Free accessResearch articleFirst published online 2010-3
Role of the oestrogen receptor (ESR1 PvuII and ESR1 325 C→G) and progesterone receptor (PROGINS) polymorphisms in genetic susceptibility to migraine in a North Indian population
We aimed to explore the single-locus, haplotype and epistasis patterns and the contribution of oestrogen receptor [ESR1 PvuII (rs2234693), ESR1 325 C→G (rs1801132)] and progesterone receptor [PROGINS (rs1042838)] polymorphisms in genetic susceptibility to migraine by analysing 613 subjects consisting of 217 migraine patients, 217 healthy controls (HC) and 179 patients with tension-type headache (TTH). Entire data were analysed by taking the Bonferroni corrected P-value into account. We found significant association of TT genotype [odds ratio (OR) 3.458, confidence interval (CI) 1.757, 6.806; P = 0.0003] and T allele (OR 1.729, CI 1.309, 2.284; P = 0.0001) of ESR1 PvuII single nucleotide polymorphism with migraine when compared with HC. Significant association was seen only in female migraine patients at both genotype (P = 0.002; OR 3.834, CI 1.625, 9.043) and allele level (P = 0.002; OR 1.721, CI 1.228, 2.413). Moreover, higher risk was limited to migraine with aura (MA) (in case of TT genotype, P = 0.002; OR 4.377, CI 1.703, 1.246; in case of T allele, P = 0.001; OR 1.888, CI 1.305, 2.735) rather than migraine without aura (MoA) (P-value of TT genotype = 0.003; OR 3.082, CI 1.465, 6.483; P-value T allele = 0.002; OR 1.630, CI 1.188, 2.236). In case of a recessive model, risk was seen with migraine patients (P = 0.0003; OR 2.514, CI 1.635, 3.867), MA (P = 0.0001; OR 3.583, CI 1.858, 6.909) and MoA patients (P = 0.002; OR 2.125, CI 1.304, 3.464) when compared with HC. No risk was observed when TTH patients were compared with HC. No significance of ESR 325 G→C polymorphism was seen in any of the models under study. Significant differences in genotypic (P = 0.0001) and allelic frequency (P = 0.0002) were seen in case of PROGINS polymorphism when migraine patients were compared with HC, showing a protective effect (for A1A2 genotype, OR 0.292, CI 0.155, 0.549; for A2 allele, OR 0.320, CI 0.174, 0.589). Moreover, significance was seen only in case of female migraine patients at genotype (P = 0.002; OR 0.344, CI 0.176, 0.684) as well as allele levels (P = 0.004; OR 0.379, CI 0.198, 0.727) in case of PROGINS polymorphism. ESR1 PvuII TT*ESR1 325 C→G CG genotype, PROGINS A1A2*ESR1 325 C→G CG genotype and ESR1 PvuII CT*PROGINS A1A2 interacted significantly, but significance was lost after Bonferroni correction. In conclusion, ESR1 PvuII polymorphism is a significant risk factor for migraine particularly in women and MA patients, but ESR 325 C→G polymorphism is not associated with migraine susceptibility. PROGINS polymorphism seems to play a protective role in genetic susceptibility to migraine in the North Indian population.
Migraine is one of the complex polygenic, multifactorial genetic disorders, and the interplay of vascular, hormonal, neurotransmitter receptor genes and environment accounts for susceptibility to this disorder.
The hormonal link between female sex and migraine can be accounted for by the fact that migraine develops in three times as many women as men during the adult years, although no gender difference in migraine occurrence has been found prior to puberty (1). In most women, migraine worsens around the time of menstruation, and declines after menopause or during pregnancy (2). It has been seen that both oestrogen withdrawal and high oestrogen concentrations increase migraine risk in women (3). The classic intracellular oestrogen receptors (ERα and ERβ) are widely expressed throughout the central nervous system (4) and are known to be localized in those regions of the brain that are considered to be involved in migraine pathogenesis. The progesterone receptors (PRs) are found in various regions of the human brain including serotonin neurons (5). Hence, it is possible that alteration in ERs and PRs plays a role in migraine susceptibility (6,7).
The human ESR1 gene coding ERs is located on chromosome 6q25.1 and contains eight exons (8). ESR1 PvuII polymorphism, also known as c.454-397T/C, IVS1-397 T/C (9), is involved in fluctuations in oestradiol levels (10). Another polymorphism, ESR1 325 C→G single nucleotide polymorphism (SNP), is located in hormonal binding region of exon 4 (11). The variant allele of ESR1 325 C→G polymorphism has been shown to confer risk of migraine (12, 13), but the risk was not significant after corrections for multiple testing (13). However, Colson et al. (11) did not find any association of ESR1 325 C→G or ESR1 PvuII polymorphisms in migraine susceptibility. The human PR (PROGINS) gene is located on chromosome 11q22. A 306-bp polymorphic Alu insertion is located in intron 7 of the PR gene (14). In a White population, it has been reported that patients carrying the PROGINS insert were 1.8 times more likely to suffer form migraine (7).
There are conflicting reports on the role genetic variants of these ESR1 genes play in migraine susceptibility. Also, there is only one study regarding the association of PROGINS polymorphism with risk of migraine. We thus planned our study to determine the role of these hormonal gene variations (ESR1 PvuII, ESR1 325 C→G and PROGINS) in migraine susceptibility in a Northern Indian population. Since non-migraine headaches could also transform into migraine at later stages (15), we compared the association of hormonal gene variants with healthy controls as well as with a group consisting of patients with tension-type headache (TTH).
Subjects and methods
Subjects
The study was ethically approved by the Institutional Committee. Patients were recruited from the Neurology Out-patient Clinic of Sanjay Gandhi Post Graduate Institute of Medical Sciences (Lucknow, India). Patients were diagnosed according to the questionnaire prepared by an experienced neurologist, based on the criteria of the International Headache Society (16). We excluded migraine patients with uncontrolled hypertension (blood pressure > 160/95), peripheral vascular syndromes (such as ischaemic bowel disease), history, symptoms or risk of ischaemic heart disease or any risk for cardiovascular disease or patients with thyroid disorders. Migraine patients (n = 217), 84 having migraine with aura (MA) and 133 migraine without aura (MoA) and 179 normotensive TTH patients were recruited. The normal control group (HC) comprised 217 age- and sex-matched healthy and normotensive volunteers free of any routine headache, neurological or vascular diseases. We also excluded volunteers who had any sort of cardiovascular disorder or diabetes. The ethnic and socioeconomic profile of healthy controls was similar to that of patient groups. The HC were recruited after taking into consideration the mean age of the selected patients. Data are presented as mean ± standard deviation (s.d.) (Table 1). The HC originated from the same geographical region (North India), were recruited in parallel, and at a similar time and geographical location.
Demographic profile of the subjects under study
Subjects
Number of subjects
Mean age ± s.d.
Total migraine
217
32.04 ± 12.24
MA
84
31.57 ± 11.85
MoA
133
32.32 ± 12.51
TTH
179
32.36 ± 15.02
HC
217
32.62 ± 12.26
Male
Total migraine
67
29.17 ± 15.72
MA
21
29.35 ± 16.46
MoA
46
29.09 ± 15.57
TTH
56
31.41 ± 16.00
HC
67
30.48 ± 10.95
Female
Total migraine
150
33.31 ± 10.14
MA
63
32.29 ± 10.00
MoA
87
34.03 ± 10.24
TTH
123
32.80 ± 14.60
HC
150
33.65 ± 12.75
HC, healthy controls; MA, migraine with aura; MoA, migraine without aura; s.d., standard deviation; TTH, tension-type headache.
Genotyping
The genomic DNA was extracted from a peripheral blood leucocyte pellet of patients' blood using the standard salting out method (17). Quantification of DNA was done by nanodrop spectrophotometer. The study was performed blinded to disease status, as samples were taken randomly for genetic analysis. Analysis for ESR1 PvuII and ESR1 325 C→G gene polymorphisms was performed by polymerase chain reaction (PCR)-restriction fragment length polymorphism method as described by Lai et al. (18) and Iwase et al. (8), respectively. Analysis of PROGINS was based on the PCR amplification of a fragment encompassing the 306-bp insertion polymorphism in intron 7 (19) as described by Rowe et al. (14). To improve the quality of genotyping and its validity, 20% of samples were re-genotyped by another laboratory member and no discrepancy in genotyping was found. Genotyping of 5% of these samples was reconfirmed through sequencing.
Statistical analysis
Sample size and power was calculated using software quanto Version 1.0 (http://hydra.usc.edu/gxe) for each genetic marker. The significance level of our study was set at 0.05 in a two-sided test. In the inheritance model, we chose the log-additive one, which is the most suitable model in polygenic diseases. Migraine disease prevalence of 12% was assumed. Setting the threshold for significance at 0.05 and assuming a minor allele frequency of 33.9, 37.3 and 9.7% (in case of ESR1 PvuII, ESR C325G and PROGINS, respectively) and a relative risk of 1.85, our study has power > 80 (99.0 for ESR PvuII, 99.18 for ESR C325G and 82 for PROGINS) to detect a SNP in linkage disequilibrium (LD) with the disease causing variant. Goodness-of-fit χ2 test was used to check if genotypes were in Hardy–Weinberg equilibrium.
All other analyses were done using spss version 15 (SPSS, Chicago, IL, USA). The entire analysis was age and sex adjusted. χ2 test was applied for the analysis of genotypic and allelic distributions. Logistic regression analysis was used to determine the contribution of genetic polymorphisms to the risk of disease. Two-tailed tests of significance were used throughout. The relative risk for different genotypes was estimated calculating the odds ratio (OR) and 95% confidence interval (CI). P < 0.05 was considered statistically significant. The levels of P-values were adjusted by Bonferroni correction to account for multiple testing issues. Since we have genotyped three polymorphisms in one sample and have also performed several comparisons within this sample per polymorphism (four subgroups like MA vs. MoA and male/female), Bonferroni correction was applied for multiple comparisons with all novel associations, with a correction factor derived from the number of SNPs and subgroups or number of haplotypes tested (P-value after Bonferroni correction = Pcorr), to avoid chances of type 1 error. The coefficient of Bonferroni correction (Pcorr) in our analysis in case of recessive, dominant and genotype model as well as at allelic level was taken as 12 (4 subgroups × 3 polymorphisms). So, the Bonferroni-corrected P-value, Pcorr = 0.05/12 = 0.004 in case of analysis of the genotypic data of migraine patients. Pcorr in case of analysis of TTH is = 0.05/6 = 0.008 [2 subgroups (male/female) × 3 polymorphisms]. In case of haplotype comparisons the Bonferroni-corrected P-value = 0.05/4 = 0.0125 (no. of haplotypes formed = 4). In case of interaction analysis of the three genes Pcorr = 0.05/9 = 0.005 (3 SNPs × 3 comparisons). For the purpose of analysis, we have taken the respective Bonferroni-corrected P-value (Pcorr) as the significant value of our study to account for multiple testing issues. Allele and genotype frequencies were computed for cases and controls for all three polymorphisms separately. We performed the genotype- and allele-specific analysis to examine the main effect of the variant genotype as well as the variant allele on risk of migraine, by fitting dominant and recessive models. Gene–gene interaction for the three polymorphisms was also performed using logistic regression analysis to examine the possibility of combined effects of the three polymorphisms in migraine susceptibility. Association was expressed as ORs or as risk estimates with 95% CIs. Corrected P-value after Bonferroni correction, i.e. Pcorr < 0.0005, was considered statistically significant.
Haplotype frequencies were calculated for the SNPs using SNP analyser V 1.2. LD results are presented as r2 and P-values, where r2 is the square of the correlation coefficient for a given marker pair and is a measure of LD between two genetic markers (for perfect LD, r2 = 1).
Results
The genotypic and allelic frequencies of subjects in the studied polymorphisms are illustrated in Table 2. The genotypic frequencies of our control population followed Hardy–Weinberg equilibrium for all three studied polymorphisms.
ESR1 PvuII (rs2234693), ESR1 325 C→G (rs1801132) and PROGINS (rs1042838) polymorphism genotype frequency distribution in studied subjects
ESR PvuII
ESR 325 C→G
PROGINS
Genotype distribution N (%)
Allele distribution N# (%)
Genotype distribution N (%)
Allele distribution N# (%)
Genotype distribution N (%)
Allele distribution N# (%)
Subjects
N
CC
CT
TT
C
T
CC
CG
GG
C
G
A1A1
A1A2
A2A2
A1
A2
Total
Migraine
217
47 (21.7)
136 (62.7)
34 (15.7)
230 (53.0)
204 (47.0)
75 (34.6)
115 (53.0)
27 (12.4)
265 (61.1)
169 (38.9)
201 (92.6)
16 (7.4)
0 (0.0)
418 (96.3)
16 (3.7)
MA
84
14 (16.7)
58 (69.0)
12 (14.3)
86 (51.2)
82 (48.8)
32 (38.1)
42 (50.0)
10 (11.9)
106 (63.1)
62 (36.9)
67 (91.8)
7 (8.2)
0 (0.0)
154 (91.7)
14 (8.3)
MoA
133
33 (24.8)
78 (58.6)
22 (16.5)
144 (54.1)
122 (45.9)
43 (32.3)
73 (54.9)
17 (12.8)
159 (59.8)
107 (40.8)
124 (93.2)
9 (6.8)
0 (0.0)
248 (93.2)
18 (6.8)
TTH
179
57 (31.8)
95 (53.1)
27 (15.1)
209 (58.4)
149 (41.6)
53 (29.6)
102 (57.0)
24 (13.4)
208 (58.1)
150 (41.9)
163 (91.1)
16 (8.9)
0 (0.0)
342 (95.5)
16 (4.5)
HC
217
88 (40.6)
111 (51.2)
18 (8.3)
287 (66.1)
147 (33.9)
81 (37.3)
110 (50.7)
26 (12.0)
272 (62.7)
162 (37.3)
175 (80.6)
42 (19.4)
0 (0.0)
392 (90.3)
42 (9.7)
Male
Migraine
67
13 (19.4)
43 (64.2)
11 (16.4)
69 (51.5)
65 (48.5)
20 (29.9)
40 (59.7)
7 (10.4)
80 (59.7)
54 (40.3)
66 (98.5)
1 (1.5)
0 (0.0)
133 (99.3)
1 (0.7)
MA
21
4 (19.0)
13 (61.9)
4 (19.0)
21 (50.0)
21 (50.0)
6 (28.6)
12 (57.1)
3 (14.3)
24 (57.1)
18 (42.9)
21 (100.0)
0 (0.0)
0 (0.0)
42 (100.0)
0 (0.0)
MoA
46
9 (19.6)
30 (65.2)
7 (15.2)
48 (52.2)
44 (47.8)
14 (30.4)
28 (60.9)
4 (8.7)
56 (60.9)
36 (39.1)
45 (97.8)
1 (2.2)
0 (0.0)
90 (97.8)
2 (2.2)
TTH
56
17 (30.4)
29 (51.8)
10 (17.9)
63 (56.3)
49 (43.8)
18 (32.1)
29 (51.8)
9 (16.1)
65 (58.0)
47 (42.0)
53 (94.6)
3 (5.4)
0 (0.0)
109 (97.3)
3 (2.7)
HC
67
27 (40.3)
32 (47.8)
8 (11.9)
86 (64.2)
48 (35.8)
28 (41.8)
31 (46.3)
8 (11.9)
87 (64.9)
47 (35.1)
58 (86.6)
9 (13.4)
0 (0.0)
125 (93.3)
9 (6.7)
Female
Migraine
150
34 (36.7)
93 (50.0)
23 (13.3)
161 (53.7)
139 (46.3)
55 (36.7)
75 (50.0)
20 (13.3)
185 (61.7)
115 (38.3)
135 (90.0)
15 (10.0)
0 (0.0)
285 (95.0)
15 (5.0)
MA
63
10 (15.9)
45 (71.4)
8 (12.7)
65 (51.6)
61 (58.4)
26 (41.3)
30 (47.6)
7 (11.6)
82 (65.1)
44 (34.9)
56 (88.9)
7 (11.1)
0 (0.0)
112 (88.9)
14 (11.1)
MoA
87
24 (27.6)
48 (55.2)
15 (17.2)
96 (55.2)
78 (44.8)
29 (33.3)
45 (51.7)
13 (14.9)
103 (59.2)
71 (40.8)
79 (90.8)
8 (9.2)
0 (0.0)
158 (90.8)
16 (9.2)
TTH
123
40 (32.5)
66 (53.7)
17 (13.8)
146 (59.3)
100 (40.7)
35 (28.5)
73 (59.3)
15 (12.2)
143 (58.1)
103 (41.9)
110 (89.4)
13 (10.6)
0 (0.0)
233 (94.7)
13 (5.3)
HC
150
61 (40.7)
79 (52.7)
10 (6.7)
201 (67.0)
99 (33.0)
53 (35.3)
79 (52.7)
18 (12.0)
185 (61.7)
115 (38.3)
117 (78.0)
33 (22.0)
0 (0.0)
267 (89.0)
33 (11.0)
A1, wild-type allele of PROGINS; A2, variant allele of PROGINS; C, wild-type allele of ESR1 (PvuII and 325C > G); ESR1, oestrogen receptor; G, variant allele of ESR 325C→G; HC, healthy controls; MA, migraine with aura; MoA, migraine without aura; N, total number of subjects; N#, total chromosome number; PROGINS, progesterone receptor; TTH, tension-type headache; T, variant allele of ESR1 PvuII.
ESR1 PvuII polymorphism
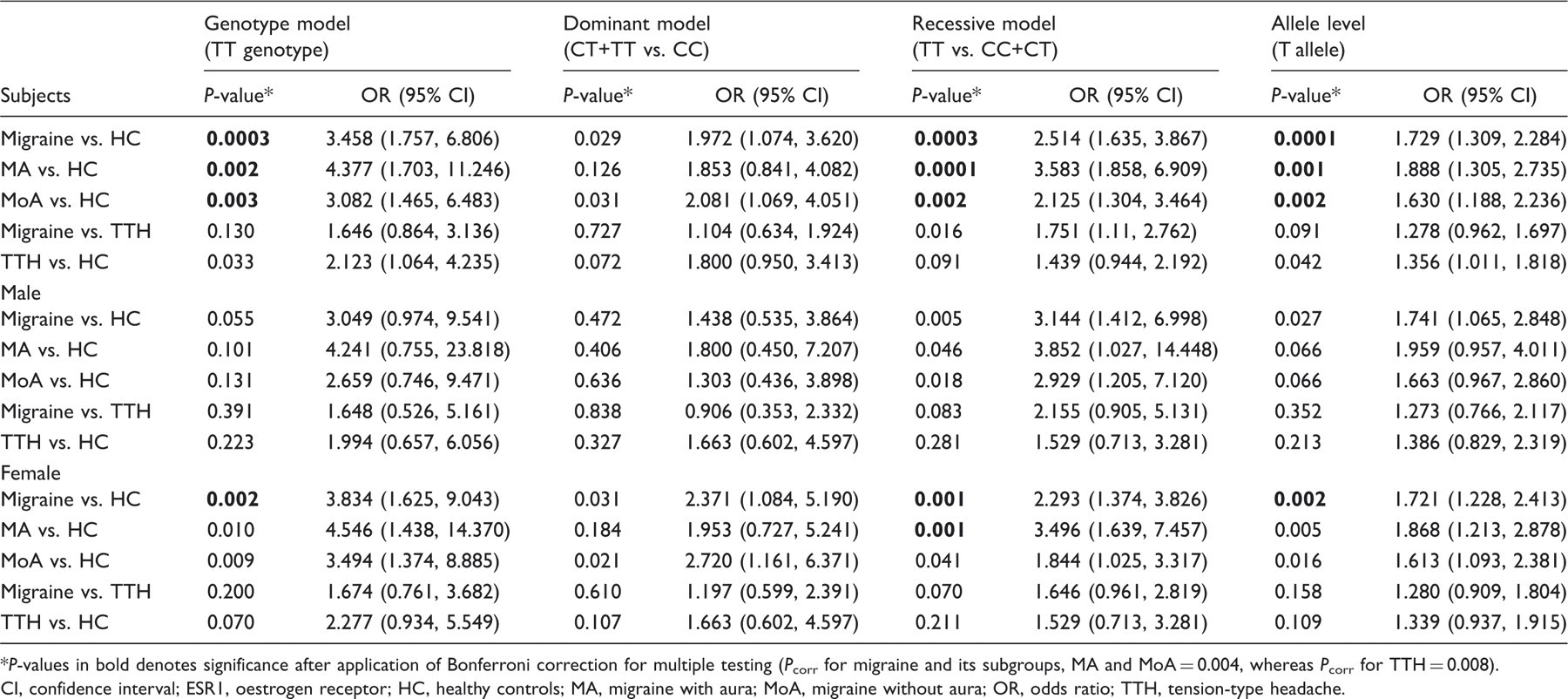
Since we have taken the Bonferroni corrected P-value (Pcorr) at genotype and allele level to be 0.004, we observed significant differences in the frequency of TT genotype when migraine patients were compared with HC (P = 0.0003; OR 3.458, CI 1.757, 6.806) and not with TTH. A similar trend was observed at allele level also (migraine vs. HC, P = 0.0001; OR 1.729, CI 1.309, 2.284). When migraine patients were segregated on gender basis, significance differences were observed only in case of female migraine patients at genotype (P = 0.002; OR 3.834, CI 1.625, 9.043) as well as allele level (P = 0.002; OR 1.721, CI 1.228, 2.413) when compared with HC. On subgrouping total migraine patients into MA and MoA, higher risk of TT genotype (P = 0.002; OR 4.377, CI 1.703, 11.246) and T allele (P = 0.001; OR 1.888, CI 1.305, 2.735) was limited to MA. After Bonferroni correction, we found an association of T allele in recessive model, even after subgrouping the patients into MA (P = 0.0001) and MoA (P = 0.002), while the significance was lost in case of dominant model. Within the MA and MoA subgroups, statistical significance after Bonferroni correction was seen only in case of female MA patients in recessive model (P = 0.001) (Table 3). No significant difference at genotype or allele level or in case of dominant and recessive models was observed on comparing TTH with HC. With the various clinical parameters of patients with migraine, the association was lost after Bonferroni correction (data not shown).
Comparison of P-value and odds ratio associated with variant ESR1 PvuII (rs2234693) genotype (TT), and T allele frequencies in study subjects*
P-values in bold denotes significance after application of Bonferroni correction for multiple testing (Pcorr for migraine and its subgroups, MA and MoA = 0.004, whereas Pcorr for TTH = 0.008).
No significant differences in genotypic and allelic frequencies was found in ESR1 325 C→G polymorphism in case of migraine when compared with HC or TTH.
Haplotype analysis
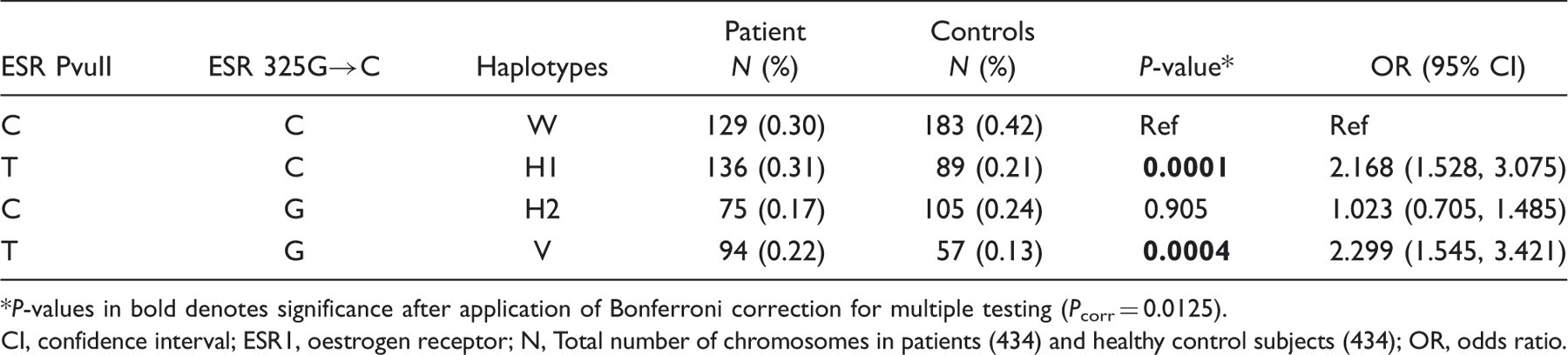
ESR1 PvuII and ESR1 325 C→G polymorphisms were not in LD, in case of either migraine patients (r2 = 0.0021; P = 0.3919 after Yate's continuity correction) or HC (r2 = 0.0010; P = 0.5844). Four different haplotypes (W, H1, H2, V) were possible (Table 4). The frequency of haplotype formed by the heterozygous variant minor alleles ESR1 PvuII-*T- ESR1 325 C→G*G (V) varied significantly (even after Bonferroni correction) between patients and controls, showing risk (P = 0.0004; OR 2.299, CI 1.545, 3.421). Similarly, ESR1 PvuII-*T- ESR1 325 C→G*C showed risk (H1) (P = 0.0001; OR 2.168, CI 1.528, 3.075) when compared with HC, and the risk could be due to the individual effect T allele of ESR PvuII polymorphism (Table 4).
Haplotype analysis results between ESR1 PvuII* ESR1 325 C→G (migraine patients vs. HC)
P-values in bold denotes significance after application of Bonferroni correction for multiple testing (Pcorr = 0.0125).
CI, confidence interval; ESR1, oestrogen receptor; N, Total number of chromosomes in patients (434) and healthy control subjects (434); OR, odds ratio.
PROGINS polymorphism
Taking Pcorr value after Bonferroni correction to be 0.004, a significant difference in genotypic (P = 0.0001) and allelic frequency (P = 0.0002) was seen in PROGINS polymorphism when migraine patients were compared with HC, showing protection (for A1A2 genotype, OR 0.292, CI 0.155, 0.549; for A2 allele, OR 0.320, CI 0.174, 0.589). Moreover, significance was seen only in case of female migraine patients at genotype (P = 0.002; OR 0.344, CI 0.176, 0.684) as well as allele level (P = 0.004; OR 0.379, CI 0.198, 0.727). Similarly, significance was observed only in case of MoA (P = 0.001; OR 0.290, CI 0.135, 0.621) when compared with HC. When TTH patients were compared with HC, significance was obtained showing protection also at allele level. No significance was obtained on subgrouping TTH group on gender basis (Table 5) or on comparison with other clinical variables (data not shown).
P-value and odds ratio in ESR1 325 C→G (rs1801132) and PROGINS (rs1042838) in migraine patients, non-migraine headache patients and healthy controls
P-values in bold denotes significance after application of Bonferroni correction for multiple testing (Pcorr for migraine and its subgroups, MA and MoA = 0.004, whereas Pcorr for TTH = 0.008).
CI, confidence interval; ESR1, oestrogen receptor; HC, healthy controls; MA, migraine with aura; MoA, migraine without aura; OR, odds ratio; PROGINS, progesterone receptor; TTH, tension-type headache; ϕ, for GG genotype; $, for A1A2 genotype.
For PROGINS at allele level (A2 allele):
P value = 0. 0002; OR = 0.320; CI = 0.174, 0.589.
P value = 0.033; OR = 0.320; CI = 0.174, 0.589.
P value = 0.004; OR = 0.379; CI = 0.198, 0.727.
P value = 0.003; OR = 0.320; CI = 0.174, 0.589.
P value = 0.008; OR = 0.320; CI = 0.174, 0.589.
P value = 0.008; OR = 0.320; CI = 0.174, 0.589.
P value = 0.028; OR = 0.353 CI = 0.139, 0.895.
P value = 0.001; OR = 0.353; CI = 0.139, 0.895.
P value = 0.010; OR = 0.353; CI = 0.139, 0.895.
Gene–gene interaction
The ESR1 PvuII TT*ESR1 325 C→G CG genotype (P = 0.047; OR 2.273, 95% CI 1.011, 5.113), PROGINS A1A2*ESR1 325 C→G CG genotype (P = 0.049; OR 0.121, CI 0.015, 0.994) and ESR1 PvuII CT*PROGINS A1A2 (P = 0.024; OR 0.373, CI 0.159, 0.877) interacted significantly in conferring migraine susceptibility, but the significance was lost after taking the Bonferroni corrected P-value (Pcorr = 0.005) into account (data not shown).
Discussion
Ours is the first report on genetic polymorphism from North India that deals with the role of hormonal genes in migraine susceptibility. The objective of this study was to determine the influence of the ESR1 (PvuII and 325 C→G) and PROGINS (insertion) polymorphism in independent involvement in migraine susceptibility, and also their interactive influence, by comparing migraine patients with patients having headache other than migraine (TTH) and healthy controls.
The variant allele frequencies of ESR1 PvuII, ESR1 325 C→G and PROGINS insertion polymorphism in our control population were 33.9, 37.3 and 9.7%, respectively, which is similar to those reported in other Indian populations (19–21).
We found the ESR1 PvuII polymorphism to be associated with migraine. Significant risk at genotype, allele as well as haplotype interface was observed in migraine as well as MA and MoA when compared with HC. Moreover, at genotype and allele level, the risk was confined to female patients. No association was seen when TTH patients were compared with HC. To define further the role of the polymorphism, we evaluated the association in terms of dominant and recessive models. When migraine, MA as well as MoA patients were compared with HC, significant association was seen only in case of recessive models, which shows that the variant T allele showed risk only when present in two copies (as in the recessive model) and not when present in one copy (as in the dominant model). Similarly, female MA patients showed risk only in case of recessive model when compared with HC. The published reports from other studies have been summarized in Table 6, showing population-specific differences between the studied populations.
Other published studies on migraine (migraine vs. HC) in ESR1 PvuII, ESR1 325 C→G and PROGINS polymorphism
(275 Migraineurs and 275 unrelated control individuals)
(MA+MoA)
0.004
χ2 = 6.50
Male (MA+MoA)
0.82
χ2 = 0.41
Female (MA+MoA)
0.005
χ2 = 10.64
MA
0.33
χ2 = 2.25
MoA
0.001
χ2 = 13.08
Population 2
(Independent follow-up population of 300 Migraineurs and 300 controls)
(MA+MoA)
0.019
χ2 = 7.92
Male (MA+MoA)
0.07
χ2 = 5.27
Female (MA+MoA)
0.09
χ2 = 4.81
MA
0.17
χ2 = 3.53
MoA
0.026
χ2 = 7.28
P-value remained insignificant after Bonferroni correction.
χ2, chi square value; OR, odds ratio; CI, confidence Interval; ESR1, oestrogen receptor; MA, migraine with aura; MoA, migraine without aura; PROGINS, progesterone receptor.
We failed to find any association of the ESR1 325 C→G polymorphism with migraine, which is consistent with the finding of Colson et al. (11) (Table 6). Moreover, ESR1 325 C→G polymorphism is a synonymous polymorphism with no associated amino acid change (P325P) (13), consequently it is unlikely that this polymorphism is causative. A Finnish study (13) showing no association of ESR1 325 C→G polymorphism with migraine after Bonferroni correction is also summarized in Table 6. Although Oterino et al. (12) found that ESR1 325 C→G polymorphism conferred risk for migraine in their Spanish population, they also stated that it is not really a causative polymorphism for migraine. Instead, they postulated that an unknown genetic variation between the two studied SNPs (ESR1 325 C→G and ESR1 594 G→A) could be responsible for the association. The absence of LD between ESR1 C325G and ESR1 G594A polymorphisms (D' = 0.004; P = NS) (12) and ESR PvuII and ESR1 G594A (D' = 0.016; P = NS) in the study by Colson et al. (11) also indicated that both polymorphisms belong to a distinct LD block. No association of ESR1 325 C→G polymorphism with patients with headache other than migraine (TTH) was seen in our studied population from North India.
The PROGINS insertion polymorphism is considered to have a deleterious effect on PR expression, through recombination or mis-splicing (14,22). However, we found an association of variant allele in case of migraine, female migraine as well as MoA patients when compared with HC, showing protection in case of PROGINS polymorphism. A study on breast cancer has also reported that the protective role of variant A2 allele of PROGINS gene is confined to premenopausal women (23). It has also been reported that the presence of oestrogens may be required for the expression of progesterone in parts of the brain, which then can act in a synergistic, antagonistic or neutral manner compared with the effects of oestrogens (24). However, Colson et al. (7) observed significant risk of PROGINS insertion polymorphism in case of migraine patients (Table 6). Our results also suggest significant population-specific differences of PROGINS polymorphism in migraine susceptibility.
What is the possible mechanism of ESR1 PvuII polymorphism in migraine pathogenesis? Being in the intronic region, the ESR1 PvuII polymorphism has no known function, but a study has shown that the PvuII polymorphism may affect the splicing of ERα mRNA, thus resulting in the alteration of protein expression (25). Its position near the gene promoter might suggest its role in either transcription regulation or mRNA processing and stability (26). As reported earlier also (27), it may be in LD with another unknown polymorphism that may be functional. A change of ESR1 gene expression by altering the binding of transcription factors and their influence on alternative splicing of ESR1 gene could be the possible mechanism of action of ESR1 PvuII polymorphism. Also, involvement of ESR1 PvuII polymorphism in variation of oestradiol levels in postmenopausal women (10) has been suggested. Moreover, we found that in case of ESR1 PvuII polymorphism, the risk was more in MA than in MoA. Previous studies also confirm the role of incidence of MA and oestrogen (3).
So, we can infer from our genetic analysis that the intronic region polymorphism, ESR1 PvuII, plays a role in migraine susceptibility in North Indian patients, but the exonic region polymorphism of ER, the ESR1 325 C→G polymorphism, does not seem to influence genetic predisposition to migraine headache. Moreover, the PROGINS polymorphism seems to have a protective effect on the genetic susceptibility of migraine. The effect of ESR1 PvuII was seen predominantly in women, both at genotype and allele level. The variant T allele appears to influence susceptibility in a recessive manner. The inconsistency of our results with other published studies suggest either true variability in ethnic groups or differences in modifying genetic or environmental factors.
Footnotes
Acknowledgment
We acknowledge the Indian Council of Medical Research, New Delhi, India for financial assistance in our study. We are very thankful to Dr Hardeep Malhotra and Dr Vibhor Pardarshini, Assistant Professor, IHBAS, New Delhi for their clinical input in our study. The Indian Council of Medical Research, New Delhi, India, supported this work.
References
1.
SilbersteinSDMerriamGR. Physiology of the menstrual cycle. Cephalalgia2000; 20: 148–54.
2.
MaggioniFAlessiCMagginoTZanchinG. Headache during pregnancy. Cephalalgia1997; 17: 765–69.
3.
SomervilleBW. The role of progesterone in menstrual migraine. Neurology1971; 21: 853–59.
4.
MartinVTBehbehaniM. Ovarian hormones and migraine headache: understanding mechanisms and pathogenesis—part I. Headache2006; 46: 3–23.
5.
BetheaCLLuNZGundlahCStreicherJM. Diverse actions of ovarian steroids in the serotonin neural system. Front Neuroendocrinol2002; 23: 41–100.
6.
ColsonNJLeaRAQuinlanSMacMillanJGriffithsLR. The estrogen receptor 1 G594A polymorphism is associated with migraine susceptibility in two independent case/control groups. Neurogenetics2004; 5: 129–33.
7.
ColsonNJLeaRAQuinlanSMacMillanJGriffithsLR. Investigation of hormone receptor genes in migraine. Neurogenetics2005; 6: 17–23.
8.
IwaseHGreenmanJMBarnesDMHodgsonSBobrowLMathewCG. Sequence variants of the estrogen receptor (ER) gene found in breast cancer patients with ER negative and progesterone receptor positive tumors. Cancer Lett1996; 108: 179–84.
9.
Van DuijnhovenFJBezemerIDPeetersPHRoestMUitterlindenAGGrobbeeDE. Polymorphisms in the estrogen receptor alpha gene and mammographic density. Cancer Epidemiol Biomarkers Prev2005; 14: 2655–60.
10.
SchuitSCde JongFHStolkLKoekWNvan MeursJBSchoofsMW. Estrogen receptor alpha gene polymorphisms are associated with estradiol levels in postmenopausal women. Eur J Endocrinol2005; 153: 327–34.
11.
ColsonNJLeaRAQuinlanSGriffithsLR. No role for estrogen receptor 1 gene intron 1 Pvu II and exon 4 C325G polymorphisms in migraine susceptibility. BMC Med Genet2006; 28: 7–12.
12.
OterinoAPascualJRuiz de AlegríaCValleNCastilloJBravoY. Association of migraine and ESR1 G325C polymorphism. Neuroreport2006; 23: 61–4.
13.
KaunistoMAKallelaMHämäläinenEKilpikariRHavankaHHarnoH. Testing of variants of the MTHFR and ESR1 genes in 1798 Finnish individuals fails to confirm the association with migraine with aura. Cephalalgia2006; 26: 1462–72.
14.
RoweSMCoughlanSJMcKennaNJGarrettEKiebackDGCarneyDN. Ovarian carcinoma-associated TaqI restriction fragment length polymorphism in intron G of the progesterone receptor gene is due to an Alu sequence insertion. Cancer Res1995; 55: 2743–45.
15.
LiptonRBCadyRKStewartWFWilksKHallC. Diagnostic lessons from the spectrum study. Neurology2002; 58: S2731–S2731.
MillerSADykesDDPoleskyHF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucl Acids Res1988; 16: 1215–1215.
18.
LaiICLiaoDLBaiYMLinCCYuSCChenJY. Association study of the estrogen receptor polymorphisms with tardive dyskinesia in schizophrenia. Neuropsychobiology2002; 46: 173–5.
19.
OnsoryKSobtiRCAl-BadranAIWatanabeMShiraishiTKrishanA. Hormone receptor-related gene polymorphisms and prostate cancer risk in North Indian population. Mol Cell Biochem2008; 314: 25–35.
20.
MitraSDesaiMKhatkhatayMI. Association of estrogen receptor alpha gene polymorphisms with bone mineral density in postmenopausal Indian women. Mol Genet Metab2006; 87: 80–7.
21.
GovindanSAhmadSNVedicherlaBKodatiVJahanPRaoKP. Association of progesterone receptor gene polymorphism (PROGINS) with endometriosis, uterine fibroids and breast cancer. Cancer Biomark2007; 3: 73–8.
22.
DonaldsonCJCrapanzanoJPWatsonJCLevineEABatzerMA. PROGINS Alu insertion and human genomic diversity. Mutat Res2002; 501: 137–41.
23.
GohrkeSWClaudeJCBecherHKiebackDGRunnebaumIB. Progesterone receptor gene polymorphism is associated with decreased risk for breast cancer by age 50. Cancer Res2000; 60: 2348–50.
24.
DluzenDERamírezVD. Progesterone enhances L-dopa-stimulated dopamine release from the caudate nucleus of freely behaving ovariectomized-estrogen-primed rats. Brain Res1989; 494: 122–8.
25.
HuangQWangTHLuWSMuPWYangYFLiangWW. Estrogen receptor alpha gene polymorphism associated with type 2 diabetes mellitus and the serum lipid concentration in Chinese women in Guangzhou. Chin Med J2006; 119: 1794–801.
26.
JakimiukANowickaMBogusiewiczMAdamiakASkorupskiPMiotlaP. Prevalence of estrogen receptor alpha PvuII and XbaI polymorphism in population of Polish postmenopausal women. Folia Histochem Cytobiol2007; 45: 331–8.
27.
LowYLDunningAMDowsettMFolkerdEDoodyDTaylorJ. Phytoestrogen exposure is associated with circulating sex hormone levels in postmenopausal women and interact with ESR1 and NR1I2 gene variants. Cancer Epidemiol Biomarkers Prev2007; 6: 1009–16.