
Abstract
Following cerebral ischemia, perilesional astrocytes and activated microglia form a glial scar that hinders the genesis of new axons and blood vessels in the infarcted region. Since glial reactivity is chronically augmented in the normal aging brain, the authors hypothesized that postischemic gliosis would be temporally abnormal in aged rats compared to young rats. Focal cerebral ischemia was produced by reversible occlusion of the right middle cerebral artery in 3- and 20-month-old male Sprague Dawley rats. The functional outcome was assessed in neurobehavioral tests at 3, 7, 14, and 28 days after surgery. Brain tissue was immunostained for microglia, astrocytes, oligodendrocytes, and endothelial cells. Behaviorally, aged rats were more severely impaired by stroke and showed diminished functional recovery compared with young rats. Histologically, a gradual activation of both microglia and astrocytes that peaked by days 14 to 28 with the formation of a glial scar was observed in young rats, whereas aged rats showed an accelerated astrocytic and microglial reaction that peaked during the first week after stroke. Oligodendrocytes were strongly activated at early stages of infarct development in all rats, but this activation persisted in aged rats. Therefore, the development of the glial scar was abnormally accelerated in aged rats and coincided with the stagnation of recovery in these animals. These results suggest that a temporally anomalous gliotic reaction to cerebral ischemia in aged rats leads to the premature formation of scar tissue that impedes functional recovery after stroke.
Studies of stroke in experimental animals have demonstrated the neuroprotective efficacy of a variety of interventions, but most of the strategies that have been clinically tested failed to show benefit in aged humans. One possible explanation for this discrepancy between experimental and clinical studies may be the role that age plays in the recovery of the brain from insult. Most experimental studies of stroke have been performed in young animals, and therefore may not fully replicate the effect of ischemia on neural tissue in aged subjects (Davies et al., 1995; Futrell et al., 1991; Popa-Wagner et al., 1998, 1999; Sutherland et al., 1996; Wang et al., 1995). A better understanding of the effects of senescence on the response of the brain to stroke and other forms of injury is needed to develop effective new treatment strategies.
Following ischemic stroke, microglia and astrocytes show both an increase in number as well as a strongly reactive phenotype in the infarcted region (Mabuchi et al., 2000; Stoll et al., 1998), and oligodendrocytes proliferate in the outer border of the infarct (Mandai et al., 1997). Glial cells thus may hold important clues to the degree of neural and behavioral recovery from stroke. In the infarct border, the processes of hypertrophic and hyperplastic reactive astrocytes extend to encompass the developing infarct, while rounded, activated microglia engulf cellular debris. These two cell types form the basis of the glial scar (Ellison et al., 1998; Gehrmann et al., 1992; Lehrmann et al., 1997). In addition, oligodendrocyte lineage cells contribute a number of molecules to the glial scar, particularly chondroitin sulfate proteoglycans (such as NG2, phosphacan, brevican, versican, and neurocan), which have been hypothesized to impede the regeneration of tissue (Asher et al., 2002; Chen et al., 2002; Morgenstern et al., 2002; Niederost et al., 1999).
Because the brains of normal, aged subjects are characterized by increased astrocytic and microglial reactivity (Goss and Morgan, 1995; Jucker et al, 1994; Nichols et al., 1995; Peters, 2002; Vogelgesang et al., 2002; Yu et al., 2002), we hypothesized that these cells might respond abnormally to stroke in aged rats, thereby contributing to age-related impairments in functional recovery. To test this hypothesis, we studied the response of astrocytes, microglia, and oligodendrocytes to middle cerebral artery occlusion in young and aged rats. Our findings indicate that a glial scar develops abnormally early in the infarcted region of aged rats, and may thereby impede the functional recovery of surrounding nervous tissue.
MATERIALS AND METHODS
All experiments were conducted in accordance with the statement regarding the care and use of animals and were approved by a federal animal care committee.
Animals
Young (3–4 months, n = 12 per age group and time point) and aged (18–20 months) male Sprague-Dawley rats, bred in-house, were used for all experiments. Body weights ranged from 290 to 360 g in the young rats and 420 to 500 g in the older rats. The animals were behaviorally tested and then subjected to reversible occlusion of the middle cerebral artery. The group of older rats kept alive for 4 weeks was larger (n = 18) to compensate for the higher postischemic mortality rate. Sham operations were also conducted by pulling back the hook within 60 seconds (n = 8, per age group and time point).
Behavioral tests
To evaluate various aspects of neurologic function before surgery, the rats were tested for their ability to traverse a rotating horizontal rod, limb placement while approaching the edge of a table, and exploration of a novel environment. After surgery, the rats were also tested for sensory function. Two persons conducted each test independently, and all sensorimotor testing was performed between 9 and 11 am.
Neurologic status (Bederson test)
After surgery, rats were observed for circling behavior if pulled gently by the tail. Rats circling toward the infarcted brain side were ranked as grade 3, rats that circled freely (not held by the tail) were graded as 2, and rats showing no reaction were graded as 1.
Spontaneous activity
Each rat was placed in front of a large cage with a small entryway. After entering the cage, the number of crossings between two walls was counted within a 30-minute test period (Garcia et al., 1995).
Rotating beam–traversing task (Rotarod)
Prior to surgery the rats were conditioned to traverse the Rotarod as a measure of motor coordination and balance. The rod, which consisted of a plastic cylinder 45 mm in diameter and 1.5 m long, rotated alternately to the left or to the right at a speed of 4 rpm. The rat was placed on the Rotarod and the time taken for the rat to traverse the rotating cylinder and join a group of rats visible at the finish line was measured. Each time a rat fell from the rod onto a cushioned surface, the clock was stopped and a fall was recorded. Rats will normally cross the cylinder with their feet squarely on the top surface of the rod. Using either a forelimb or hindlimb on the side of the beam was considered to be a fault (Kolb and Whishaw, 1983). The rats were tested every other day starting the first day after surgery.
Inclined plane
The inclined plane was constructed according to the methods of Rivlin and Tator (1977) and was designed to measure the rat's ability to maintain its position at a given angle. The relative angle at which the rat fell was taken as a measure of functional impairment after the surgery. The test was conducted once before surgery and daily after surgery.
Symmetry of limb movement
The scores in the symmetry of limb movement and body proprioception tests relate to the occurrence of cortical infarcts on the right side. To assess the symmetry of limb movement, the rat was gently held in the air by the tail and limb movements were scored as follows: 3, all four limbs extended symmetrically; 2, limbs on the left side extended more or less slowly than those on the right; 1, limbs on the left side showed minimal movement; and 0, forelimb on the left side did not move at all (Garcia et al., 1995).
Body proprioception
The rat was touched lightly with a blunt stick on each side of the body, and the reaction to the stimulus was observed and scored as follows: 3, rat reacted by turning its head and was equally startled by the stimulus on both sides; 2, rat reacted slowly to the stimulus placed on the left side; and 1, rat did not respond to the stimulus placed on the left side.
Surgery
Blood flow through the middle cerebral artery was transiently interrupted as previously described (Popa-Wagner et al., 1998). Throughout surgery, anesthesia was maintained by spontaneous inhalation of 1% to 1.5% halothane in a mixture of 75% nitrous oxide and 25% oxygen. Body temperature was controlled at 37°C using the Homeothermic Blanket System (Harvard Apparatus). The anesthetized animals were immobilized in a supine position and the tail artery was catheterized to enable the continuous measurement of blood pressure and collection of blood samples for determination of pH and blood gasses (Blutgassystem IL 1620, Instrumentation Laboratory, Munich) and arterial glucose levels (Omnican7 Balance, B. Braun, Melsungen). The middle cerebral artery was slowly lifted with a tungsten hook attached to a micromanipulator (Maerzhaeuser Precision Micro-manipulator Systems, Fine Science Tools) until blood flow through the artery was completely interrupted. The common carotid arteries were then occluded by tightening the pre-positioned thread loops. The surgical field was kept warm and moist with artificial cerebrospinal fluid (Liquicheck, Spinal Fluid Control, BioRad) for the duration of the procedure. The local changes in blood flow were monitored using a laser Doppler device (Perimed, Stockholm, Sweden).
After 70 minutes, the middle cerebral artery and the common carotid arteries were reopened, allowing full reperfusion of the brain. After survival times of 3, 7, 14, or 28 days, the rats were deeply anesthetized with 2.5% halothane in 75% nitrous oxide and 25% oxygen and perfused with buffered saline followed by buffered 4% freshly depolymerized paraformaldehyde. The brain was removed, postfixed in 4% buffered paraformaldehyde for 24 hours, cryoprotected in 20% sucrose prepared in 10-mmol/L phosphate-buffered saline, flash frozen in isopentane, and stored at −70°C until sectioning.
Light microscopy
A Nikon microscope was used for light microscopy. Images (768 × 1,024 pixels) were captured electronically using a CCD camera (Optronics). The digital images were arranged and labeled using Adobe Photoshop software (Adobe Systems, San Jose, CA, U.S.A.) and printed using a Kodak XLS 8000 digital printer (Eastman Kodak Co., Rochester, NY, U.S.A.). For a group of micrographs, the camera settings for exposure, gain, and contrast enhancement were similar.
Statistical analysis
The main effect of age and time and the interaction between these two variables were evaluated using a two-way analysis of variance followed by Tukey post hoc analysis to detect differences between the means using SPSS software (SPSS Inc., Chicago, IL, U.S.A.). The level of significance (two-tailed threshold) was set at P≤0.05.
Immunohistochemistry
Sections (25 μm) were cut on a freezing microtome and processed for immunohistochemistry as free-floating material as previously described (Popa-Wagner et al., 1998, 1999). To obtain uniform staining of all tissue sections (about 360 per experiment), we constructed a semiautomated device (PCT, device assembly for preparing and analyzing tissue for microscopic examinations, DE 199 45 621 A1 and WO 01/22052 A1, 2001), making it possible to process and stain all sections simultaneously under identical conditions.
In short, after blocking in 3% donkey serum, 10-mmol/L PBS, and 0.3% Tween 20, sections were incubated overnight at 4°C with monoclonal antibodies recognizing one of the following: (1) a cytoplasmic determinant of brain macrophages (clone ED1, Serotec, UK); (2) the astrocytic marker glial fibrillary acidic protein (GFAP) (clone G-A-5; Boehringer Mannheim, Mannheim, Germany); (3) the rat endothelial cell antigen (RECA) (clone HIS52, Serotec); (4) the oligodendrocytic marker chondroitin sulfate proteoglycan, NG2 (Chemicon, Hofheim, Germany); or (5) the neuronal marker, NeuN (Chemicon) diluted in PBS containing 3% normal donkey serum and 0.3% Tween 20. The primary antibody was detected using the ABC system (Vectastain Elite Kit; Vector, Burlingame, CA, U.S.A.).
Determination of infarct volume
To assess the size of the infarct induced by transient focal ischemia, every twentieth section was stained with an immunologic marker of neuronal viability (NeuN). Low-power images of the stained sections were taken with a video camera and then printed. Then the infarcted area was demarcated using a pencil and the images were scanned. Finally, the area and partial volume of every twentieth section of the ipsilateral cerebral hemisphere were obtained using the NIH image analysis program. Integration of the resulting partial volumes gave the volume of the ipsilateral hemisphere along with the volume of the cortical infarct; lesion volume was then expressed as percent of the total volume of the hemisphere.
RESULTS
Behavior
Twenty-four hours after surgery, the clinical status of the animals was assessed using the Bederson test. At this point, about 70% of animals that had undergone cerebral ischemia showed obvious neurologic deficits, including paresis of the contralateral extremities, particularly the hind legs, and rotation when lifted above the cage floor. Although hematologic parameters showed some age-associated differences, notably in blood pressure, the differences were not statistically significant (Table 1).
Serum arterial blood gas, glucose, and blood pressure values
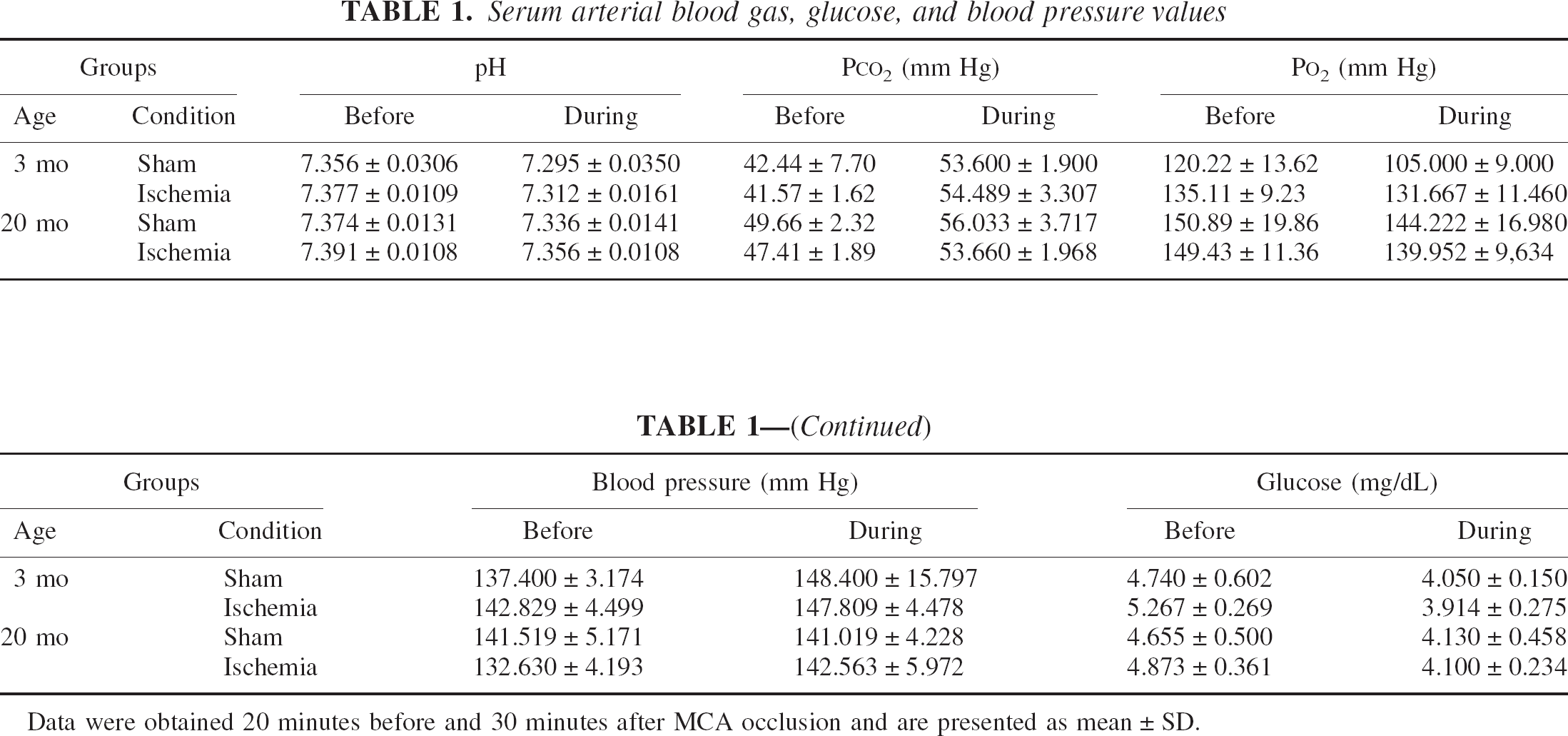
Data were obtained 20 minutes before and 30 minutes after MCA occlusion and are presented as mean ± SD.
Before surgery, the only statistically significant difference in performance between the two age groups was a 30% decrease in spontaneous activity of the aged rats relative to the young animals (Fig. 1E). Following infarction, most infarcted animals showed disturbances of sensorimotor function, with aged animals recovering more slowly, if at all, in some tests. As a general observation, all rats had diminished performance on the first postsurgical day, part of which was attributable to the surgery itself (as evidenced by impaired Rotarod performance in sham-operated rats, Fig. 1B). The decrease in performance on day 1 also depended on both age and the difficulty of the testing. For instance, in the Rotarod test, aged rats showed a decline of about 50% on day 1 while the young rats showed a decline of about 30% (Fig. 1D). In addition, young rats fully recovered after 3 to 14 days, depending on the type of test, while the aged rats recovered only to about 70% of presurgery performance levels. The differences in the degree of recovery between the two age groups were statistically significant except for spontaneous activity (Fig. 1E) and symmetry of limb movement (Fig. 1C), probably because of the high interanimal variability among aged rats on these tests. There were, however, two notable exceptions regarding the age-related differences in the extent of recovery: first, the extent of recovery in spontaneous activity was rather low for both young and aged animals (Fig. 1E); second, after a brief period of improvement in the first 3 days of recovery, performance on the inclined plane test declined among aged rats after day 3 and never returned to control values. By comparison, the young rats easily mastered the inclined plane task during the recovery period (Fig. 1F). Similarly, there was poor recovery of bodily proprioception in aged rats (Fig. 1A). Note that the sham-operated rats showed recovery rates that exceeded 100%, which could be attributable to a training effect (Fig. 1B).
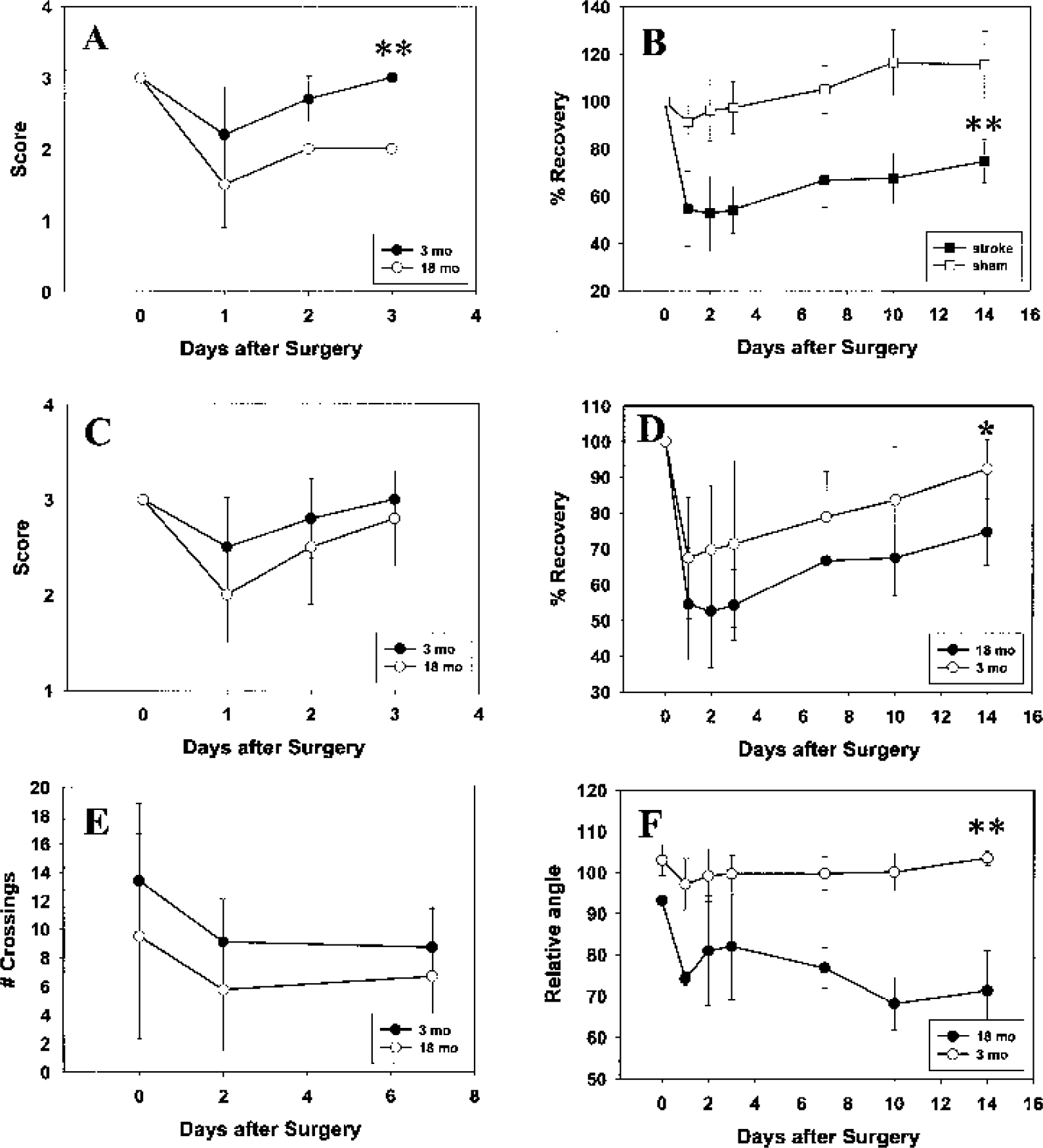
Functional recovery (
Histology
Acid-vanadium-fuchsin staining, a marker of neuronal degeneration, revealed age-related differences in the viability of cells around the infarct core. The intensity of staining for necrotic nuclei at day 14 in the penumbra was greater in young animals (Fig. 2A) than in aged animals (Fig. 2B). The infarct core was also visualized by immunohistochemistry for NeuN, a sensitive marker of neuronal degeneration (Figs. 2C and 2D). Based on this stain, the areas most affected by stroke were the parietal and, to a lesser extent, posterior frontal cortex. The morphology of viable neurons that stained positively for NeuN in the periinfarct region was not essentially different between young and aged rats (Figs. 2C and 2D, insets), though fewer viable neurons were observed in the periinfarct region of young animals (in accordance with the acid-vanadium-fuchsin staining). Measurement of the infarct volume using NeuN immunohistochemistry revealed no significant difference in the volume of cortical infarcts between young and aged rats (37% ± 5.1% vs 41% ± 3.9%, respectively).
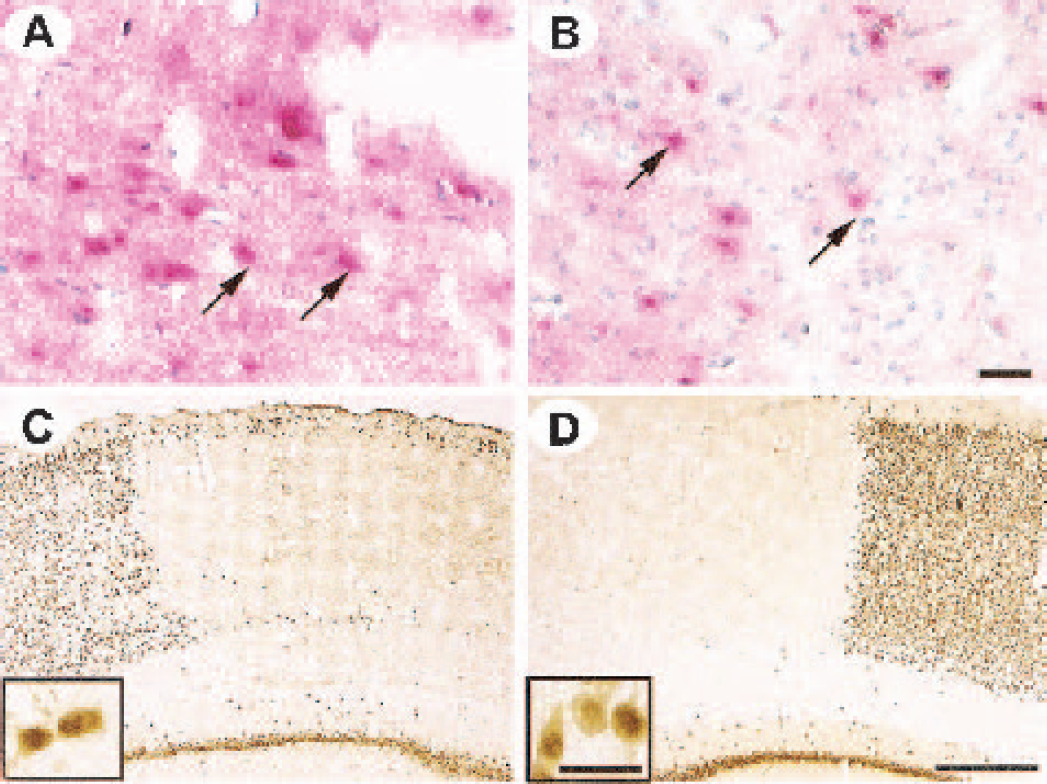
Histologic staining of the infarcted tissue. Acid-vanadium-fuchsin staining revealed that the intensity of degeneration of necrotic cells (arrows) in young animals (
Time course of ED1 expression
Using a monoclonal antibody raised against the cytoplasmic determinant of brain macrophages (clone ED1), in young rats 3 days after surgery, immunostaining was associated with a small number of cells at an incipient stage of activation (Fig. 3A, upper panel and the inset). The process of activation continued in the ensuing days so that by day 7 there were many activated microglia-like cells that still displayed distinct processes (Fig. 3B, upper panel, inset). The microglial cells were fully activated at day 14 (Fig. 3C, upper panel, inset) and remained highly activated thorough day 28 (Fig. 3D, upper panel). It should be noted that the antibody we used cannot distinguish between brain-derived microglia and blood-borne macrophages, so that some cells that we detected in the infarcted area are probably hematogenous macrophages that leaked into the lesion site through the damaged blood–brain barrier.
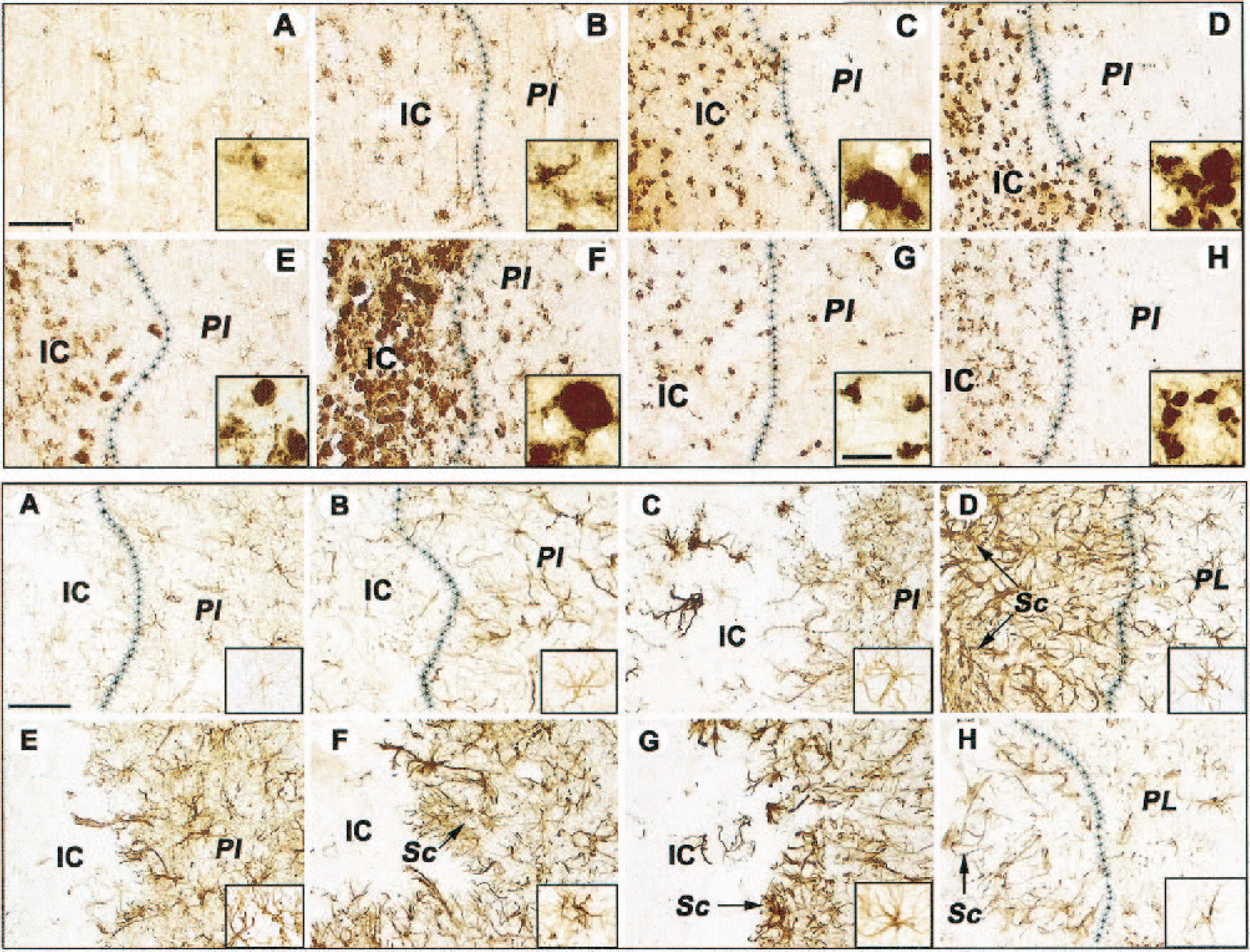
Upper panel, Development of microglial activation after cerebral ischemia. Note the gradual activation of microglia in the infarcted area of young rats at days 3 (
The situation was quite different in aged animals. Many microglia-like cells were fully activated 3 days after surgery (Fig. 3E, upper panel), and the process reached a maximum at day 7 (Fig. 3F, upper panel). Thereafter the intensity of the microglial reaction diminished progressively with time, but was still evident even at day 28 (Fig. 3G, H, upper panel, insets).
Time course of GFAP expression
Three days after the ischemic event, clusters of GFAP-immunopositive cells were occasionally observed in the ipsilateral cortex of young rats (Fig. 3A, lower panel). The cells had numerous filigreed processes resembling an early stage of the reactive phenotype, and the boundary between infarct core and periinfarct region began to become evident. By day 7, the reactive phenotype in the periinfarct zone became more prominent with enlargement of the cell bodies (Fig. 3B, lower panel), and this phenotype remained essentially unchanged at day 14 (Fig. 3C, lower panel). However, by day 28 two layers became apparent: one layer located very close to the infarct core that contained long filamentous processes that formed part of the scar tissue, and a peripheral layer containing typical reactive astrocytic morphology (Fig. 3D, lower panel). A reactive astrocytic phenotype is normally associated with advancing age and was consistently observed in the contralateral region of aged animals. However, after an ischemic episode, there was an intense astrocytic reaction in the periinfarct region 3 days after surgery, which is in sharp contrast with the mild reaction observed in the damaged cortex of young animals at the same time point. At this early stage, the cells had shorter and thicker processes than those found in young rats (Fig. 3E, lower panel), indicating a more advanced reactive phenotype. Although the reactive astrocytic phenotype persisted at the remaining time points (Figs. 3F–3H, lower panel), on day 7 the pattern in aged rats resembled that seen on day 28 in young rats; i.e., we could distinguish two layers around the core, an outer area populated with astrocytes with long and thin processes that were part of the scar tissue, and a peripheral layer populated with astrocytes with a slightly less reactive phenotype (Fig. 3 F, lower panel). Interestingly, we noted an intensification of GFAP staining in the contralateral hemisphere of aged, but not young, rats (Figs. 3E–3G, lower panel, insets).
Time course of RECA expression
We used double labeling to assess the relationship between rat endothelial cells, which form the inner lining of mature blood vessels, and astrocytic phenotype at different time points after middle cerebral artery occlusion in the two age groups. Although RECA by itself does not specifically stain a discrete cellular structure within the scar tissue, it does result in diffuse staining within the scar region, which helps to locate the scar.
In control, 3-month-old rats, the vascular tree in the parietal cortex that was most affected by stroke was intermingled with astrocytes that were evenly distributed in layers I to III (Figs. 4A–4C). In the first week after the ischemic event, a disintegration of blood vessels in the periinfarct zone, concomitant with an invasion of the infarct core by astrocytes, was apparent (Figs. 4D–4F). After 4 weeks a thin, compact scar penetrated by numerous astrocytes with long processes had formed (Figs. 4G–4I and the inset). Occasionally, newly formed blood vessels in tight association with astrocytes were visible beyond the scar (Figs. 4G and 4H).
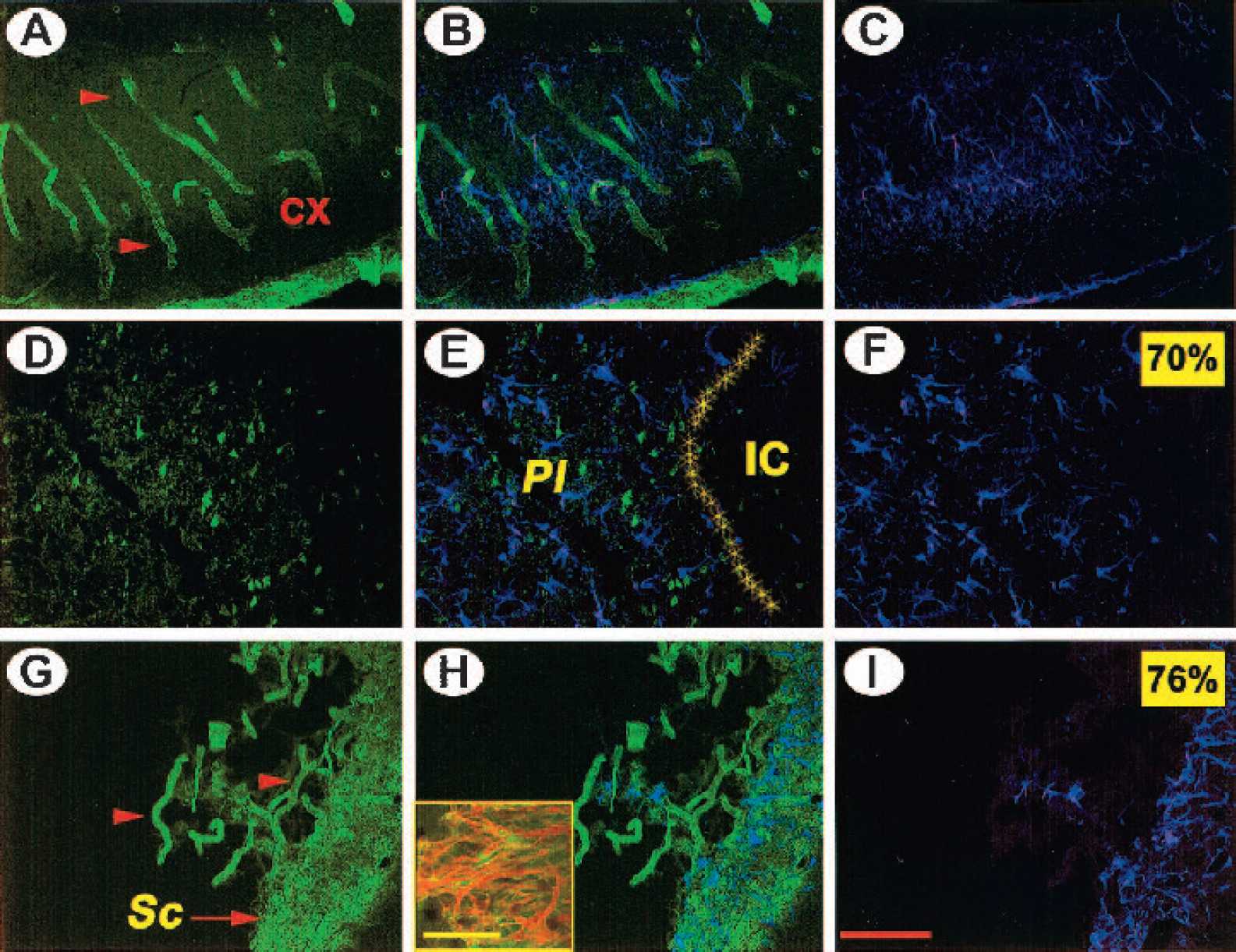
Development of scar tissue after cerebral ischemia in young rats. Double immunofluorescence for the endothelial cell marker, RECA (green), and the astrocytic marker, GFAP (violet). In the contralateral hemisphere at day 7, note the close association of astrocytes with blood vessels in cortical layers II to III (
In aged control rats, the cortical blood vessels were thinner and often had a fragmented appearance (Figs. 5A–5C). In aged, infarcted rats, a sequence of events similar to that observed in young rats also occurred, but at a much faster pace. Thus, 3 days after stroke, blood vessels were fragmented, and some RECA immunoreactivity co-localized with astrocytes in the periinfarct region (Figs. 5D–5F). By 1 week, a scar-like formation that was fully penetrated by reactive astrocytes was already apparent (Figs. 5G–5I and inset). By 14 days after stroke, the infarct core was firmly demarcated by well-formed scar tissue (Figs. 5K–5M).
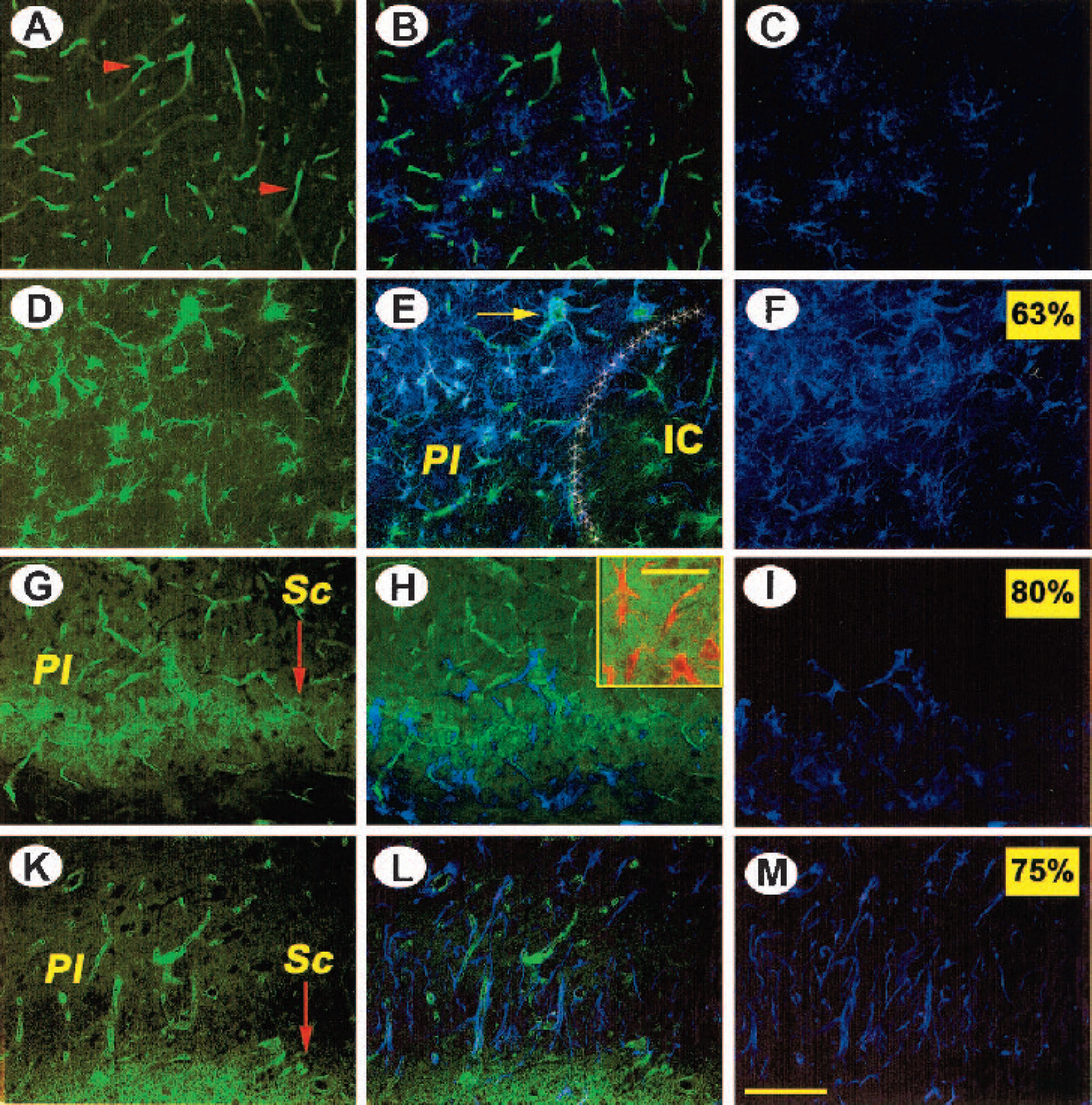
Development of scar tissue after cerebral ischemia in aged rats. Double immunofluorescence for the endothelial cell marker, RECA (green), and the astrocytic marker, GFAP (violet). Note the thin, short blood vessels intermingled with astrocytes in layers II to III of aged (unlesioned) rats (
Time course of chondroitin sulfate proteoglycan expression
Three days after stroke, NG2 immunostaining showed strong activation of oligodendrocytes, which otherwise are barely visible under our experimental conditions, both in young and aged rats (Figs. 6A and 6B, green). In the infarct core at one week post-lesion, scattered NG2 immunoreactivity (Fig. 6C) was intermingled with moderately activated astrocytes (Fig. 6C, red) as well as nuclei that were most likely of hematogenous origin (Fig. 6C, blue). In aged rats, activated oligodendrocytes displaying NG2 immunoreactivity (Fig. 6D) persisted in the outermost scar region at 1 to 2 weeks. Highly activated astrocytes (red) penetrated the outermost scar region (Fig. 6D, arrow). At 4 weeks, the image from young rats was reminiscent of that of aged rats (Fig. 6E) at 2 weeks, while in aged rats we noted a decreased astroglial reaction in the scar region on a chondroitin sulfate background (Fig. 6F).
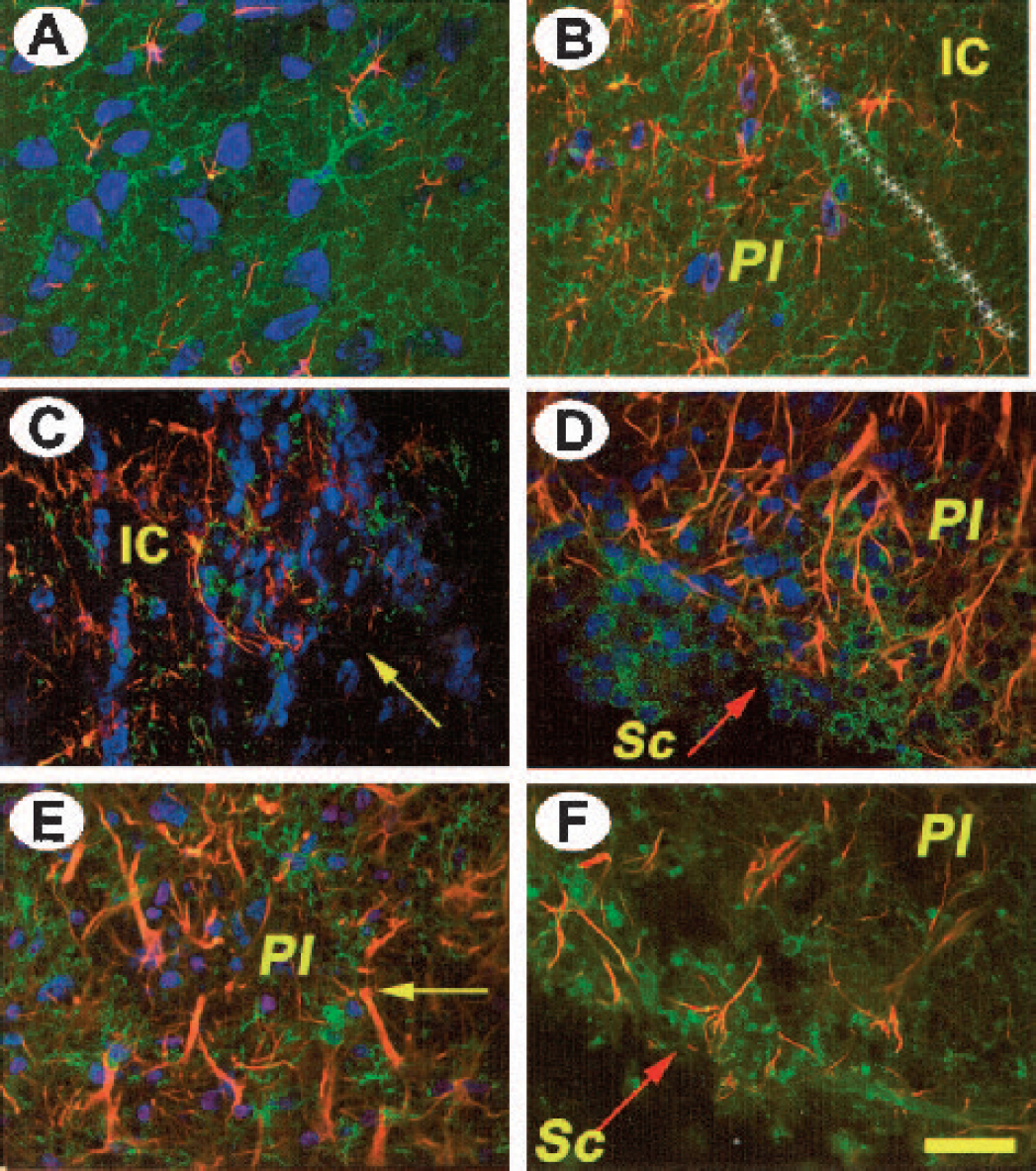
Development of oligodendrocytic activation after cerebral ischemia. Nuclei are shown in blue, astrocytes in red, and oligodendrocytes in green. Note the strong oligodendrocytic reaction at 3 days after stroke in both young (
DISCUSSION
Aging is a major risk factor for stroke, but how aging affects the response of the brain to insult is poorly understood. The influence of aging on functional recovery is complex (Shapira et al., 2002). In this study, we tested the hypothesis that a temporally abnormal glial reaction in the brains of aged rats may impair functional recovery after stroke. Our findings show that astrocytic, microglial, and oligodendrocytic reactivity peaks very early after transient ischemic insult in aged rats, resulting in the premature formation of scar-like tissue and correlating with diminished recovery.
Cerebral ischemia produces a necrotic core in which the blood–brain barrier is disrupted. At a cellular level, focal cerebral ischemia elicits a strong inflammatory reaction initiated by the activation of astrocytes, resident microglia, and perivascular macrophages. After transient ischemia, activated microglia express numerous antigens, including CD4 and major histocompatibility complex class I and II antigens, as well as phagolysosomes recognized by the monoclonal antibody ED1. In young animals, microglia in the periinfarct area evolve into a phagocytosing phenotype after 3 to 7 days (Gehrmann et al., 1992; Kato et al., 1999; Lehrmann et al., 1997). In this model, reactive astrogliosis takes 2 to 4 days to develop and is associated with the induction of many proteins with potent biological effects, such as transforming growth factor (TGF) β, β-amyloid precursor protein, apolipoprotein E, and lipocortin (Clarke et al., 1993; Norenberg, 1994). Thereafter the number of reactive astrocytes and microglia decreases in the lesion core but persists up to 7 days at the boundary zone (Chen et al., 1993). Between days 7 and 14, the number of ramified microglia and astrocytes greatly declines in the outer boundary of the lesion. Our results emphasize that the time course of glial activation is strongly age dependent.
Although the extent of neuronal degeneration in the penumbra tended to be most pronounced in young rats, histologic staining consistently has shown that the extent of neuronal loss and infarct size are comparable in young and aged rats after cerebral ischemia (Popa-Wagner et al., 1998, 1999; Shapira et al., 2002), suggesting that the necrotic focus is larger in aged rats. Greater postischemic tissue damage could contribute to a stronger and more precipitous glial reaction in aged rats.
There is reason to suspect that glial cells might respond differently to ischemia in the aged brain, since earlier studies have shown that microglia and astrocytes increase both in number and reactivity in normal older subjects. For example, the levels of GFAP and its messenger ribonucleic acid increase in the astrocytes of rats as a function of age (Goss and Morgan, 1995; Nichols et al., 1995), and microglial cells also display an age-associated augmentation of reactivity in a variety of mammalian species (Goss and Morgan, 1995; Jucker et al, 1994; Nichols et al., 1995; Peters, 2002; Vogelgesang et al., 2002; Yu et al., 2002).
Generally there is an increased astrocytic reaction following injury to the aging CNS. For example, Gordon et al. (1997) found that nigrostriatal deafferentation causes a time-dependent induction of GFAP that is larger and more persistent in aged rats than in young animals. Interestingly, the authors noted an intensification of GFAP staining in the contralateral hemisphere of aged (but not young) rats, a phenomenon that was confirmed by our findings. We also found that tissue morphology at 1 to 2 weeks after stroke in aged rats is similar to that at 4 weeks in young rats; i.e., in both instances we noted a scar-like formation, fully penetrated by reactive astrocytes. We also noted a strong activation of oligodendrocytes at early stages of infarct development in both young and aged rats that persisted in older rats, suggesting the formation of an even stronger barrier to regeneration-like phenomena.
Intriguingly, a colocalization of RECA-like immunoreactivity with GFAP-like immunoreactivity was noted in aged rats shortly after the ischemic event. It is conceivable that RECA-like immunostaining is due to angioblastlike circulating endothelial cell progenitors that are present in adult animals (Asahara et al., 1997; Rafii, 2000). Another possibility is that circulating endothelial progenitor cells from bone marrow have leaked into the brain and contribute to the RECA immunopositivity (Zhang et al., 2002). Still another possibility is that activated astrocytes, which co-localize with cerebral blood vessels 7 to 28 days after ischemia, exhibit antigens specific for neuropilin-1 that are normally expressed by endothelial cells (Zhang et al., 2001).
The early formation of the glial scar may impede functional recovery in aged rats. The temporal correlation between an increase in glial activity after stroke in aged rats and the decline in function lends support to this hypothesis. Interestingly, before surgery the aged rats performed as well as young rats on most of the sensorimotor tests (except the spontaneous activity test and, to a lesser extent the inclined plane test). In more dynamic tests like Rotarod, the relatively good performance of aged rats could partly be attributable to the beneficial effects of training prior to surgery. After the experimental ischemic episode, however, young animals generally recovered more rapidly and to a greater extent than did aged rats. A peculiar finding was that performance in the inclined plane test decreased in aged rats 3 days after ischemia and never recovered to prestroke levels.
Depending on the time course of their expression and the type of lesion, reactive astrocytes may play a dual role in tissue remodeling after injuries. Recent studies have suggested that reactive astrocytes secrete neurotrophic factors at the lesion site in response to injuries (Clarke et al., 2001; Miyazaki et al., 2001) and may provide a permissive substrate for axonal regrowth (Ridet et al., 1997). However, at later stages, a scar-specific astrocytic phenotype that may be an obstacle to axonal regrowth can emerge (Asher et al., 2002; Chen et al., 2002; Morgenstern et al., 2002; Niederost et al., 1999). This latter view has engendered several possible strategies that attempt to diminish the extent of scar formation after cerebral ischemia.
For example, it has been shown that TGF-β is a potent reducer of the fibrotic scar by reducing the levels of laminin, fibronectin, and chondroitin sulphate proteoglycan (Logan et al., 1999). Likewise, scar formation after injury could be reduced by the action of matrix metalloproteinases 2 and 9 (Duchossoy et al., 2001), or by neutralizing the action of the proteoglycan neurocan with antineurocan antibodies (Chen et al., 2002). Recently it has been recognized that the control of the glial reaction is likely to be an important therapeutic target. Thus, treatment with TGF-β1 and TGF-β2 resulted in a reduction of scar formation (Moon and Fawcett, 2001). Likewise, treatment with 2-propyloctanoic acid significantly reduced the expression of S-100β and glial fibrillary acidic protein in activated astrocytes in the periinfarct area, concomitant with a significant attenuation of neurologic deficits (Tateishi et al., 2002). Another therapeutic possibility would be to diminish the astrocytic response to stroke in aged rats by blocking the L-type calcium channels with verapamil (Klepper et al., 1995) or by using the immunosuppressant agent cyclosporin (Wakita et al., 1995). Our finding that the glial reaction in the aged brain is inappropriately accelerated supports the view that modulation of glial function might improve the prognosis for recovery from stroke and other forms of brain injury in the elderly.