
Abstract
Cognitive processing was investigated interictally in 18 children with migraine without aura and 18 age-matched controls by measuring event-related potentials (ERPs) and reaction times (RTs) during an acoustic oddball paradigm. Results showed that N100 amplitude evoked by frequent stimuli was significantly smaller in patients compared with controls. Habituation of target P300 amplitude was observed in patients but not in controls. Mean RTs were equivalent in the two groups, but migraine children made more errors than controls.
Introduction
Visual and auditory evoked potentials have been largely investigated in adult and juvenile migraine patients. Several studies report that, in the interictal period, increased amplitude and lack of habituation (i.e. amplitude decrease), or even potentiation, of visual evoked potentials (VEPs) N1-P1 component characterize migraineurs (especially with aura) compared with controls (1–4). With repeated presentation of the same auditory stimulus at increasing intensities, the auditory evoked potentials (AEPs) of migraineurs show a steeper amplitude/intensity function slope compared with healthy controls. The stronger intensity dependence of AEPs N1-P2 component between attacks has been repeatedly observed in both adults and children with migraine (5–7).
Overall, these dysfunctions are thought to reflect functional cortical hyperexcitability, possibly depending on abnormal central aminergic transmission (8). The migraine attack could be the final result of the failure of cortical mechanisms protecting against sensory overstimulation (9–11).
The recording of VEPs and AEPs provides a tool to assess sensory processing but does not yield information about later stages of stimulus evaluation, involving cognitive functions such as attention and memory. Clinical observations and neuropsychological evaluations of cognitive abilities in adult and young migraineurs indicate an association between migraine (especially with aura) and short- and long-term impairment of cognitive abilities, such as memory, attention, visuomotor processing and reasoning (12–15). In this view, the recording of modifications of electrical brain activity that are largely dependent on psychological variables, i.e. the event-related potentials (ERPs), represents a useful method to investigate the temporal succession of elementary cognitive processes.
Both adult and young migraine sufferers are generally reported to show higher interictal contingent negative variation (CNV) amplitude compared with controls, indicating increased cortical excitability and abnormal orienting activity (16, 17). There is evidence that the higher mean CNV amplitude recorded in migraineurs is the result of retarded habituation over blocks of trials (18–20).
Compared with CNV data, the results of ERP studies employing the oddball paradigm to investigate cognitive processing in migraine sufferers during the pain-free period are much less conclusive. Adults suffering from migraine without aura have been reported to show prolonged latency and reduced amplitude of target P300 compared with controls (21, 22). Other investigators did not find any significant difference between pain-free migraineurs without aura and tension-type headache sufferers in either P300 amplitude or latency elicited in an oddball task (23). The lack of a control group, however, makes it difficult to give this evidence a clearcut meaning.
Lack or deficit of normal habituation of target P300 amplitude and latency was observed in adult migraineurs without aura in the interictal interval (9, 24–27). These results, together with the above-mentioned VEP and CNV data, suggest that deficient habituation across repetitive stimulation may represent a general characteristic of migraine patients between attacks, affecting both early and advanced steps of information processing.
Migraine children have been reported to show increased P300 amplitude and reduced habituation of both P300 amplitude and latency compared with healthy controls (28). Therefore, some abnormalities in cognitive processing are thought to be already detectable in young migraineurs, and not only in adult patients.
Although most electrophysiological studies on cognitive functions in migraineurs have focused on P300 data, the amplitude and/or latency of earlier ERP components, i.e. N100, P200 and N200, have also been assessed (21–29). Whereas no differences between patients and controls have been repeatedly reported for other ERP components, some results indicate that auditory N100 latency shows abnormal features in migraineurs. Specifically, N100 latency was reduced in adult patients and prolonged in migraine children, compared with controls (22, 29). Therefore, thorough investigation of N100 latency abnormalities deserves further consideration.
Studies reporting reaction times (RTs) and error data (omissions to targets and responses to standards) recorded during an oddball paradigm either document no significant differences between pain-free patients and controls in mean RTs (26, 28), or longer RTs and more errors in migraineurs than in controls (22). Thus, it is not entirely clear if some kind of cognitive impairment in migraine patients is also reflected in the level of speed and quality of task performance.
An expansion of research on cognitive processing in juvenile migraine by means of ERPs and performance data recording is needed in order to understand better which features differentiate young patients from healthy children, and to clarify their functional and pathophysiological meaning. The present study was aimed at investigating ERPs, measured interictally during an auditory oddball paradigm, in children suffering from migraine without aura. In addition to N100 and P300 latency and amplitude, the mismatch negativity (MMN) was also computed. This negative deflection is thought to reflect an early, preattentive mismatch-detection process activated by a change in repetitive auditory stimulation (30). To our knowledge, MMN in migraine patients is an issue which has never been addressed by researchers so far. By evaluating MMN in migraine children we wanted to investigate the possible association of the migraine disease with abnormalities in preattentive processes, such as auditory sensory memory and automatic change detection. Strict criteria for sample selection and control age-matching were employed, thus avoiding possible confounding variables due to sample inhomogeneity. Measurement of RTs and errors was also utilized, in order to assess speed and accuracy of behavioural responding and allow correlation of ERP component amplitude and latency with behavioural performance.
Our evaluation was focused on the interictal interval, since we were interested in the putative cognitive abnormalities that may represent a basal condition in the migraine individual (9).
Methods
Participants
Eighteen patients (eight males, 10 females) participated in the study. All the participants were out-patients of the Department of Paediatrics, University of Padova. Mean age was 10.6 ± 2.1 years (range 8–14 years).
Physicians experienced in headache diagnosis had documented they were affected by migraine without aura according to the diagnostic criteria of the International Headache Society (31). Inclusion criteria were: at least one migraine episode per month, at least 6 months’ illness duration, last migraine episode at least 3 days prior to investigation. Neurological examination had excluded the presence of any neurological disorder. The clinical data of the patients are presented in Table 1.
Clinical data of the migraine children

At the time of investigation no patients were taking any prophylactic medication or receiving non-pharmacological treatments.
Their participation in the study took place on a symptom-free day.
The control group included 18 age-matched healthy participants (11 males, seven females), mainly recruited in local schools and sport/leisure centres. They had no recurrent headache episodes, or first-degree relatives affected by migraine. Mean age was 10.7 ± 2 years (range 8–14 years).
Procedure
Written informed consent was obtained after the nature and aim of the study had been fully explained to the parents of the children.
Participants were seated in a comfortable armchair in a quiet, light-attenuated room and were presented with a white fixation cross at the centre of a 19-in computer monitor placed 1 m in front of them. Three hundred stimuli were presented in three blocks of 100 trials each. Standard (80%) and target (20%) tones (1000 and 2000 Hz sinusoids, 100 ms duration, 65 dB SL) were delivered binaurally through an earphone every 2 s, in a pseudorandom fashion (no two targets were presented in succession; the number of standard stimuli between targets varied between two and six, each appearing four times per block). Tones presentation and RT recording were accomplished by MEL 2.0 software (Micro Experimental Laboratory, Psychology Software Tools, Inc., Pittsburgh, PA, USA) (32).
Participants were instructed to keep their eyes on the fixation cross and to limit eye movements to spontaneous blinking for the entire task duration. The space bar on a comp Keyboard had to be pressed with the forefinger of their dominant hand as soon as a target was presented. Mean RTs were recorded in ms from the pressing of the space bar, and the proportion of errors (false alarms, omissions, and total) was calculated.
At the end of each block, a notice on the monitor signalled the participant that it was possible to take a pause and that he/she was free to operate the beginning of the following block by pressing the space bar on the keyboard.
One training run of stimuli (including 10 standards and three targets) was done before starting data recording, to ensure that all participants understood the procedure. Total task duration was about 15 min.
ERP recording and measurement
The electroencephalogram (EEG) was recorded from Fz, Cz and Pz according to the 10–20 International System (33) with three surface Ag/AgCl electrodes referred to linked mastoids. Electrode impedance was kept below 10 kΩ. Vertical and horizontal eye movements were recorded with electrodes placed above and below the left eye and on the outer canthi of both eyes. High-pass and low-pass filters were set at 0.16 Hz and 40 Hz, respectively. Sampling rate was 500 Hz. Signals were filtered and amplified by a SynAmps unit amplifier (Neuroscan, Inc., Compumedics, Ltd, El Paso, TX, USA).
The EEG was corrected for eye movements and blink artefacts using a regression-based weighting coefficients technique (34), as implemented in the SCAN 4.1 software (Edit module; Neurosoft, Inc.). EEG epochs of − 200 to 1000 ms poststimulus, baseline corrected by subtraction of the average prestimulus voltage, were averaged separately for rare and frequent tones. Trials with signal exceeding ± 100 µV amplitude in any recording channel were excluded from averaging. For the target tone stimuli, only correctly detected responses were averaged; for the standard stimuli, false alarms were rejected. The mean number of target and standard epochs in the average was 229 and 57, respectively.
After visual inspection of grand averages and individual averages, peak amplitudes in the 70–150 and 250–400-ms windows from stimulus presentation were calculated for N100 and P300, respectively. Peaks were automatically quantified in the above-indicated windows by a peak detection option in SCAN 4.1 software, and visually checked for accuracy by a trained experimenter blind as to subjects’ group. If the program made inaccurate measures due to broad or multiple peaks, the experimenter corrected the measurement and the new value was recorded. Latency was calculated as the time of peak from stimulus presentation.
In addition, ERPs to the frequent stimuli were subtracted from ERPs to the deviant stimuli to obtain a difference wave. Peak amplitude in the 170–300-ms window in Fz was calculated as an estimation of the MMN, and its latency was also measured.
P300 habituation was quantified by evaluating amplitude and latency in the three blocks separately.
Statistical analysis
Repeated measures
Unadjusted P-values for effects within variables having more than two levels are reported, together with the Geisser–Greenhouse epsilon (35).
Newman–Keuls tests (P < 0.05) were used for post hoc comparisons.
Student's t-test for independent samples (two-tailed) was used to compare the two groups in MMN amplitude and latency, and in mean RTs and error proportions. RTs in the three blocks were assessed by means of a 2 (Group: Controls and Migraineurs) × 3 (Block: I, II, III)
Correlations between behavioural (RTs and proportions of response errors), clinical (illness duration and frequency of attacks) and electrophysiological (ERP components amplitude and latency) data were computed. Pearson product-moment correlation coefficients (r) are reported.
Results
Tables 2a and 2b report
Results of 2 (Group) × 2 (Stimulus) × 3 (Site)

Results of 2 (Group) × 3 (Site) × 3 (Block)

N100
Figure 1 shows the grand averages for target and standard stimuli in juvenile migraineurs and controls.

Grand average waveforms at Fz, Cz and Pz for target (right panel) and standard stimuli (left panel) in controls and migraineurs.
A significant group–stimulus interaction was found, showing that N100 amplitude to standard stimuli was significantly smaller in migraineurs than in controls (Fig. 2).
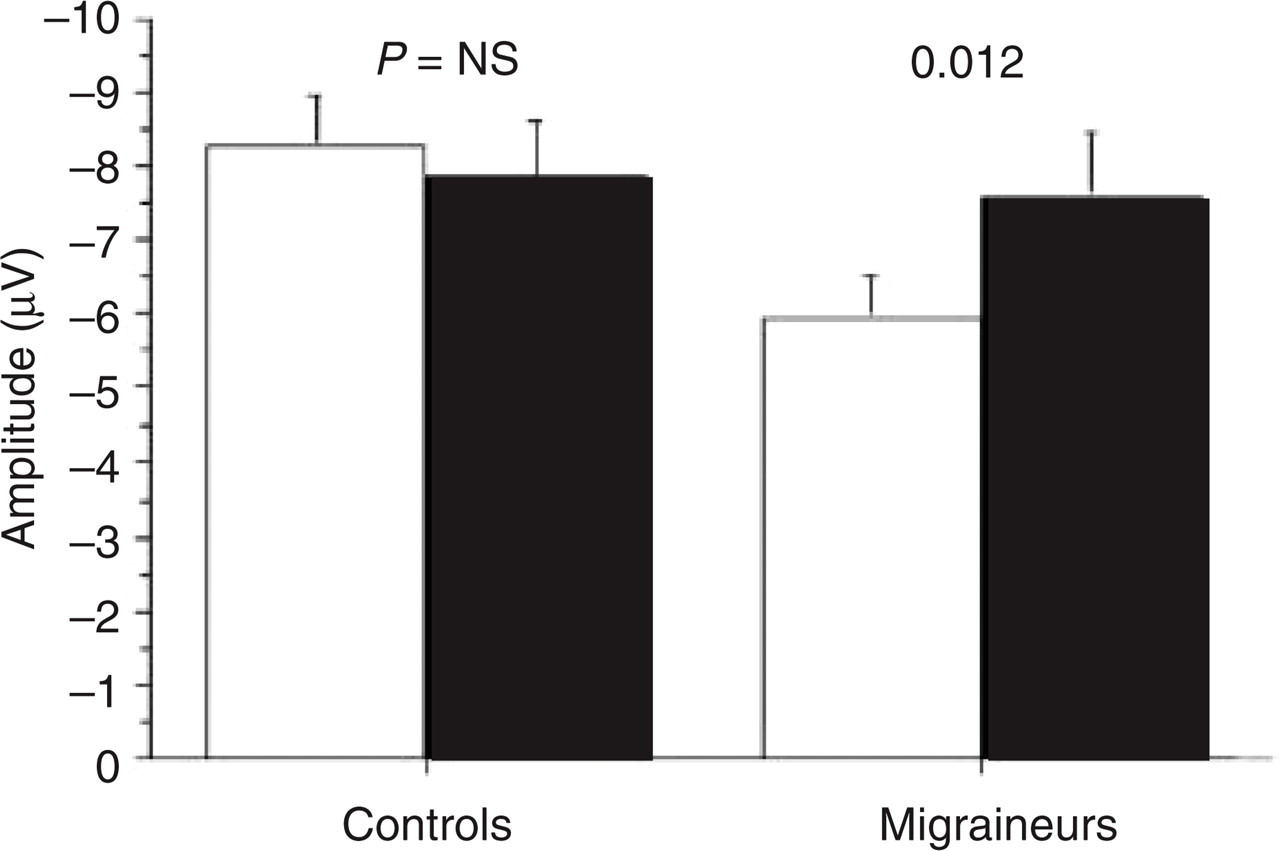
N100 amplitude for standard and target stimuli in controls and migraineurs. □, Standard; ▪, target.
N100 amplitude had a frontal distribution, being larger at Fz (−11.2 µV) compared with Cz (−7.4 µV) and Pz (−3.68 µV). The difference between Cz and Pz was also significant.
The stimulus–site interaction showed that target N100 was significantly greater than standard at Cz and Pz, but not at Fz. Moreover, standard N100 amplitude was not significantly different in Cz compared with Pz.
Significant variations in N100 latency were found as a function of site, with the longest latency recorded at Fz (119.5 ms), followed by Cz (109.75 ms) and Pz (105.4 ms). Each site differed significantly from the other two.
No other significant effect emerged for N100 latency.
MMN
The t-test yielded no significant differences between the two groups, either for MMN amplitude (t = 1.32, P < 0.194) or for its latency (t = 1.28, P < 0.209). Nevertheless, it is worth noting a tendency for migraineurs to show greater amplitude and shorter latency compared with controls (−3.30 µV and 221.03 ms vs− 0.931 µV and 236.09 ms; see Fig. 3).

Difference waveforms (target minus standard) of the grand-average event-related potentials at Fz in controls and migraineurs.
P300
Target stimuli showed significantly greater P300 amplitude than standards.
P300 amplitude also varied between sites, being parietally maximal (Pz: 14.17 µV) and frontally minimal (Fz: 4.14 µV), with intermediate values at Cz (10.36 µV). Post hoc tests showed that amplitude at each site differed significantly from the other two.
The difference between P300 amplitude to rare and frequent stimuli was more evident in Pz compared with Cz and Fz, even though post hoc tests revealed that the differences between the two stimuli were significant in all three sites.
P300 latency showed a marginally significant group–stimulus interaction (F[1,34]= 3.18, P < 0.08), indicating that P300 latency for frequent stimuli was longer in migraineurs than in controls. Moreover, controls displayed approximately the same latency for standards and targets, whereas in migraineurs a slower latency was observed for frequent than for rare stimuli.
The significant site main effect indicated that latency was longest at Fz (330.25 ms) compared with Cz (309.5 ms) and Pz (303.32 ms). The difference between Cz and Pz was non-significant.
Evaluation of target P300 amplitude in the three blocks showed a main effect of site, with maximal amplitude at Pz (24.43 µV), intermediate at Cz (18.88 µV), and minimal at Fz (8.54 µV). As indicated by post hoc tests, each site differed significantly from the other two.
A significant P300 amplitude reduction across blocks was observed; post hoc tests showed that only block I and block III significantly differed from each other (18.65 and 15.67 µV, respectively).
The significant group–block interaction showed that target P300 amplitude dropped in the third block in migraine children, whereas controls showed no significant change across task.
The group–block–site interaction illustrates how the above-mentioned effect was stronger at Fz and Cz compared with Pz (Fig. 4).

P300 amplitude to target stimuli at Fz, Cz and Pz in the three blocks in controls and migraineurs. ♦, Controls; □, migraineurs.
The same analysis performed on P300 latency showed a block main effect, with P300 latency increasing significantly from block I (307.29 ms) to block III (317.75 ms). Block II (312.76 ms) did not differ significantly from blocks I and III.
A main effect of site showed that latency was significantly longer at Fz (326.63 ms) than Cz and Pz (305.6 and 305.55 ms, respectively). The difference between Cz and Pz was non-significant.
Task performance
RT data from one patient were lost because of equipment failure, therefore the age-matched control was also excluded from statistical analysis. As a consequence, for RTs the number of subjects in each group is 17.
Speed of response (RTs) was equivalent in the two groups (t = 0.99, NS; migraineurs = 378.50 ± 79.48 ms, controls = 404.26 ± 71.16 ms). Although performance accuracy was fairly high in both groups, the total proportion of errors (false alarms and omissions) was significanly higher in migraine children than in controls (1.81 and 0.33%, respectively; t =−2.13, P < 0.04; Fig. 5). Separate analysis for omissions and false alarms showed significance for omissions only.

Mean proportion of errors (omissions and false alarms) in controls and migraineurs.
Evaluation of RTs in the three blocks only revealed a block main effect (F [2,64]= 6.47, P < 0.002, ε= 0.79), showing that RTs increased significantly from block I (379.32 ms) to block III (399.71 ms). Block II (395.09 ms) did not differ significantly from block III.
Correlations
N100 and P300 latency and amplitude for standard and target stimuli were correlated with RTs and proportions of errors. In the patients group, target P300 latency at Pz was significantly correlated with mean RTs, showing that longer latencies corresponded to slower RTs (r = 0.64; P < 0.006; the correlations for the other sites were only marginally significant). Moreover, target P300 amplitude at Fz and Cz (and marginally at Pz) showed a significant negative correlation with mean RTs (r = −0.65; P < 0.004, and r =−0.53; P < 0.026, respectively), indicating that smaller P300 amplitude corresponded to longer RTs.
Illness duration and frequency of attacks were correlated with behavioural (RTs and proportions of errors) and ERP data (N100 and P300 amplitude and latency of standard and target stimuli at Fz, Cz and Pz) in the patients group. A marginally significant correlation (r = 0.45; P < 0.056) was found between illness duration and standard N100 amplitude recorded at Fz, showing that longer disease durations corresponded to smaller standard N100 amplitudes.
The other correlations were non-significant. No significant correlations emerged in the controls group.
Discussion
A significant group difference in ERPs emerged for N100 amplitude to standard stimuli, with migraine children showing smaller amplitude than healthy controls. Selective attention has been shown to modulate N100 amplitude, which is larger to attended stimuli compared with ignored ones (36). Unlike controls, migraine children showed significant amplitude difference between standard and target N100, which may indicate they focused their attention on task-relevant stimuli while ignoring the irrelevant ones.
On the other hand, experimental evidence suggest that the auditory N100 has multiple subcomponents, one of which is supposed to be sensitive to alertness level (37). Indeed, N100 amplitude reduction reflects decrements in arousal or vigilance level (38, 39). The smaller N100 amplitude to the frequently occurring stimuli in patients compared with controls might indicate a lower level of alertness, possibly due to the highly repetitive stimulation features (40).
Correlations between behavioural, clinical and electrophysiological data showed that in migraine children longer illness durations were marginally correlated with smaller standard N100 amplitudes. This finding indicates that the overall standard N100 amplitude reduction observed in the group of patients compared with controls is directly related to the length of time each child has been suffering from migraine. Indeed, there is at least some evidence in the literature suggesting a relationship between disease duration and VEP alterations in adult migraineurs (41, 42). These data, together with our findings, would favour the viewing of some electrophysiological abnormalities in migraineurs as a consequence of the disease, or a perpetuating factor, more than a pathogenic characteristic.
The observed tendency towards greater amplitude and shorter latency of MMN in young patients compared with controls may indicate an enhanced orienting response towards physical stimulus change.
Overall, differences in information processing between migraine children and healthy controls may involve early attentive functions and/or level of arousal, but further study is needed to corroborate this assumption and clarify its theoretical and practical implications.
A somehow striking finding, that target P300 amplitude habituated in migraine children but not in controls, contradicts the literature data about VEPs, CNV and, partially, the oddball P300, indicating lack of habituation in migraineurs. One possible explanation is related to the fact that P300 amplitude is sensitive to motivation, insofar as increased motivation results in greater amplitude (43). The observed lack of P300 amplitude decrease across blocks in controls may indicate that the same degree of motivation was maintained for the whole task duration. In contrast, migraine children may have undergone a decline in task involvement due to motivational deficit or fatigue.
The significant positive correlation between RTs and target P300 latency in the patient group is an indication that overt response was emitted after evaluation of the task-relevant stimulus was completed. However, speed of response was inversely related to the amount of attentional resources employed for stimulus processing (represented by P300 amplitude), indicating that when fewer resources were used, motor response was delayed. This pattern of relationships between speed of response and P300 features may also be related to the poorer performance accuracy observed in migraine children compared with controls, even though correlations with the proportion of errors were non-significant.
It is also possible that the habituation deficit found for target P300 latency of migraineurs in visual active oddball paradigms (25) is related, at least in part, to the hyperexcitability of the visual cortex, which has been repeatedly documented by VEP recording in migraine with and without aura (4). Hypersensitivity of other brain sensory areas in migraine patients is also supposed on the basis of empirical evidence (44), but it might play a less prominent role in determining deficient habituation when auditory stimuli are employed in active oddball tasks.
Another important aspect of migraine pathology might explain the P300 amplitude decrease in our group of patients. In adult migraineurs, habituation of VEPs and intensity dependence of AEPs (45, 46), habituation of P300 latency (26) and CNV amplitude (47, 48) tend to show maximal abnormality shortly before the attack, then normalize during the attack and for the next few days after the crisis. Periodic changes are also present in young migraineurs’ CNV amplitude and habituation (49). In the subjects selected for our study, the last attack had occurred at least 3 days prior to the investigation, therefore all the patients had presumably completed the normalization period. However, the delay between the day of recording and the next ictal episode was not recorded in our present investigation. Considering the wide range of frequency of attacks in our patient sample (one to eight per month), the time interval to the next crisis is also supposed to be greatly variable. Therefore, the patients might have happened to be investigated in very different phases of the migraine cycle, ranging from the refractory interval following the attack to the preattack period. As a consequence, varying degrees of interictal abnormalities could have been detected. This source of uncontrolled variability might be partially responsible for our findings and represent a potential limitation.
Some methodological reasons could also account for P300 amplitude habituation in our group of migraine children. The lack of target P300 amplitude habituation (and, indeed, potentiation) in migraine subjects has been found in studies using acoustic passive oddball tasks (9, 24). This condition is probably more sensitive for the process of habituation to emerge in controls, since repetitive stimulation is not associated with the active attentional engagement that is necessary to detect and respond to task-relevant stimuli (50). The lack of P300 amplitude habituation in controls might indicate that they succeeded in maintaining cognitive involvement and attentional allocation over the whole task, whereas its decrease in migraine children may reflect their gradual withdrawal of attentional engagement and/ or motivation. Similar considerations have been recently proposed by other authors, albeit stimulated by results obtained with adult migraineurs in a ‘conditioning-testing’ paradigm (51).
Lastly, other methodological reasons for the discrepancies between our results and those reported in the literature, as well as among different studies on this topic, include several procedural features of the oddball paradigm known to affect ERP components, such as number of trials, length of intertrial interval, and standard/target ratio, and other elements related to diagnostic criteria for patient selection and sample size (52). Research projects adopting converging criteria in task construction and sample selection could address and possibly overcome inconclusiveness and inconsistencies in the results.
The electrophysiological and behavioural data obtained in this study support the view that children suffering from migraine without aura show some evidence of cognitive disturbances, possibly involving attentive and, less substantially, preattentive functions. Difficulties in adjusting their alertness level and motivational engagement to task demands may also be important elements contributing to the general picture.
Systematic investigation of ERPs in childhood migraine should be pursued to elucidate which malfunctions of cognitive processes are distinctive features of migraine pathology and which might be related to adjustments to the illness and are possibly shared by other pain-related diseases.
Footnotes
Acknowledgements
The authors are grateful to Sandro Bettella for programming assistance and to Sonia Mele for her help in data collection.