intracellular proteolysis, as well as pericellular proteolysis.
Spheroids of living human BT20 breast carcinoma cells only degraded DQ-collagen IV pericellularly as we had previously observed for this cell line in monolayer culture [15]. Interestingly, individual BT20 cells in the process of invading through the surrounding matrix also degraded DQ-collagen pericellularly, a finding that contrasts with what we have observed for individual invading U87 glioma cells [16]. Our studies, thus, indicate that both pericellular and intracellular pathways of proteolysis contribute to tumor invasion and that which pathway(s) is used varies with the tumor cells.
There is a growing literature supporting a role for tumor-associated stromal and inflammatory cells in progression, angiogenesis, invasion, and metastasis of colon and breast cancers [36]–[45]. To our knowledge, the present study is the first to provide a direct demonstration that the interaction of colon and breast cancer cells with stromal fibroblasts dramatically increased proteolysis of the basement membrane protein type IV collagen. Monocytes, which did not degrade DQ-collagen IV themselves, were also ineffective at stimulating degradation in cocultures with tumor cells. Macrophages in the cocultures increased degradation, primarily due to the macrophages themselves degrading DQ-collagen IV intracellularly. Fibroblasts seemed to be more effective in increasing pericellular proteolysis, as well as intracellular proteolysis by the tumor cells. Of significance is that we were able to reduce the proteolysis of DQ-collagen by cocultures of colon carcinoma/colon fibroblasts with inhibitors of the plasminogen cascade, MMPs, and cathepsin B, yet, we were not able to reduce the proteolysis of DQ-collagen IV by colon carcinoma cells alone with an inhibitor of the plasminogen cascade. Urokinase has been reported to be present in stromal fibroblasts but absent in colon carcinoma cells [30]. The urokinase receptor is present on both cell types with the receptor on the surface of the colon tumor cells participating in activation of plasminogen by binding urokinase from the tumor-associated fibroblasts [30]. This is consistent with our observations that aprotinin, a plasmin inhibitor, reduced degradation of DQ-collagen IV by the cocultures, but not by the tumor cells alone. Another component of the plasminogen cascade is the urokinase receptor associated protein (uPARAP/Endo18), a protein that has been hypothesized to play a regulatory role in pericellular proteolysis [46]. uPARAP has recently been shown to play a central role in endocytosis of collagens including collagen IV [47], a process required for the intracellular degradation of DQ-collagen IV observed in our studies. uPARAP, thus, may provide a linkage between the pericellular and intracellular pathways of degradation and between the plasminogen cascade and cathepsin B.
The confocal microscopy assay described here was able to image proteolysis by living cells (tumor, stromal, and inflammatory) in three-dimensional cocultures and, therefore, provides an in vitro model in which one can study the ability of the various cell components of a solid tumor to degrade proteins and how those components interact to facilitate tumor proteolysis. The parallels between proteolysis in the present in vitro model and proteolysis in vivo suggest that we may be able to use this model to identify proteases that degrade specific extracellular matrix proteins in vivo. Ongoing studies in which the fibroblasts used in the cocultures come from mice deficient in specific proteases indicate that this should be possible and that this in vitro model may be useful for screening probes to image protease activity and protease inhibitors. For example, this in vitro model could be used to confirm the role of cysteine proteases using the protease-activated probes developed for imaging cysteine proteases by Bogyo et al. [48,49] or the roles of MMPs and cathepsin B using the near-infrared probes developed for in vivo imaging of protease activity by Weissleder et al. [8,50–52].
Footnotes
Acknowledgments
This work was supported by National Institutes of Health (NIH) Grant CA 56586 and a Breast Cancer Center of Excellence Award, DAMD17-02-1-0693. The Microscopy and Imaging Resources Laboratory is supported, in part, by NIH Center Grants P30ES06639 and P30CA22453. The Barbara Ann Karmanos Cancer Institute Comprehensive Cancer Center Cell Resources core was supported by NIH Center Grant P30CA22453.