
Abstract
In vertebrates, the interconversion of lactate and pyruvate is catalyzed by the enzyme lactate dehydrogenase. Two distinct subunits combine to form the five tetrameric isoenzymes of lactate dehydrogenase. The LDH-5 subunit (muscle type) has higher maximal velocity (Vmax) and is present in glycolytic tissues, favoring the formation of lactate from pyruvate. The LDH-1 subunit (heart type) is inhibited by pyruvate and therefore preferentially drives the reaction toward the production of pyruvate. There is mounting evidence indicating that during activation the brain resorts to the transient glycolytic processing of glucose. Indeed, transient lactate formation during physiological stimulation has been shown by 1H-magnetic resonance spectroscopy. However, since whole-brain arteriovenous studies under basal conditions indicate a virtually complete oxidation of glucose, the vast proportion of the lactate transiently formed during activation is likely to be oxidized. These in vivo data suggest that lactate may be formed in certain cells and oxidized in others. We therefore set out to determine whether the two isoforms of lactate dehydrogenase are localized to selective cell types in the human brain. We report here the production and characterization of two rat antisera, specific for the LDH-5 and LDH-1 subunits of lactate dehydrogenase, respectively. Immunohistochemical, immunodot, and western-blot analyses show that these antisera specifically recognize their homologous antigens. Immunohistochemistry on 10 control cases demonstrated a differential cellular distribution between both subunits in the hippocampus and occipital cortex: neurons are exclusively stained with the anti-LDH1 subunit while astrocytes are stained by both antibodies. These observations support the notion of a regulated lactate flux between astrocytes and neurons.
Keywords
Determination of arteriovenous differences for metabolic intermediates of the brain have clearly shown that glucose is virtually the exclusive metabolic substrate of the brain and that it is almost entirely oxidized to water and carbon dioxide (Siesjö, 1978). However, clear regional differences in glucose utilization exist in register with neuronal activity (Sokoloff, 1981). Interestingly, during activation of specific neuronal pathways, glucose utilization, as monitored with the 2-deoxyglucose autoradiographic technique, increases in the neuropil, i.e., in regions enriched in dendrites, axons, and the astrocytic processes that ensheath synapses, not in the cell body (Kadekaro et al., 1985). These observations have raised the possibility that during activation astrocytes could contribute to 2-deoxyglucose uptake and phosphorylation. Recently, evidence has been provided in primary cultures of astrocytes and neurons (Pellerin and Magistretti, 1994, Pellerin et al., 1995) and in noncultured guinea pig retina (Poitry-Yamate and Tsacopoulos, 1992) that support this view. In the guinea pig retina, glucose is taken up during activation by glial cells, which then process it to products of glycolysis such as lactate (Poitry-Yamate et al., 1995). In primary astrocyte cultures, application of glutamate, which mimics activation, stimulates glucose utilization and lactate production (Pellerin and Magistretti, 1994). Stimulation of glucose uptake and phosphorylation by glutamate has recently been confirmed in rat astrocytes (Takahashi et al., 1995). In view of these observations, and of the growing evidence that lactate can be an adequate substrate for neurons to maintain synaptic activity (Schurr and Rigor, 1988), we have formulated the hypothesis of a metabolic cellular compartmentation whereby during activation glucose is also taken up by astrocytes, which process it glycolytically to lactate for the use of neurons (Pellerin and Magistretti, 1994). Indeed, the cytological relationships of astrocytes strongly support this view: astrocytic end-feet surround virtually all brain capillaries (Andriezen, 1893; Cajal, 1909; Peters et al., 1991), indicating a likely role in the transit of metabolic substrates originating from the circulation, while other astrocyte processes ensheath synapses and possess receptors as well as uptake sites for neurotransmitters (Barres, 1991). These features make astrocytes ideally geared to modulate uptake of substrates from the circulation and “sense” synaptic activity; in other words, astrocytes are well positioned to couple neuronal activity to energy metabolism (Magistretti et al., 1995; Tsacopoulos and Magistretti, 1996).
As lactate emerges as an important energy metabolite for brain cells, it follows that the cellular localization of lactate dehydrogenase (LDH, EC 1.1.1.27), the rate-limiting enzyme in lactate fluxes, deserves further attention. In vertebrates, the five isoenzymes of LDH are a tetrameric combination of two different polypeptide chains (Markert and Appella, 1961), the A (muscle or M) subunit that we will call here the LDH-5 subunit, and the B (heart or H) subunit that we will term the LDH-1 subunit. The different catalytic properties of the five isoenzymes of LDH (B4 or LDH-1, B3A or LDH-2, B2A2 or LDH-3, BA3 or LDH-4, and A4 or LDH-5) are in proportion to the ratio of LDH-5 to LDH-1 subunits (Cann et al., 1962; Dawson et al., 1964; Bishop et al., 1972). Furthermore, the relative distribution of LDH-5 and LDH-1 subunits is tissue-specific (Plagemann et al., 1960; Wroblewski et al, 1960). The LDH-5 isozyme is predominantly found in lactate-producing tissues, while the LDH-1 isozyme is active in aerobic tissues (e.g., the heart) (Cahn et al., 1962; Markert et al, 1975). It has been proposed (Cahn et al., 1962) and generally accepted (Pevzner, 1983) that the LDH-1 isoform is better designed to oxidize lactate to pyruvate, while the LDH-5 isoform preferentially converts pyruvate to lactate (Cahn et al., 1962).
In view of the foregoing considerations, we have produced and characterized specific antibodies against both types of subunits, and we have used them in two regions of the adult human brain to reveal the cellular localization of each subunit.
MATERIALS AND METHODS
Preparation of tissues
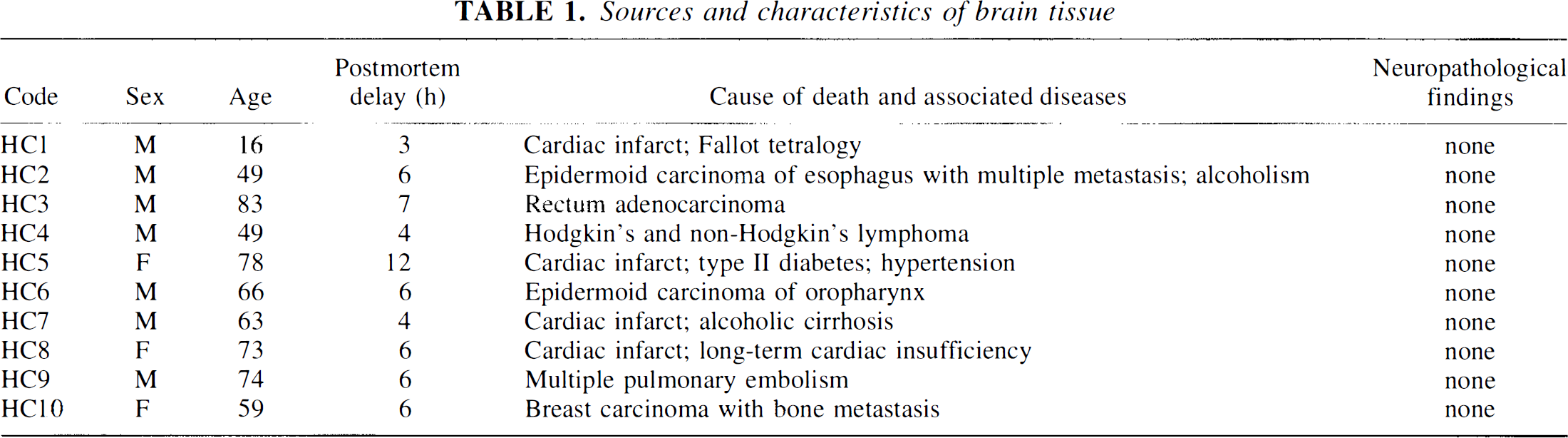
The brains of 10 control patients with no history of neurologic, metabolic, or psychiatric disorders and without histopathological lesions (Table 1) were obtained at autopsy from the Hospitals of the University of Geneva School of Medicine. Postmortem delays ranged between 3 and 12 h. The use of human tissue for this study was approved by the local ethics committee. The brains were routinely screened for histopathological lesions, using conventional histological stains and immunohistochemistry procedures, as described in detail elsewhere (Bouras et al., 1994). Some blocks of the hippocampal formation and occipital cortex were immediately frozen with CO2 for biochemical analyses, while others were fixed with 4% paraformaldehyde in 0.1 M phosphate buffered saline (PBS; pH 7.4) for 18 h, then immersed in a solution containing 5% sucrose for 48 h, and in 10% sucrose for another 48 h before being stored at −80°C. For control experiments, rabbit heart and psoas muscle freshly dissected from a rabbit killed with CO2 were frozen or fixed as described above.
Sources and characteristics of brain tissue
Production and characterization of antisera
LDH isoenzymes purified from rabbit muscle (LDH-5) and heart (LDH-1) were purchased from Sigma (Buchs Switzerland). Both products were dialyzed overnight against H2O at 4°C and subsequently stored lyophilized. Four Wistar male rats were used for each immunogen. Two rats received approximately 200 μg immunogen per injection, while the other two received approximately 50 μg per injection. At days 0 and 15, subcutaneous injections of immunogens were given in 0.2 or 0.025 ml saline emulsified with the same volume of complete Freund's adjuvant (FA). Three booster injections emulsified with incomplete FA were given intraperitoneally at day 30, 45, and 60. Ten days after the last injection, blood was removed from the tails by puncture, and serum was immediately extracted by centrifugation and stored at 4°C in the presence of 0.001% sodium azide. A first screening for the presence of antibodies was performed by immunohistochemistry in rabbit muscle and heart sections according to the procedure described below. Rats that produced reactive antisera were anesthetized with Nembutal and completely exsanguinated through the heart. Antisera were stored frozen at −80°C.
Immunohistochemistry
Sera (1:200) diluted in PBS, pH 7.4, containing 3% bovine serum albumin (BSA) and 0.1% Triton X-100 were applied overnight at 4°C on 12-μm-thick cryostat sections prepared from rabbit heart and muscle or human hippocampus and occipital cortex. Following incubation, sections were rinsed in PBS and allowed to incubate for 1 h with rabbit anti-rat IgG conjugated to peroxidase (Dako, Zug, Switzerland) at a dilution of 1:400. Immunoreaction products were visualized by exposing the sections to a solution of 0.02% 3,3′-diaminobenzidine tetrahydrochloride (DAB; Sigma) and 0.005% H2O2 in PBS for 20 min at 20°C. For comparative purposes, a set of adjacent brain sections were processed with a commercial antibody against the glial fibrillary acid protein (GFAP; rabbit IgG anti-GFAP, dilution 1:1000; Sigma).
Western blot analysis
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed according to the method of Laemmli (1970) with slight modifications. Briefly, tissue extracts from human hippocampi, rabbit heart, muscle, and brain; and mouse brain were homogenized on ice with diluted sample buffer (see below) and centrifuged at 10,000 g for 20 min. All extracts were heated at 100°C for 5 min, and approximately 5μg of protein were loaded onto the gels. The sample buffers contained 100 mM dithiothreitol in the absence of β-mercaptoethanol. For anti-LDH1, 100 mM iodoacetamide was added to the samples after heating, while for anti-LDH5, 8 M urea was added to the sample buffer and the gels contained 4 M urea. The extracts were then electrophoresed on Polyacrylamide gels (12%) and transferred electrophoretically onto a nitrocellulose membrane (Bio-Rad, Glattbrugg, Switzerland). After being blocked with nonfat dry milk (Bio-Rad) and washed in PBS, the nitrocellulose membrane was incubated for 5 h with the polyclonal antibodies anti-LDH5 and anti-LDH1 at a dilution of 1:1000. Detection of the bound antibodies was performed as described in the section Immunohistochemistry, above. Molecular weight markers were used in all gels (Kaleidoscope Prestained Standards from Bio-Rad).
Immunodot analysis
Immunodots were processed as described by Larsson (1981), with some modifications (Charnay et al., 1992). Briefly, human LDH-1 from erythrocytes and LDH-5 from placenta (Sigma), as well as the rabbit LDH-1 and LDH-5 immunogens were deposited in decreasing amounts, ranging from 10 μg to 50 pg of protein. Samples were dried on a cellulose polyacetate membrane (Sepraphore 111, Gelman Sciences Inc., Ann Arbor, MI, U.S.A.) and fixed for 90 min at 80°C with paraformaldehyde vapors. The membranes were then processed and revealed as described above (antisera dilution 1:500).
RESULTS
Characterization of the antibodies
Overall, biochemical and immunohistochemical controls clearly showed that anti-LDH1 and anti-LDH5 were specific to the LDH-1 and LDH-5 subunits, respectively. It can be seen that paraformaldehyde fixation did not alter immunoreactivity in either case (Figs. 1 and 2). Moreover, it was observed that preincubation (1 h at 37°C) of the anti-LDH1 and anti-LDH5 with an excess of their homologous antigens totally abolished the corresponding immunoreactivity visualized in human brain tissue sections (data not shown). The details of the characterization are given below.
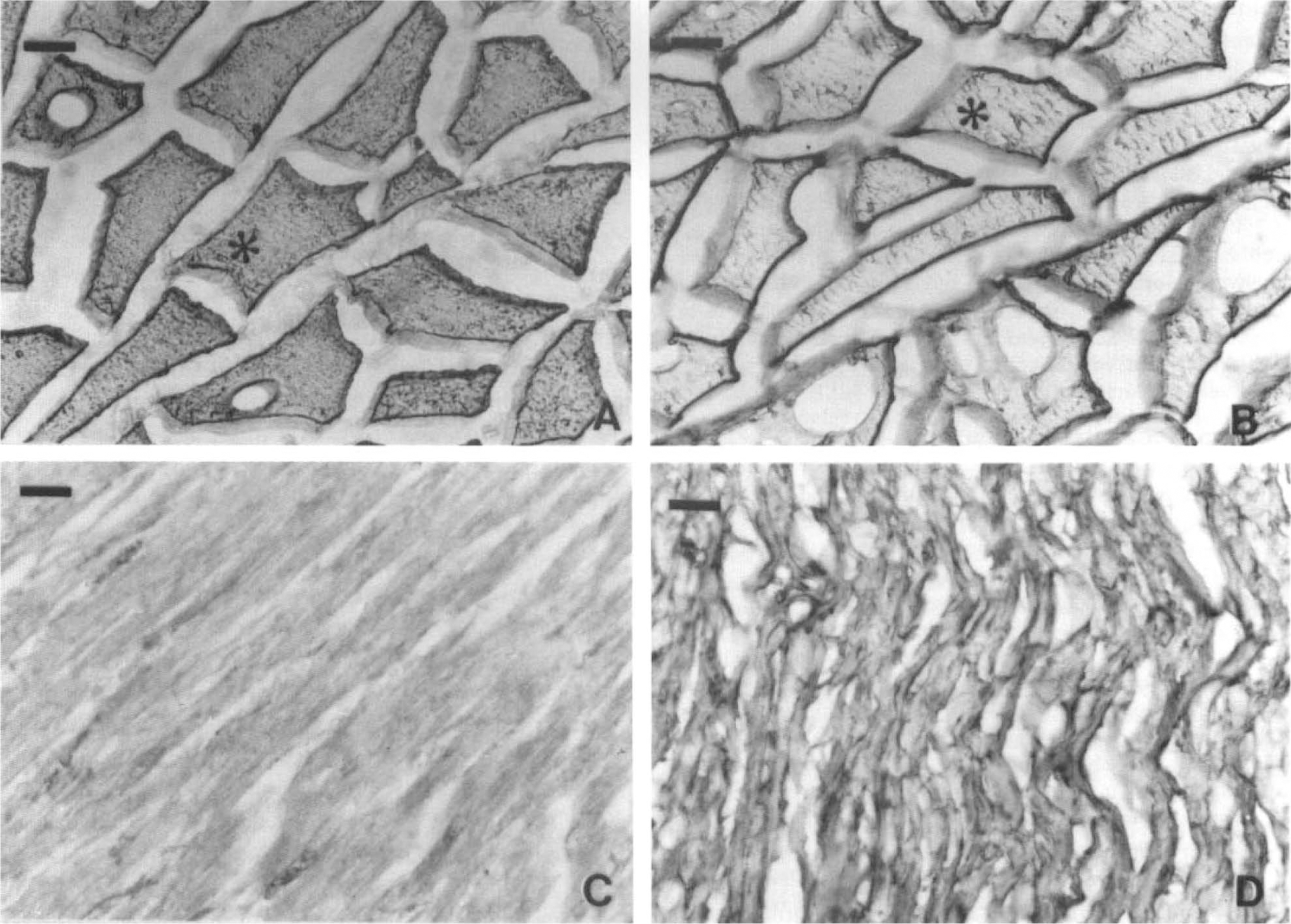
Immunohistochemical specificity of anti-LDH5 and anti-LDH1 polyclonal antibodies on rabbit muscle
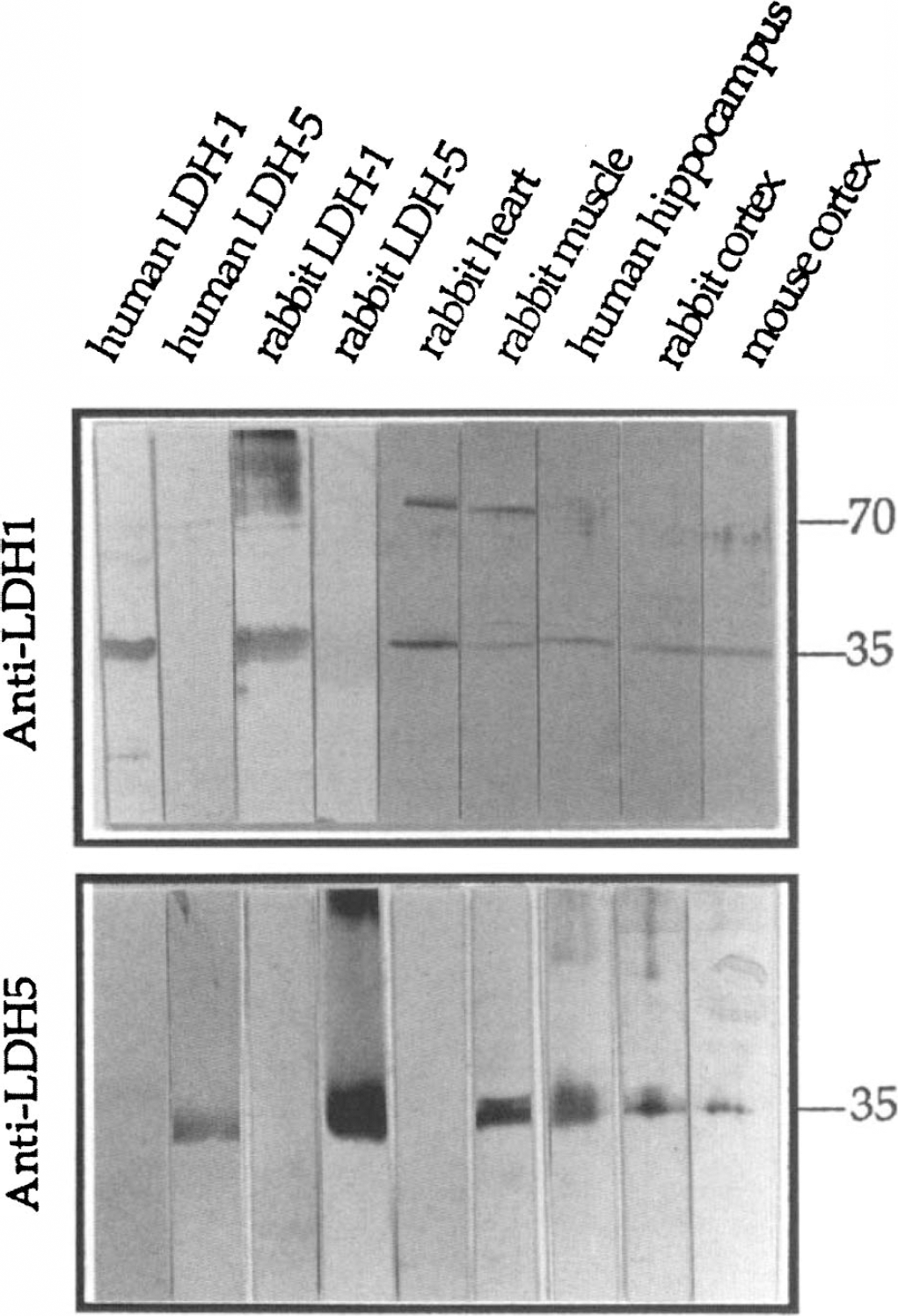
Biochemical characterization of anti-LDH1 and anti-LDH5 polyclonal antibodies by SDS-PAGE.
Immunodots. The antisera deposited on spots of LDH-1 and LDH-5 from human and rabbit sources showed specific reaction of anti-LDH5 and anti-LDH1 with LDH-5 and LDH-1, respectively. Both antisera reacted with their corresponding isoenzymes from both species, although at different dilution thresholds. Thus, in the same conditions, anti-LDH1 reacted at the lowest threshold of approximately 360 ng for human LDH-1 protein, and 38 ng for rabbit LDH-1. For anti-LDH5, thresholds were 36 ng for human LDH-5 protein, and 8 μg for rabbit LDH-5 protein (data not shown).
Immunohistochemistry on rabbit heart and muscle cryostat sections. Figure 1 shows the immunoreactivity of anti-LDH5 and anti-LDH1 in rabbit heart and muscle sections prepared and revealed in conditions identical to those used for human brain sections. Immunoreactivity of the anti-LDH5 was intense in rabbit muscle (Fig. 1A) and virtually absent in rabbit heart (Fig. 1C). In contrast, anti-LDH1 reacted faintly with rabbit muscle (Fig. 1B) and strongly with rabbit heart (Fig. 1D). Although the conformation of the fixed LDH in situ is unknown, Fig. 1 shows that tissue preparation for immunohistochemistry did not alter antigenicity.
Western blots. Figure 2 illustrates the complete characterization of anti-LDH5 and anti-LDH1 against a variety of denatured antigens from different tissues and species. In all cases, both antibodies were specific for the monomeric form of their corresponding subunit, whose molecular weight is 35 kDa for both the LDH-5 and LDH-1 monomers. The anti-LDHl reacted only with a 35-kDa band on all brain extracts from three different species, i.e., human, rabbit, and mouse as well as with pure LDH-1 from human erythrocytes and rabbit heart. Since the anti-LDH1 did not recognize pure LDH-5 subunits purified from human and rabbit tissues, it can be assumed that the 70-kDa bands observed with rabbit muscle and heart extracts are probably a homodimeric LDH-1 subunit form that renatured in the course of electrophoresis; this phenomenon is not observed on extracts from human hippocampi or rabbit and mouse brains. The anti-LDH5 reacted specifically with a 35-kDa band from pure human and rabbit LDH-5, rabbit muscle extracts, as well as from human, rabbit, and mouse cerebral cortical extracts. No cross-reactivity was observed with human and rabbit LDH-1 and with rabbit heart extracts.
Immunohistochemistry
The characterized antisera against LDH-1 and LDH-5 subunits revealed patterns of immunostaining that were constant in all 10 human brains investigated (Figs. 3–6).
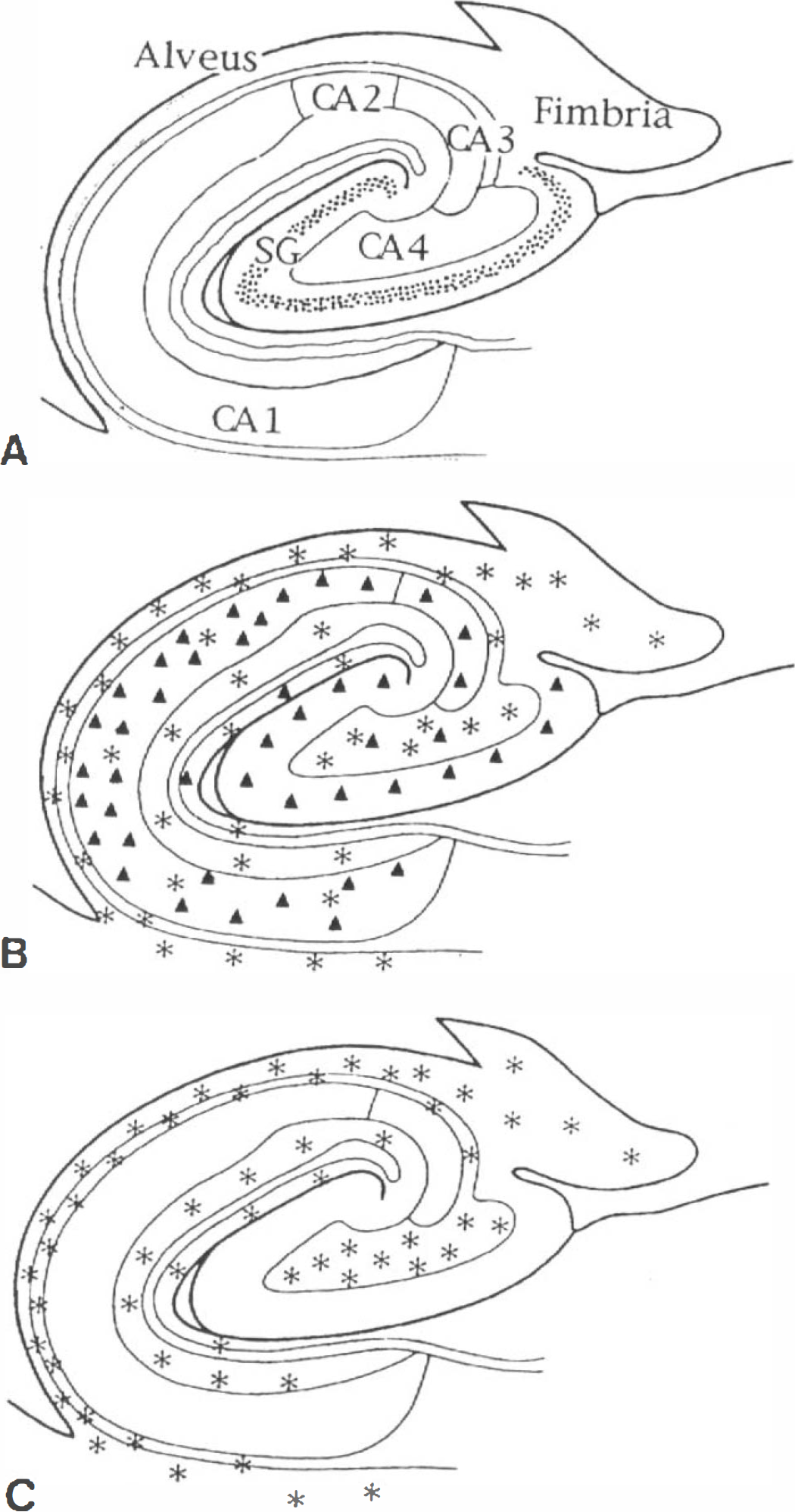
Mapping of LDH-1
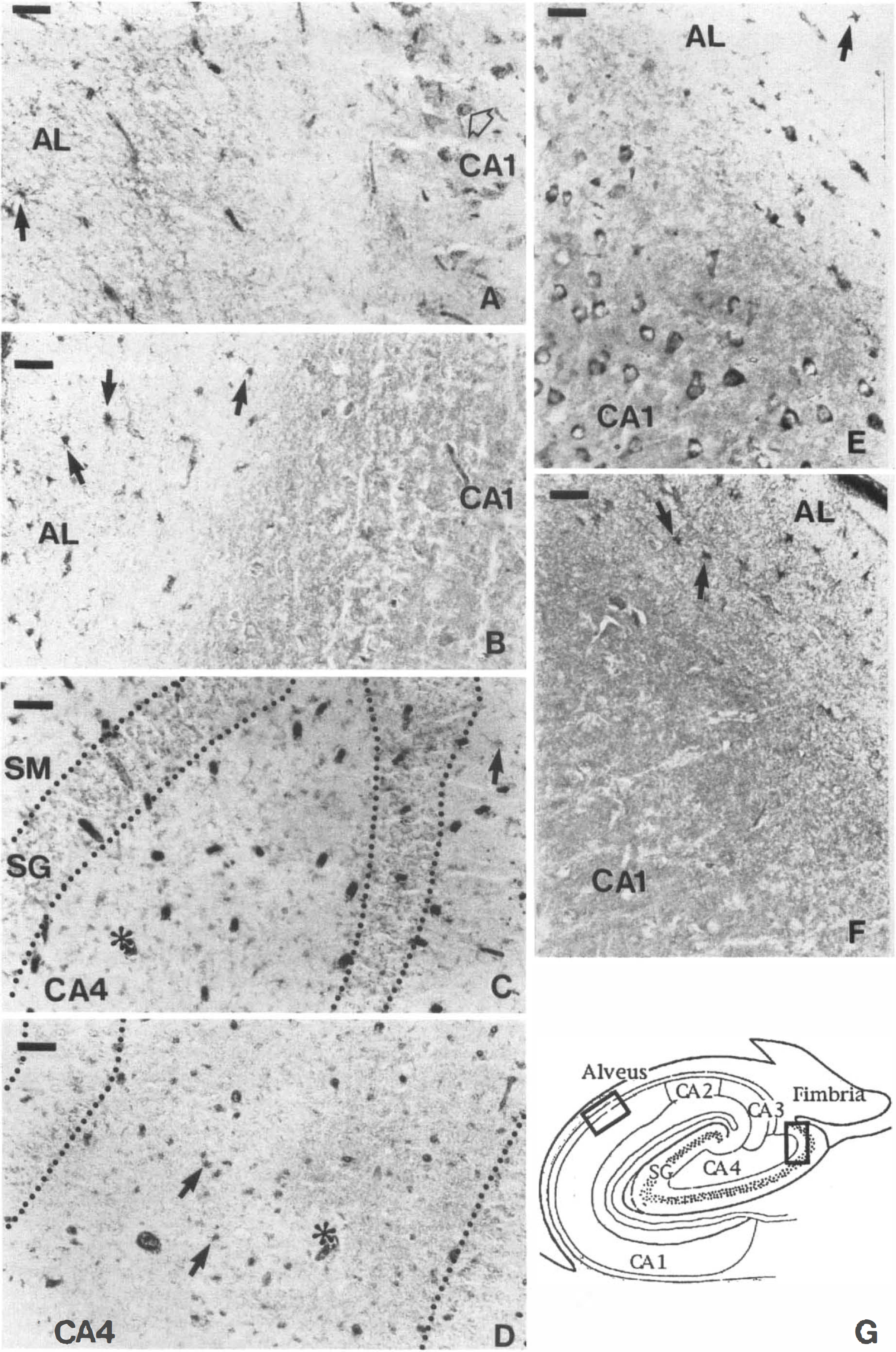
Comparative photomicrographs showing
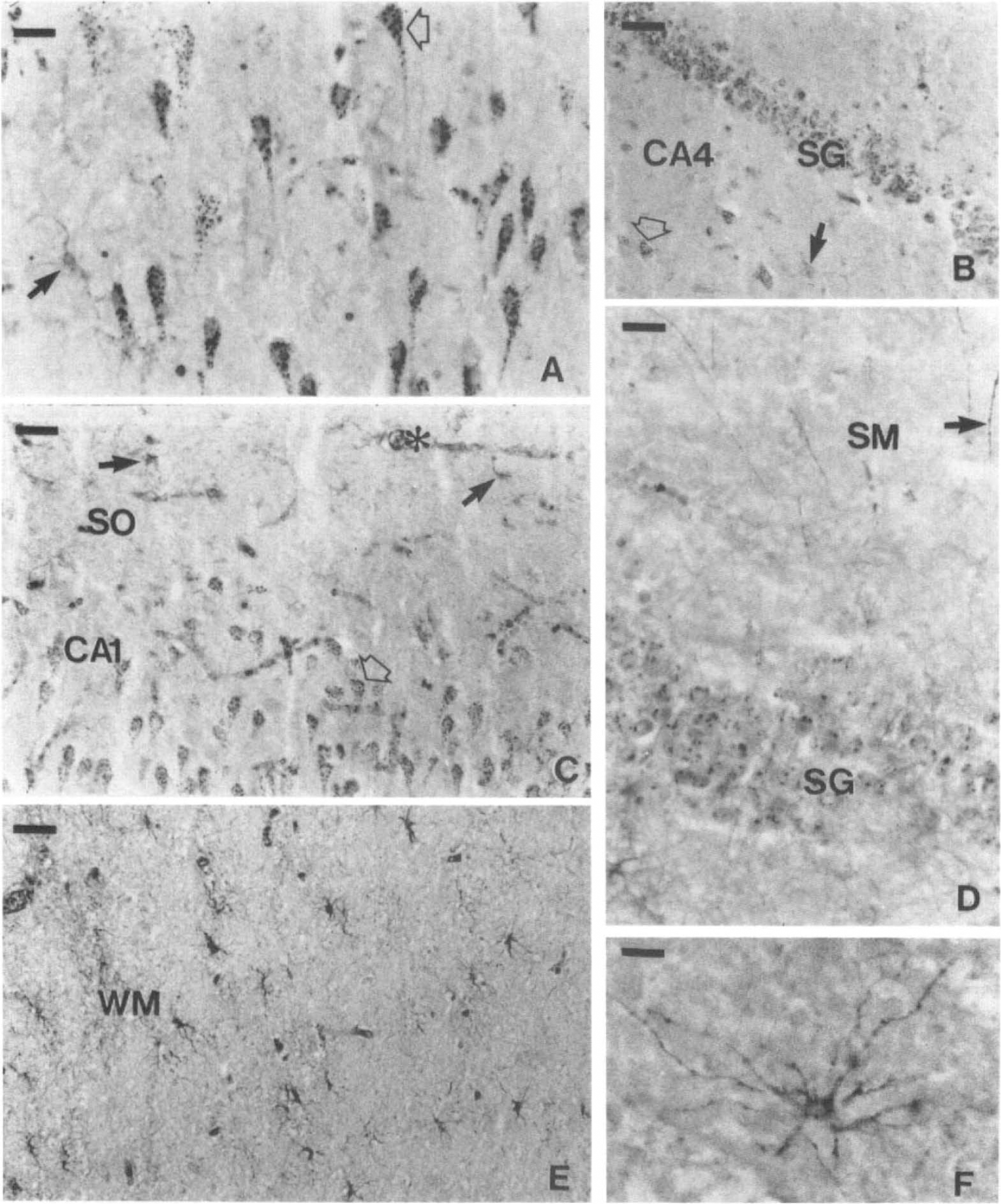
Photomicrograph showing LDH-1 immunoreactive cells in the human hippocampus.
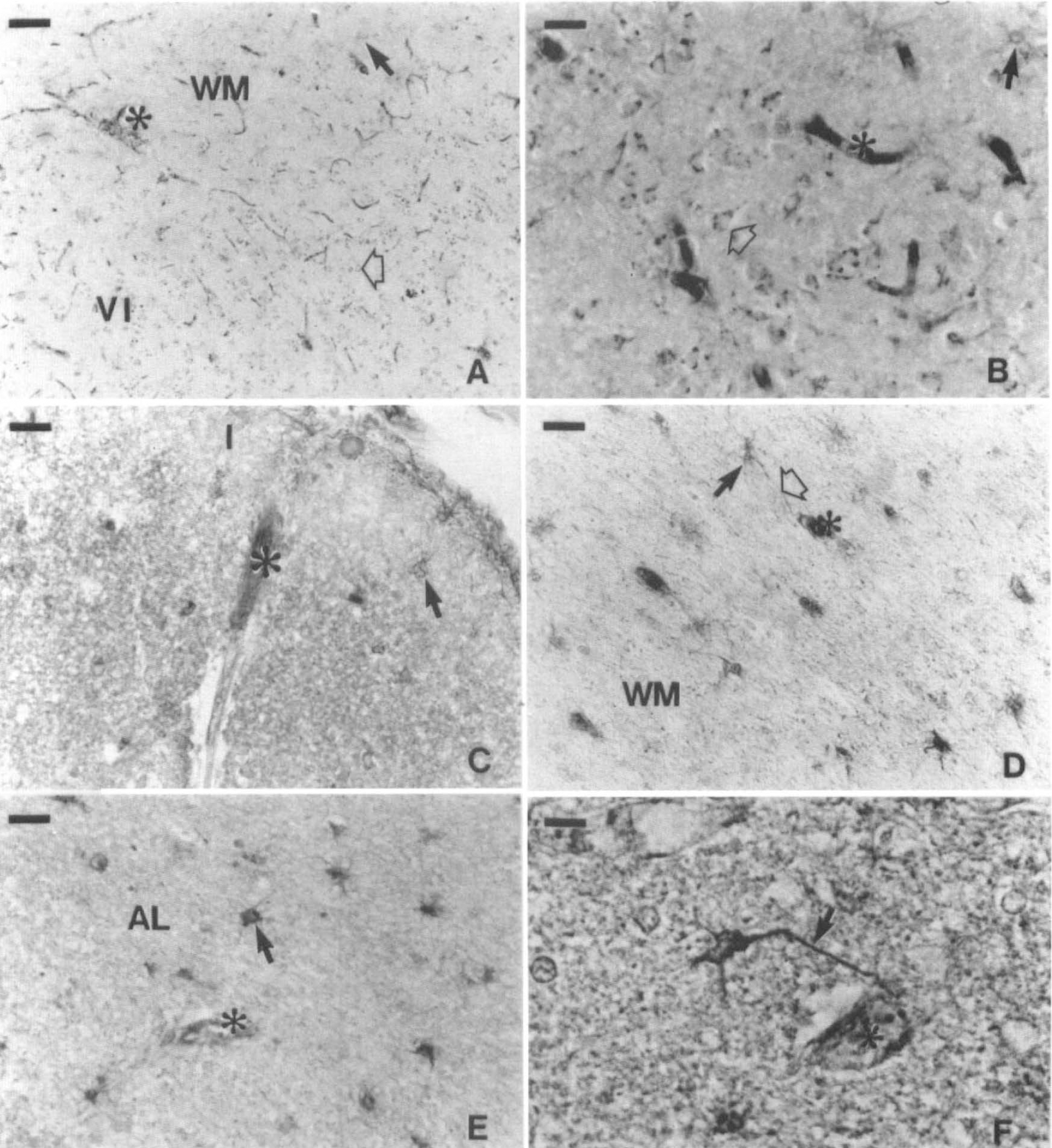
Photomicrograph showing anti-LDH1
Human hippocampus. As summarized in Table 2, the anti-LDH5 immunoreactivity was only observed within astrocytes. Its distribution was usually uniform and homogenous (Fig. 3C), but clusters of several astrocytes displaying a particularly intense staining were occasionally observed. The anti-LDH5 immunoreactivity was most intense in astrocytes localized throughout the occipital (Fig. 6D) and the hippocampal white matter (not shown, but similar to Fig. 6D), the alveus, the stratum oriens (Fig. 4B and F), and the CA4 (Fig. 4D). They were less abundant than astrocytes stained with a polyclonal antibody directed against the GFAP on adjacent sections. Unlike the typical astrocytes stained with the anti-LDH5 (Fig. 6E and F), some had a granular staining comparable to the granules observed in anti-LDH1 immunoreactive neurons (Fig. 5A). This granular astrocytic staining was principally observed in CA4 (Fig. 4D) and stratum moleculare. In the stratum moleculare, as well as in the stratum radiatum, a lower density of astrocytes stained with the anti-LDH5 was observed; their processes were not as clearly visualized as those of other regions. Astrocytes stained with the anti-LDH5 were occasionally observed in the CA1 (Table 2), although the intensity and definition of their staining was considerably weaker in this region. Rarely, varicose fibers stained with the anti-LDH5 were observed in the alveus. Their density was low compared to the fibers stained by the anti-LDH1 (not shown, but similar to Fig. 5D). Most remarkably, neurons were never stained by the anti-LDH5 serum (Fig. 4B, D, and F and Table 2).
Semiquantitative analysis in hippocampus a
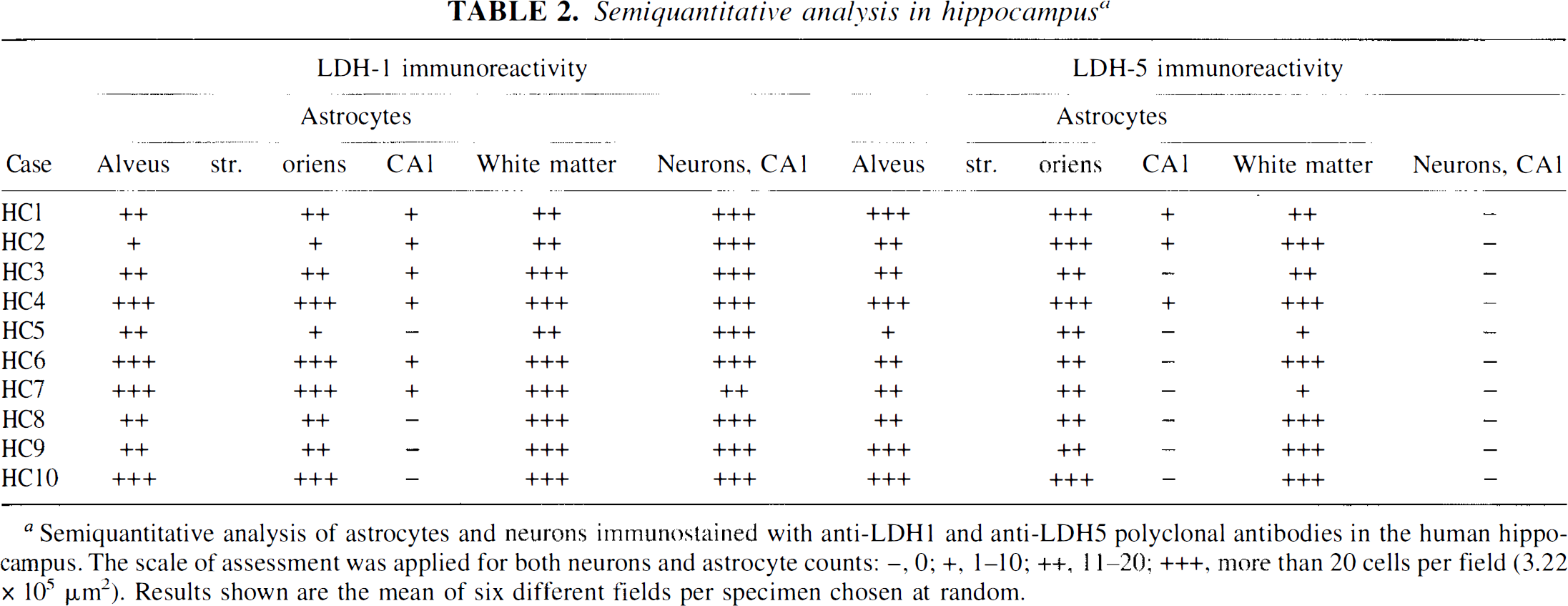
Semiquantitative analysis of astrocytes and neurons immunostained with anti-LDH1 and anti-LDH5 polyclonal antibodies in the human hippocampus. The scale of assessment was applied for both neurons and astrocyte counts: -, 0; +, 1–10; ++, 11–20; +++, more than 20 cells per field (3.22 × 105 μm2). Results shown are the mean of six different fields per specimen chosen at random.
The anti-LDH1 immunoreactivity in the hippocampus is illustrated in Fig. 5 and its distribution summarized in Fig. 3B. Cell bodies were stained (Figs. 4A, C, and E and 5A-D) and varicose fibers could be observed in the alveus (not shown), in the stratum moleculare (black arrow of Fig. 5D) and stratum radiatum (not shown). Astrocytes were also stained in the gray (solid arrows of Fig. 5A and B) and the white matter (Fig. 5E and F). They were abundant in the alveus and stratum oriens (Fig. 4A and E, 5C). The number of astrocytes stained with the anti-LDH1 was generally slightly greater than those stained by the anti-LDH5 and comparable in number with astrocytes stained with a polyclonal antibody directed against GFAP.
Occipital cortex. Despite a smaller number of samples available (cases HC4, HC9, HC 10), a similar trend could be observed in visual areas 17 and 18. The anti-LDH5 immunoreactivity was still principally observed in astrocytes of the first cortical layer (Fig. 6C) and white matter (Fig. 6D). Neurons were essentially unlabeled by the anti-LDH5, although a weak and diffuse staining was very rarely seen in neurons of layers IVc and VI of area 17. Interestingly, the anti-LDH5 background staining was also more intense in these two visual areas.
The anti-LDH1 immunoreactivity revealed a pattern similar to that observed in the hippocampus: astrocytes were stained in the white (solid arrow of Fig. 6A) and gray matter (solid arrow of Fig. 6B), granules could be observed in virtually all neurons (open arrows of Fig. 6A and B), and a few varicose fibers were noted in gray matter (not shown). In area 17, the density of neuronal granules was higher in layers IVc and VI (not shown). Similar immunostaining patterns were observed in area 18, with only a decrease in the background staining of the anti-LDH5 and a uniform anti-LDH1 staining density of the neurons throughout all cortical layers.
DISCUSSION
Immunohistochemistry
In this article, we have described the immunohistochemical distribution of LDH-1 and LDH-5 subunits in two regions of the human brain. Previous histochemical studies in the monkey occipital cortex demonstrated that LDH activity was mainly localized in layers IV and presumptively in layer VI as well as in astrocytes of the first cortical layer (Friede and Fleming, 1963). The immunohistochemical distribution patterns observed in the human occipital cortex also indicate a predominant localization of the anti-LDH1 and anti-LDH5 immunoreactivity in these layers, as well as in astrocytes of the first cortical layer. In addition, the anti-LDH5 immunoreactivity was detected in astrocytes of the occipital white matter, where no LDH activity had been detected (Friede and Fleming, 1963). Interestingly, layers IV and VI, which appear enriched in LDH immunoreactivity, exhibit a higher uptake of deoxyglucose in the visual cortex of the monkey (Kennedy et al., 1976).
In hippocampus, a good correlation exists between the distribution patterns of immunoreactivity reported here for the human brain and previous histochemical stainings in the rat and monkey brains. Thus, in monkey, LDH activity was high in the perikarya of pyramidal cells and in the fascia dentata, alveus, stratum oriens, and moleculare, while it was low in the stratum radiatum (Friede and Fleming, 1963). Another study reported low LDH activity in the stratum lacunosum-moleculare and the outer dentate molecular layer of the rat (Borowsky and Collins, 1989). These authors also found a maximal LDH activity in the stratum oriens, where LDH-5 and LDH-1 subunit immunoreactivity was localized mainly in astrocytes (Figs. 4A, B, E, F, and 5B). Despite a generally good correlation between our observations and previous histochemical studies, some discrepancies were observed. Thus, high LDH activity was found in regions weakly stained by anti-LDH5 and anti-LDH1, such as the stratum radiatum and the inner third of the dentate molecular layer (Borowsky and Collins, 1989). The neuronal cell bodies of both CA and fascia dentata were not stained histochemically (Borowsky and Collins, 1989), while showing immunoreactivity for anti-LDH1 (Fig. 5A–D). This latter finding correlates well with observations in monkeys, where a strong activity in the inner third of the stratum moleculare was found and neuronal cell bodies of CA and fascia dentata were stained (Friede and Fleming, 1963).
Results described in this article are also in agreement with previous electrophoretic analyses indicating the predominance of LDH-1 subunits .in the adult brain of various mammalian species including human (Miura, 1966; Gerhardt-Hansen, 1968). In the present study, LDH-1 subunits are immunolocalized both in astrocytes and neurons. However, results reported in this article indicate that the LDH-5 subunits appear to be exclusively localized in astrocytes both in hippocampus and visual areas 17 and 18. Our observations are in agreement with a previous study reporting an enrichment of LDH-5 subunits in selected astrocyte populations of the rat brain (Venkov et al., 1976) and also in astroblasts cultured from chick embryo brains (Tholey et al., 1981).
Biochemical considerations
Pyruvate represents the end point of glycolysis in mammalian cells. One of the determining factors in its subsequent fate is the NADH/NAD+ ratio. When this ratio is low, pyruvate is converted preferentially to acetyl coenzyme A by pyruvate dehydrogenase (which is normally inhibited by high levels of NADH). Subsequently, it enters the tricarboxylic acid cycle to be oxidized and produce energy through the coupled oxidative phosphorylation. Under conditions of high NADH/NAD+ ratio however, which may occur when glycolysis exceeds the capacity of the tricarboxylic acid cycle, pyruvate is converted to lactate by LDH (Stryer, 1988). In order to sustain a high glycolytic rate, this reaction, which uses NADH as a cofactor, is essential for the regeneration of NAD+ used by the glycolytic enzyme glyceraldehyde phosphate dehydrogenase (Stryer, 1988).
LDH is therefore at the junction between glycolysis and the tricarboxylic acid cycle. Tissues that display intense but brief increases in activity generally can sustain high glycolytic rates and produce high amounts of lactate. This is the case in the fast-twitch fibers in skeletal muscles, which produce and release lactate in the circulation during activity. Such tissues tend to contain higher proportions of the LDH-5 subunit, while others such as the heart, which can utilize lactate as a substrate, are enriched in LDH-1 subunit (Plagemann et al., 1960; Wroblewski et al., 1960; Cahn et al., 1962).
Although the kinetic characteristics of enzymes in vitro do not necessarily correspond to their true activity in vivo, they do set a limit to their activity and are a useful indication of their potential significance. Despite numerous influences (including pH and temperature) and the consequent limitations in comparing the kinetic differences between LDH subunits, two major points can be underlined that remain true throughout several conditions and species (Gerhardt-Hansen, 1968; Holbrook et al., 1975): first, LDH-1 has a higher substrate affinity (lower Michaelis constant) than LDH-5; LDH-1 maximal velocity (Vmax) is inferior to that of LDH-5 (Nitisewojo and Hultin, 1976). Second, under physiological conditions, LDH-1 is allosterically inhibited by pyruvate to a considerably greater degree than LDH-5 (Dawson et al., 1964; Gerhardt-Hansen, 1968; Bishop et al., 1972). Lactate also inhibits LDH-1, but at much higher concentrations than pyruvate (Holbrook et al., 1975).
Homozygous deficiencies of both LDH-1 and LDH-5 subunits, which emphasize the glycolytic nature of the LDH-5 subunit, have been reported in humans. Thus the absence of the LDH-5 subunit has been linked to exertional myopathies with myoglobinuria (Kanno et al., 1983). During exercise under ischemic conditions, the LDH-5 subunit-deficient patients clearly show an inability to release lactate in the venous blood, as compared to control patients (Kanno et al., 1988). Instead, a pyruvate peak is observed in these patients, indicating that the LDH-5 subunit is necessary in vivo for the production and release of high amounts of lactate and that it cannot be efficiently replaced by the LDH-1 subunit for that purpose.
It has been proposed that the LDH-5 isoenzyme is most often associated with glycolytic tissues because it is better adapted to their requirements, being optimally designed to process large amounts of substrate (Dawson et al., 1964). In contrast, kinetic constraints of the LDH-1 isoenzyme are more compatible with the requirements of oxidative metabolism (Dawson et al., 1964), i.e., a high affinity for substrates in a low pyruvate environment, since pyruvate is avidly oxidized via the tricarboxylic acid cycle to face energy demands. In such tissues, including the heart, which maintain low pyruvate concentrations, lactate is not only a substrate for LDH, but it becomes an adequate metabolic fuel. Indeed, after conversion into pyruvate facilitated by the characteristics of the LDH-1 subunit, lactate becomes an extremely valuable energy fuel, potentially yielding 18 ATP molecules. It can therefore be reasonably argued that cells expressing high levels of LDH-5 subunits are likely to display high glycolytic rates and have the capacity to produce large amounts of lactate. On the other hand, cells expressing high levels of LDH-1 subunits in the relative absence of LDH-5 subunits will display a metabolic oxidative profile with a better capacity to utilize lactate as an energy fuel.
In summary, the enrichment of LDH-5 subunits in astrocytes and the presence of LDH-1 subunits in neurons support the hypothesis of a predominant lactate production in astrocytes and oxidation by neurons after conversion of lactate to pyruvate. The predominant immunolocalization of pyruvate dehydrogenase in neuronal perikarya in the rat (Bagley et al., 1989) and also in the human hippocampus (manuscript in preparation) are consistent with this view.
Physiological implications
The foregoing immunohistochemical observations and biochemical considerations are consistent with the fact that lactate is the major metabolic substrate released by astrocytes, particularly during situations of high energy demand such as activation (Pellerin and Magistretti, 1994).
There is increasing evidence from functional brain imaging studies that the brain transiently resorts to glycolysis to face the metabolic needs linked to activation. Thus, the combined analysis of blood flow, oxygen consumption, and glucose utilization by PET has indicated an uncoupling during activation, whereby blood flow and glucose utilization increase in excess of oxygen consumption (Fox and Raichle, 1986; Fox et al., 1988). This observation documents the occurrence of activation-induced glycolytic processing of glucose. Further confirming the existence of activation-induced glycolysis are 1H-magnetic resonance spectroscopy (MRS) studies in which a lactate peak during physiological activation is observed (Prichard et al., 1991; Sappey-Marinier et al., 1992). Transient lactate formation during activation has also been demonstrated in rat brain with microdialysis studies (Fellows et al., 1993). Given the marked (if not exclusive) enrichment of LDH-5 subunits in astrocytes, this set of converging evidence suggests that the increased lactate production during activation is due to the glycolytic processing of glucose in astrocytes. Indeed, the observation that glutamate, the main excitatory neurotransmitter released from activated excitatory terminals, stimulates glucose uptake (Pellerin and Magistretti, 1994; Takahashi et al., 1995) and lactate release by astrocytes (Pellerin and Magistretti, 1994) is consistent with this view. Although the possibility of some lactate losses in the circulation cannot be entirely ruled out under activated conditions, the presence of LDH-1 subunits in neurons and their processes suggests that conversion of lactate to pyruvate for subsequent oxidation is likely to occur. Indeed, a lactate transfer from glia to neurons has been demonstrated in the guinea pig retina (Poitry-Yamate et al., 1995). This sequence of events would predict a “recoupling,” whereby oxygen consumption should increase, with a short temporal delay after activation, and the lactate peak should disappear.
Thus biochemical, immunohistochemical, and functional brain imaging studies support the existence of an “astrocyte-neuron lactate shuttle” whereby during activation, glucose, taken up by astrocytic end-feet surrounding intraparenchymal capillaries, is processed glycolytically to lactate which is released by astrocytes, providing an energy substrate for neurons.
Footnotes
Abbreviations used
Acknowledgment:
This work was supported in part by FNRS grants 31-40565.94 to P.J.M. The authors wish to thank Drs. A. Paoloni-Giacobino and F. Fasel for making available human samples, P.-Y. Vallon for developing the microphotographs, Maria Surini for technical help, and Dr. P. Giannakopoulos for critically reading the manuscript.