
Abstract
Adenovirus-mediated gene transfer to blood vessels is relatively inefficient because binding of adenovirus to vessels is limited. The authors have reported that incorporation of cationic polymer and lipids with adenovirus augments gene transfer to blood vessels ex vivo. In this study, the authors determined whether complexes of adenovirus and cations improve efficiency of gene transfer in vivo. Poly-l-lysine, lipofectamine, or lipofectin was complexed with adenovirus encoding β-galactosidase. Optimum ratios of the cations per adenovirus were determined by gene transfer to fibroblasts. After injection of the adenovirus into the cisterna magna of anesthetized rabbits, transgene activity was greater in the adventitia of intracranial arteries and meninges after injection of the complexes than adenovirus alone. Thirty minutes after application of adenovirus with the cations, binding of adenovirus to fibroblast cells in vitro or the basilar artery in vivo (by Southern blot analysis) was augmented, which suggests that enhanced binding of virus contributes to augmentation of transgene expression. Thus, cationic polymer and lipids improve transgene expression in intracranial arteries, primarily in the adventitia, after adenovirus-mediated gene transfer in vivo. This strategy may be applicable to studies of gene transfer and eventually for gene therapy.
Vascular gene transfer has made rapid progress since the first report 10 years ago (Nabel et al., 1990), and clinical trials for patients with coronary and peripheral vascular diseases are already under way (Baumgartner et al., 1998; Mann et al., 1999). In contrast, there are far fewer studies of gene transfer to cerebral blood vessels in vivo, even in animal models. A reason for this lag may be that intravascular gene delivery, which is feasible for coronary and peripheral arteries, would require interruption of blood flow to the brain if applied to cerebral arteries (Heistad and Faraci, 1996). As an alternative approach, the authors and others have used perivascular delivery of genetic vectors to cerebral arteries through cerebrospinal fluid (CSF) (Ooboshi et al., 1995; Christenson et al., 1998). With this approach, adventitia is the main site for transgene expression in vessels.
Adenoviral vectors encoding vasoactive enzymes and peptides modulate vasomotor tone of cerebral arteries after perivascular delivery (Chen et al., 1997; Toyoda et al., 2000b). Absence of adenoviral fiber receptor activity in fibroblast cells, however, limits viral binding to the adventitia, and thus expression of transgene products (Seth et al., 1994; Wickham et al., 1996).
Incorporation of adenoviral vectors with nonviral molecules including a cationic polymer (Fasbender et al., 1997), viscous polyol (Feldman et al., 1997), lipids (Fasbender et al., 1997), and calcium phosphate precipitates (Fasbender et al., 1998) improves viral binding to target cells and increases expression of transgene products. The nonviral molecules are generally inexpensive, and easy to use by mixing with virus to form a noncovalent complex. The authors have reported cationic polymer and lipid augment efficiency of virus-mediated gene transfer to carotid arteries ex vivo (Toyoda et al., 1998). Transgene expression was greatly increased in endothelium, but the effect of the complexes on gene transfer to the adventitia was not clearly demonstrated. That is important because adventitia is the major site of transgene expression within the vessel wall after gene transfer to cerebral arteries using perivascular delivery through CSF (Ooboshi et al., 1995; Christenson et al., 1998). Thus, it was not clear whether cations would be useful for augmentation of adenovirus-mediated gene transfer in vivo through CSF.
The first goal of this study was to determine, in studies in vitro, the optimal conditions of complexes of adenovirus with cationic polymer and lipids for gene transfer to fibroblasts. The second goal was to determine whether the complexes augment efficiency of perivascular gene transfer to cerebral arteries in vivo. The authors also determined effects of the complexes on gene transfer to the surface of the brain including meninges, an important target for gene therapy to brain parenchyma (Betz et al., 1995). The third goal was to clarify how gene transfer was augmented after using the complexes. The authors hypothesized that viral binding to target cells soon after injection of the virus is the major factor to regulate efficiency of transgene expression, and used Southern blot analysis to compare amounts of bound virus to cells with and without cationic polymer and lipids.
MATERIALS AND METHODS
Adenovirus-cation complexes
Recombinant adenovirus vector of serotype 5 encoding nuclear-targeted β-galactosidase expressed from a long terminal repeat of Rous sarcoma virus as a promoter (AdRSVßgal) was generated using standard methods by the University of Iowa Gene Transfer Vector Core (Iowa City, IA, U.S.A.) at titers of 3 × 1010 plaque-forming units (pfu)/mL (1 × 1012 particles/mL) (Toyoda et al., 2000b). Polymers of poly-l-lysine hydrochloride with an average molecular mass of 37.0 kDa (Sigma), lipofectamine (Gibco) formulated in a 3:1 weight ratio of the polycationic lipid 2,3-dioleyloxy-N-[2(spermine-carboxamido)ethyl]-N,N-dimethyl-l-propanaminiumtrifluoroacetate (DOSPA) to the neutral lipid dioleoyl phosphatidylethanolamine (DOPE), and lipofectin (Gibco) formulated in a 1:1 weight ratio of the cationic lipid N-[1-(2,3-dioleyloxy)propyl]-N,N,N-trimethylammonium chloride (DOTMA) to DOPE were used as cationic molecules.
Complexes of adenovirus and cations were made by adding viral vectors and cationic agents to phosphate-buffered saline (PBS), mixing by gentle pipette tip aspiration, and incubation for 30 minutes at room temperature before application to cells or animals (Fasbender et al., 1997; Toyoda et al., 1998). The ratio of cations was expressed as a calculated average number of molecules of poly-l-lysine, DOSPA, or DOTMA per adenovirus particle.
Gene transfer to cultured cells
NIH-3T3, a fibroblast cell line, was cultured on 35-mm dishes in Dulbecco's modified Eagle's medium (DMEM, high glucose) supplemented with 10% fetal bovine serum, 100 U/mL penicillin, and 100 μg/mL streptomycin. Cells were seeded at 3 × 104/cm2, and infected 18 to 24 hours after seeding when cells were ≈70% confluent. The medium was replaced with 1 mL of DMEM containing 5 × 109 particles of adenovirus (multiplicity of infection ≈100) with or without cations, each cation alone, or vehicle (PBS). Cells were incubated in a 5% CO2 humidified atmosphere at 37°C unless otherwise noted. The infection solution was removed after 2 hours, fresh serum-containing DMEM was added, and incubation continued for 48 hours.
Gene transfer to rabbits in vivo
All animal procedures were approved by the Animal Care and Use Review Committee at the University of Iowa. Male New Zealand White rabbits weighing 2.4 to 3.0 kg were anesthetized with 5 mg/kg xylazine intramuscularly and 40 mg/kg ketamine intramuscularly. The authors infused 250 μL of suspension of incorporation of 1010 particles of AdRSVßgal with or without cations, each cation alone, or vehicle (PBS) into the cisterna magna as previously described (Toyoda et al., 2000b). Because the total volume of freely flowing CSF in the rabbit is 2 to 3 mL, the final viral titer in CSF was estimated to be 1 to 1.5 × 108 pfu/mL.
Five days after injection, unless otherwise noted, rabbits were anesthetized with 50 mg/kg pentobarbital intravenously, exsanguinated, and the brain stem was removed. For AdRSVßgal, 5 days duration is optimal for expression of transgene products both in cerebral arteries (Toyoda et al., 2000c) and brain surface (Betz et al., 1995). The basilar artery, meninges, choroid plexus in the fourth ventricle, and brain stem tissue were used for assays.
Assays for transgene expression
β-Galactosidase activity was determined using chemiluminescent assay as described previously (Toyoda et al., 1998, 2000b). Activity was assayed in 3T3 cells 48 hours after viral application. Forty-eight hours seems to be the maximum time to perform experiments with little damage of 3T3 cells (Toyoda et al., 2000c). Activity was assayed in the basilar artery (3-mm-long), meninges over the ventral brain stem, choroid plexus in the fourth ventricle, and ventral half of brain stem tissue from the proximal to distal end of the basilar artery 5 days after viral application in vivo. Five days are optimum duration to obtain maximal expression of transgene products in vivo (Toyoda et al., 2000c).
Histochemical staining for detection of β-galactosidase also was used to examine localization of transgene in basilar arteries after gene transfer in vivo as described previously (Ooboshi et al., 1995; Toyoda et al., 2000b).
Assays for adenoviral binding
To examine adenoviral binding to target cells, 3T3 cells were exposed to 5 × 109 particles (1.5 × 108 pfu) of AdRSVßgal (multiplicity of infection 100) with or without cationic molecules at 4°C, at which temperature cells do not internalize adenovirus (Toyoda et al., 1998). Thirty minutes after exposure, adenoviral DNA was isolated from cells and quantified using Southern blot analysis as described previously (Toyoda et al., 1998, 2000a). Autoradiographs were scanned by QCS3200 flatbed scanner (Imapro) and analyzed for density of bands of adenoviral DNA (area × mean) with Volume Trace Motif version 1.21 (University of Iowa Image Analysis Facility). Proportion of adenoviral binding to applied AdRSVßgal was calculated compared with the density of standard bands (Toyoda et al., 1998, 2000a). The authors also examined the basilar artery after injection of virus in vivo, to determine adenoviral association (both viral binding and internalization into the artery). Rabbits were euthanized 30 minutes after injection of 1010 particles (3 × 108 pfu) of AdRSVßgal with or without cations into the cisterna magna, and viral DNA was quantified from the basilar artery using Southern blot analysis. Because virus contacts the artery at ≈39°C in vivo, the authors could not exclude the possibility that virus is internalized in part in the artery.
Statistical analysis
Values are expressed as mean ± SEM. One-way factorial analysis of variance followed by Bonferroni test was used for comparison of variables among groups. An unpaired t-test was used for comparison of β-galactosidase activity after gene transfer in vivo. P < 0.05 was accepted as statistically significant.
RESULTS
Transgene expression in cell cultures
First, optimal conditions of the complexes of adenovirus with cationic polymer and lipids by gene transfer to fibroblasts in culture were determined. There was minimal β-galactosidase activity after exposure to AdRSVßgal alone (Fig. 1). Incorporation of AdRSVßgal with 30 to 3000 molecules of poly-l-lysine per virus particle greatly increased the activity, with the maximal increase (43-fold) by 300 molecules per particle. The activity also was significantly increased by 3 × 103 to 3 × 105 molecules of lipofectamine per virus particle with the maximal increase (23-fold) by 3 × 104 molecules per particle, and by 3 × 103 to 105 molecules of lipofectin per virus particle, with the maximal increase (7-fold) by 3 × 104 molecules per particle. Thus, complexes of adenovirus with cationic polymer and lipids augmented gene transfer to fibroblast cells.
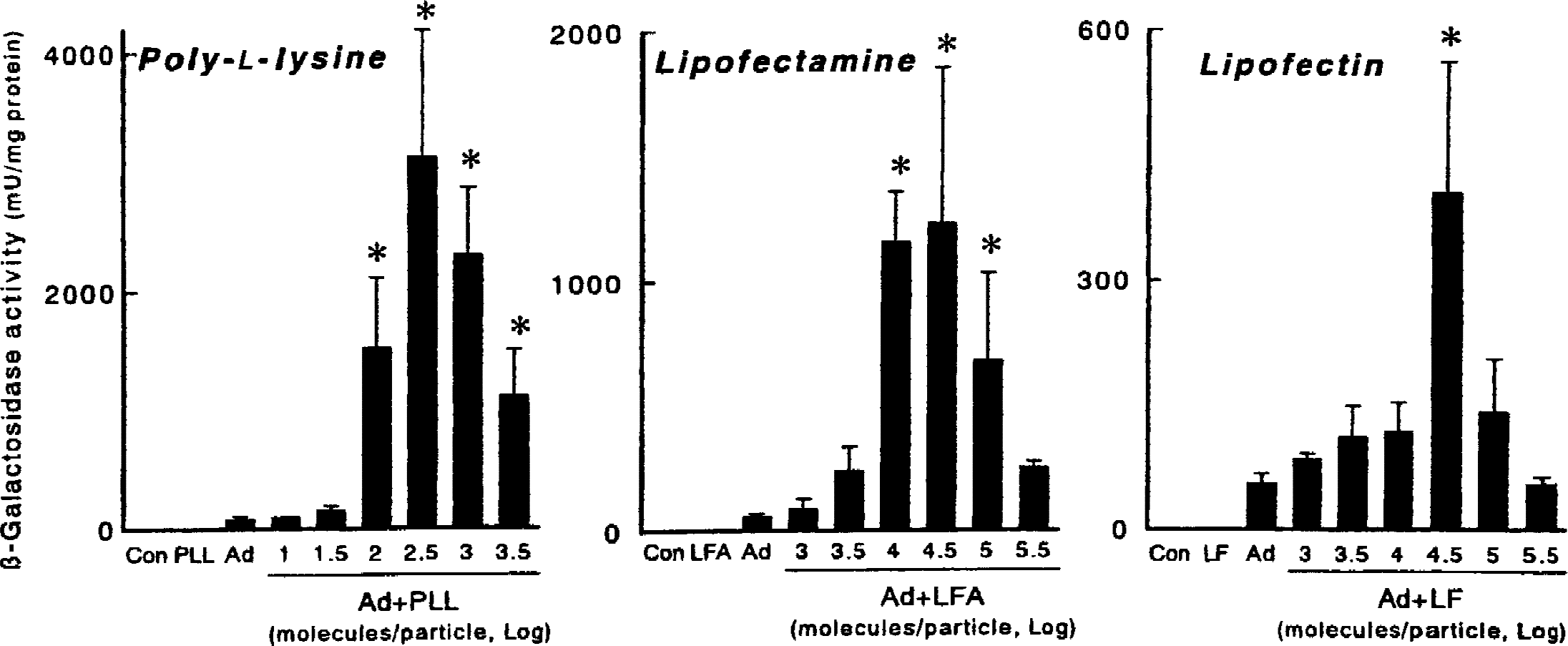
β-Galactosidase activity in NIH-3T3 cells 48 hours after application of AdRSVßgal (100 multiplicity of infection) with poly-l-lysine, lipofectamine, and lipofectin. Con, control (no viral application); PL, poly-l-lysine alone (same dose as 300 molecules/virus particle); LFA, lipofectamine alone (same dose as 3 × 104 molecules/virus particle); LF, lipofectin alone (same dose as 3 × 104 molecules/virus particle); Ad, AdRSVßgal alone. Values are mean ± SD (n = 6 for each group). * P < 0.005 versus Ad. Note that the scales are different in the three panels.
Transgene expression after gene transfer in vivo
Next, the authors studied effects of cationic molecules on adenovirus-mediated gene transfer to the brain in vivo. Based on findings in 3T3 cells, 100 molecules of poly-l-lysine per virus particle and 3 × 104 molecules of lipofectamine per particle were used. Lipofectin produced smaller effects on gene transfer to 3T3 cells than lipofectamine, and thus was not used for studies in vivo.
All rabbits tolerated injection of the coprecipitates without detectable neurologic deficits or alteration of rectal temperature (39.3°C ± 0.2°C before injection and 39.1°C ± 0.1°C 5 days after injection). Five days after injection of virus into CSF of rabbits, poly-l-lysine significantly increased β-galactosidase activity in the basilar artery by 40-fold, in the meninges by 38-fold, in the choroid plexus by 13-fold, and in the brain stem by 13-fold, relative to AdRSVßgal alone (Fig. 2). Lipofectamine significantly increased the activity in the basilar artery by 12-fold, in the meninges by 17-fold, in the choroid plexus by 17-fold, and in the brain stem by 12-fold, relative to AdRSVßgal alone. Poly-l-lysine increased the activity more than lipofectamine in the basilar artery and meninges (P < 0.02).
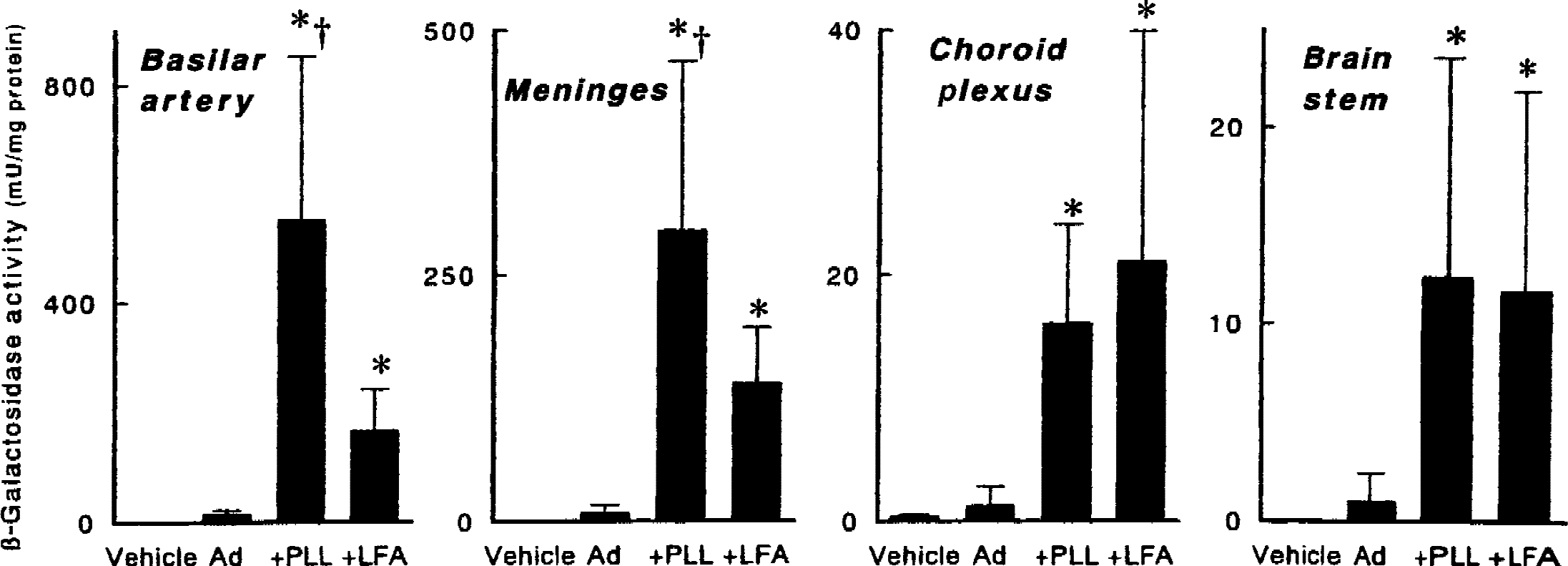
β-Galactosidase activity in basilar arteries, meninges over the brain stem, choroid plexus in the fourth ventricle, and brain stem tissues from rabbits 5 days after injection into the cisterna magna in vivo of vehicle, AdRSVßgal alone (Ad, 1010 particles), AdRSVßgal with 100 molecules of poly-l-lysine per adenovirus particle (+PLL), or AdRSVßgal with 3 × 104 molecules of lipofectamine per adenovirus particle (+LFA). Values are mean ± SD (n = 2 rabbits for vehicle, and 6 for others). * P < 0.02 versus Ad. †P < 0.02 versus +LFA. Note that the scales are different in the four panels.
By histochemistry of the basilar artery, augmentation of transgene expression was demonstrated in the adventitia after injection of AdRSVßgal with poly-l-lysine and lipofectamine compared with after AdRSVßgal alone (Fig. 3). From cross-sections, β-galactosidase was not detected in the medial layer or endothelium. Thus, complexes of adenovirus with cationic molecules augmented gene transfer in vivo to the adventitia of cerebral arteries and surrounding meninges.
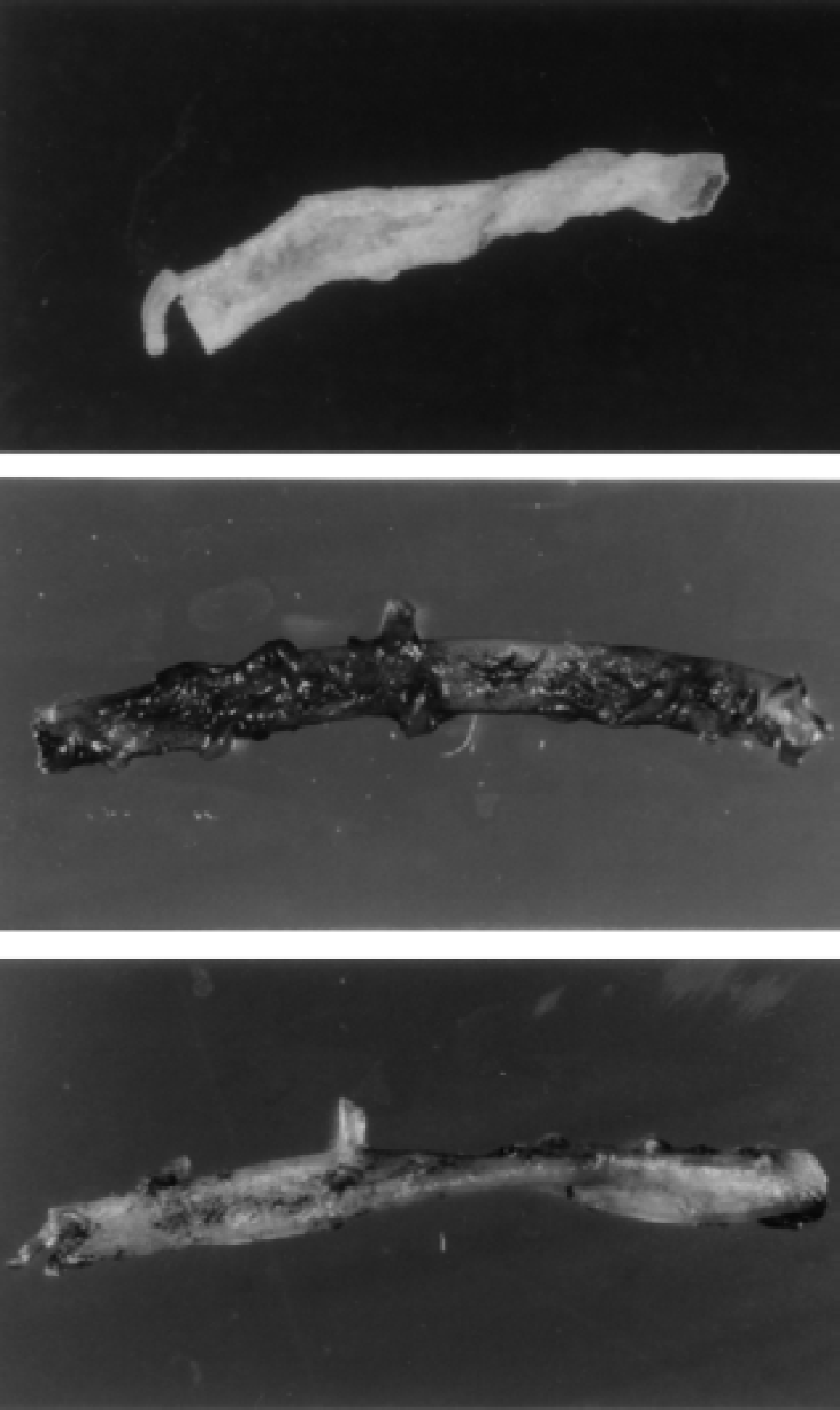
Histochemical localization of β-galactosidase in rabbit basilar arteries 5 days after injection into the cisterna magna in vivo of AdRSVßgal alone
Adenoviral binding
As a mechanism for augmented adenovirus-mediated gene transfer by cationic polymer and lipids, the authors tested the hypothesis that binding of adenovirus to fibroblasts and cerebral arteries was improved by cationic molecules. Thirty minutes after exposure of 3T3 cells to complexes of AdRSVßgal with 100 molecules of poly-l-lysine per virus particle, adenoviral DNA bound to cells was 5.1% ± 0.5% of all adenovirus used, which was 9-fold greater than viral DNA after AdRSVßgal alone (0.5% ± 0.1%, n = 6 for both groups;P < 0.0001; Fig. 4). Viral DNA bound to cells after exposure to AdRSVßgal with 3 × 104 molecules of lipofectamine per virus particle was 16-fold greater than after AdRSVßgal alone (n = 6, P < 0.0001). Viral DNA bound to cells after exposure to AdRSVßgal with 3 × 104 molecules of lipofectin per virus particle was 10-fold greater than viral DNA after AdRSVßgal alone (n = 6, P < 0.0005).
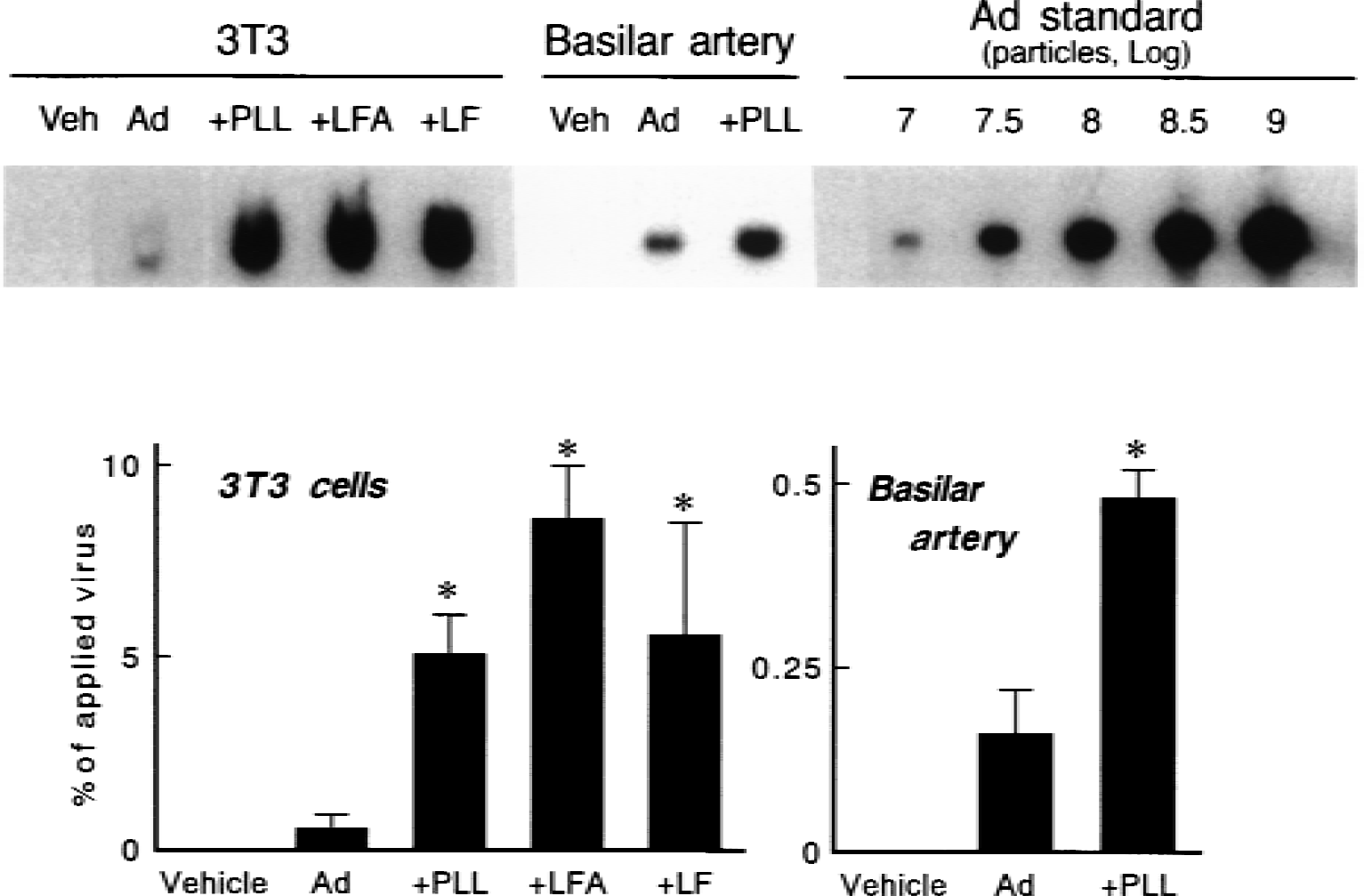
Adenoviral DNA bound to NIH-3T3 cells and basilar arteries from rabbits 30 minutes after exposure to vehicle, AdRSVßgal alone (Ad), AdRSVßgal with 100 molecules of poly-l-lysine per adenovirus particle (+PLL), AdRSVßgal with 3 × 104 molecules of lipofectamine per adenovirus particle (+LFA), and AdRSVßgal with 3 × 104 molecules of lipofectin per adenovirus particle (+LF).
The authors also measured adenoviral DNA associated with the basilar artery 30 minutes after injection of AdRSVßgal with 100 molecules of poly-l-lysine per virus particle in vivo. Viral DNA was 3-fold greater after AdRSVßgal with poly-l-lysine than viral DNA after AdRSVßgal alone (0.49% ± 0.02% vs. 0.16% ± 0.02%, n = 3 for both groups;P < 0.0005; Fig. 4). The authors did not measure viral DNA associated with the basilar artery after adenovirus with lipofectamine.
DISCUSSION
The major new findings in this study include the following: first, that the incorporation of adenovirus with cationic polymer and lipids augments transgene expression in fibroblasts in culture. Second, this incorporation augments transgene expression in the adventitia of the basilar artery and surrounding brain surface tissues after gene transfer in vivo. Third, the augmentation seems to result, at least partly, from enhanced adenoviral binding to the tissues. Thus, cationic polymer and lipids are useful for augmentation of adenovirus-mediated gene delivery to the brain through CSF in vivo.
Adenovirus binds to cells through its fiber coat protein, followed by internalization into cells through its penton base coat protein (Kovesdi et al., 1997). Two unrelated cellular receptors for the fiber have been identified, Coxsackie B and adenovirus receptor (CAR) (Bergelson et al., 1997) and the α2 domain of MHC Class I (Hong et al., 1997). In human airway epithelial cells, resistance to adenovirus infection is caused by absence of CAR and MHC Class I at the apical surface (Walters et al., 1999). Although the number of CAR or MHC Class I in blood vessels is not known, the fiber receptor activity in the adventitia appears to be low (Hidaka et al., 1999). Hence, binding and subsequent internalization of adenovirus into the adventitial cells under normal conditions may be limited.
Cationic polymers and lipids are useful for nonviral gene transfer, which incorporate with plasmid DNA or oligonucleotides and deliver the nucleic acids to cells through fusion with cell membranes or receptor-mediated endocytosis (Felgner et al., 1987; Simons et al., 1992). Among them, lipofectamine (Muller et al., 1994) and lipofectin (Nabel et al., 1992) were reported as vectors for gene transfer in vivo to blood vessels. Complexes of plasmid DNA with the cationic polymers and lipids are more potent by coupling to inactivated adenovirus or herpes virus of Japan (Wagner et al., 1991; Kaneda et al., 1989). Once inside cells, viral proteins escape from the endosome and enter into the nucleus (Greber et al., 1993) and enhance gene transfer. Here, the authors incorporated cationic polymers and lipids with recombinant adenovirus which encodes transgene in adenovirus DNA. In this strategy, because adenovirus is not inactivated and covalent linkage between virus and cations is not used, there may be less danger of inactivating important viral functions. The same strategy was used for gene transfer to the airway epithelium (Fasbender et al., 1997).
Among the three cations examined, poly-l-lysine seems to be best for augmentation of adenoviral gene transfer to fibroblast cells both in vitro and in vivo (the basilar artery and meninges). Increased adenovirus DNA in cell culture and the basilar artery suggest that cations enhance viral binding soon (30 minutes) after application of the virus. Poly-l-lysine was more effective than lipofectamine in increasing transgene expression, but not in viral binding to the fibroblast cells. Thus, at least for poly-l-lysine, there may be mechanisms in addition to improvement of viral binding that contribute to augmentation of adenoviral gene transfer. Poly-l-lysine does not appear to be more effective than lipofectamine for adenoviral gene transfer to choroid plexus or brainstem.
In the current study, an excess ratio of poly-l-lysine and lipids did not improve transgene expression in fibroblast cell culture. A possible explanation is that a high ratio of cations to adenovirus particle appears to produce aggregation of viral particles and prevent efficient viral infection (Fasbender et al., 1997). A second possibility is that extremely high concentrations of cationic polymer or lipids are toxic to cells. Lee et al. (1999) described that 7,500 ng poly-l-lysine (corresponding to ≈30.000 molecules per adenovirus particle in this study) or 100 μg lipofectin (corresponding to ≈107 molecules per particle) with adenovirus increased death of cells in cultures. Hence, it was important to determine an appropriate ratio of poly-l-lysine and lipids to adenovirus particle to be used for gene transfer in vivo, based on findings in cell cultures. Under conditions in vivo, animals did not develop neurologic or systemic signs after injection of optimal dose of the cationic polymer and lipids together with adenovirus in the current and previous studies using mice (Fasbender et al., 1997). Function of isolated arteries ex vivo was not impaired after exposure to the adenovirus-cations complexes (Toyoda et al., 1998). Thus, the dose of the polymer and lipids in this study seems to be safe in vivo.
The authors speculate that improvement of adenovirus-mediated gene transfer to blood vessels using the cationic molecules may potentially be useful for treatment of cerebrovascular diseases. In the authors' previous study in vitro, increased expression of endothelial nitric oxide synthase in the isolated carotid artery using adenovirus-cations complexes contributed to augmentation of nitric oxide-mediated vascular function (Toyoda et al., 1998). Thus, it was possible to obtain greater vasoactive effects of transgene products after gene transfer of adenovirus together with cationic polymer and lipids than adenovirus alone. Alternatively, it may be possible to reduce the amounts of virus or time of viral exposure using the adenovirus-cations complexes to obtain the same efficiency of gene transfer to cerebral arteries and to reduce the risk of immune responses mediated by viral infection.
Gene therapy using viral vectors can prevent vasospasm after experimental subarachnoid hemorrhage, and may be applicable to treatment of human subarachnoid hemorrhage (Toyoda et al., 2000c). In addition, it is possible that gene therapy may eventually be used to treat some neurologic diseases, including neurodegeneration, brain ischemia, and brain injury (Davidson and Bohn, 1997). Gene delivery to the brain surface and surrounding CSF may potentially alter pathophysiologic mechanisms in brain parenchyma. For example, injection of adenovirus encoding interleukin-1 receptor antagonist through CSF reduced brain edema and infarct volume both in cortex and basal ganglia after focal brain ischemia (Betz et al., 1995). Because cationic polymers and lipids improve expression of transgene products in the brain tissue and meninges, the cations may be applicable to adenovirus-mediated gene therapy of neurologic diseases other than cerebrovascular diseases.
Footnotes
Acknowledgments:
The authors thank Pamela K. Tompkins and Sonya Mehta for technical assistance and Arlinda LaRose for typing the manuscript. The authors also thank Dr. Frank M. Faraci for assistance with the manuscript, and Dr. Beverly L. Davidson, Richard D. Anderson, and the University of Iowa Gene Transfer Vector Core for preparation of virus.