
Abstract
Although antibodies are commercially available to allow investigation into the biology of the age-regulating protein Klotho, problems with antibody specificity and application functionality are significant barriers to progress. Chief among these limitations is the inability of current tools to allow in vivo validation of binding partners originally identified through transfection of tagged proteins. To overcome this barrier, we generated a series of hybridoma cell lines by immunizing rats with a GST-KL1 fusion protein. Purified antibodies generated from these cell lines differentially detect human or mouse Klotho protein via Western blot, immunocyto/histochemistry, and immunoprecipitation. Specificity of antibody binding to Klotho was confirmed by mass spectrometry following immunoprecipitation. With this confidence in antibody specificity, co-immunoprecipitation was utilized to validate the interaction of Klotho/FGFR and Klotho/wnt7a in mouse kidney lysates.
Introduction
P
Generating high quality, specific antibodies to KL is not trivial. KL is highly homologous across mammalian species and is a plasma membrane resident protein subject to shedding, implicating a likely increased tolerance to antibody induction. Although commercial antibodies are available, none are optimal, as defined by an ability to detect protein with high specificity across species and function in multiple experimental paradigms. For example, the most widely utilized and specific antibody available, KM2076, detects KL from human and rodent samples by Western blot when highly expressed, namely from transfected cells or in the kidney.(16–18) However, detection of KL at lower levels of expression is complex, even when mRNA is readily detectable.(19) As well, although KM2076 is robust via Western blot, its ability to work in immunohistochemistry (IHC) varies across laboratories. Antibody AF1819 detects only murine KL and appears to be the best option for detection of KL by IHC, revealing highly specific staining of KL in wild-type but not knockout tissues.(19–21) Immunoprecipitation (IP), critical for identification and validation of KL binding partners, is limited to use of tagged proteins in transfection paradigms as available antibodies are insufficient to allow IP of endogenous, untagged protein. This has resulted in a striking lack of in vivo confirmation of binding partner interactions with KL. While IP of FGFR (fibroblast growth factor receptor) from kidney allowed detection of KL,(9) the reciprocal IP was not possible and KL/FGFR remains the lone binding partner interaction to be confirmed under endogenous conditions. Without the ability to confirm and expand upon in vitro findings using physiologically relevant expression levels, our ability to further dissect the meaning of protein-protein interactions for KL biology remains limited. Herein we report the generation of rat monoclonal antibodies capable of both detecting multiple species of KL and functioning in diverse experimental applications.
Materials and Methods
Plasmid construction
The human KL plasmid was generated by cloning KL(7) into the pcDNA3.1 myc-His vector using the BamHI/XhoI restriction sites. The stop codon was mutated from mouse KL (IMAGE clone, Thermo Scientific, Waltham, MA) and restriction sites added to allow transfer from the vendor vector to pcDNA3.1A myc-his vector (Life Technologies, Grand Island, NY) using HindIII/XhoI (NEB) restriction sites.
Animals
Sprague Dawley rats were obtained commercially for immunization. The KL knockout line was obtained from M. Kuroo.(1) Eight-week-old male KL knockout mice and wild-type litermate controls (n=3; 129/SvImJ strain) were used. Animals were euthanized by terminal perfusion with tyrode solution (137 mM NaCl, 2.7 mM KCl, 1 mM MgCl2, 1.8 mM CaCl2, 0.2 mM Na2HPO4, 12 mM NaHCO3, and 5.5 mM glucose). Following perfusion, kidneys were flash frozen in isopentanes and stored at −80°C until use. Alternatively, kidney and brain were post-fixed in Bouin's fixative prior to paraffin embedding. All procedures were approved by the UAB IACUC committee.
Antigen preparation
The region corresponding to amino acids 55-261, and thus the N-terminal half of the KL1 domain of human KL, was PCR amplified from a pcDNA3.1 vector containing the full cDNA sequence using primers: F-5′-TCCGGAGGATCCGGCCTCTTCCAGGGCACCTTCC; R-5′-ACCCGGACTCGAGCTACGGGCTGCCCCGGATGCCGGG. The resulting KL fragment and pGEX-6P2 (GE Healthcare, Pittsburg, PA) were digested using BamHI and XhoI (New England Biolabs, Ipswich, MA) and KL ligated in-frame with GST. Recombinant protein was generated by IPTG (isopropyl-β-D-1 thiogalactopyranoside; 0.4 mM) induction at room temperature for 16 h. Bacteria were pelleted and lysed in STE buffer (100 mM NaCl, 10 mM Tris [pH 7.6], 1 mM EDTA) with protease inhibitors (RPI). Lysates were treated with lysozyme (0.1 mg/mL) for 15 min. Dithiothreitol (DTT) and sarcosyl were then added to a final concentration of 5 mM and 1.5%, respectively. Following sonication, Triton-X100 (1% final concentration) was added to the cleared supernatants followed by incubation with glutathione-agarose beads (Thermo Scientific) overnight. The GST-KL bound beads were washed in NETN buffer (0.5% NP-40, 1 mM EDTA, 20 mM Tris [pH7.6], and 100 mM NaCl). GST-KL was eluted from the beads in 10 mM glutathione. The concentration of fusion protein was estimated by comparison to BSA standards on Coomassie stained gel.
Hybridoma generation and initial screening
Rats were immunized with a fusion protein consisting of the amino acids 55-261 of KL fused to GST (GST-KL). Rats received a primary immunization of 200 μg of GST-KL emulsified in complete Freund's adjuvant administered subcutaneously in the thigh of each rear leg. Fourteen days after the primary immunization, rats were boosted with 200 μg of KL in incomplete Freund's adjuvant administered subcutaneously near the initial immunization sites. The final boost of 100 μg was administered in saline on day 20 and injected subcutaneously at the base of the tail. On day 21, rats were euthanized and lymphocytes from the popliteal and inguinal lymph nodes were fused to P3U-1 myeloma cell line. Hybridoma supernatants were screened by ELISA for reactivity with GST-KL1 fusion protein or GST alone. Hybridomas that reacted exclusively with GST-KL were subcloned by limiting dilution, isotyped, and expanded for supernatant collection and protein G purification. The derived antibodies KL-115 and KL-234 are available through the University of Iowa Developmental Studies Hybridoma Bank.
Cell culture and Western blot analysis
HEK293T cells (ATCC) were cultured in DMEM containing 10% FBS and 1% penicillin/streptomycin (Mediatech, Herndon, VA). Cells were maintained under standard growth conditions. To assess KL protein expression, HEK293T cells were transfected with plasmids expressing human KL cDNA, mouse KL cDNA and empty vector. After 24 h, cells were homogenized in RIPA buffer (150 mM NaCl, 50 mM Tris [pH 7.5], 1% Triton-X100, 0.5% sodium deoxycholate, and 1% sodium dodecyl sulfate) containing protease inhibitors (Thermo Scientific). Protein from mouse kidney was isolated by homogenization and sonication in RIPA buffer containing protease inhibitors. Protein concentration was determined by BCA assay, per the manufacturer's instructions (Thermo Scientfic). Thirty μg of total protein from transfected cells and 50 μg of total protein from kidney were loaded onto 10% polyacrylamide gels. Following transfer to nitrocellulose membranes (Fisher, Waltham, MA), non-specific binding was blocked by incubation in 5% non-fat dry milk in TBST. Primary antibodies were incubated overnight at 4°C with gentle agitation in 0.3% BSA/TBST. Primary antibodies utilized were KL-115 (1:1000; 0.9 mg/mL), KL-234 (1:400; 1.2 mg/mL), tubulin (1:5000, DSHB), FGFR-1 (1:250 SC7945, Santa Cruz Biotechnology, Dallas, TX), wnt7A (1:500 SC26361, Santa Cruz Biotechnology), AF1819 (1:100, R&D Systems, Minneapolis, MN), KM2076 (1:500, Sceti KK, Japan), and myc (SC789, Santa Cruz Biotechnology). Following binding of secondary antibodies conjugated to HRP protein was detected using chemiluminescent reagents (Immobilion, Millipore; SuperSignal, Thermo Scientific) by exposure to film (Phenix Research, Candler, NC). Tubulin was detected following membrane stripping in One Minute Stripping Buffer (GE Healthcare).
Immunocytochemistry
HEK293T cells were transfected with plasmids expressing human KL cDNA, mouse KL cDNA, and empty vector pcDNA per the manufacturer's protocol using Attractene (Qiagen, Valencia, CA). After 24 h, cells were fixed in 4% paraformaldehyde and antigen retrieval was performed in hot 10 mM citrate buffer. Endogenous peroxidases were inactivated in 0.3% H2O2, followed by permeabilization in 0.5% Triton-X100 in phosphate buffered saline (PBST). PBS (phosphate buffered saline) washes were conducted between each subsequent step. Cells were blocked in blocking buffer (5% horse serum and 0.03% PBST) and then exposed to KL-115 (1:100, 0.9 mg/mL) and KL-234(1:100, 1.2 mg/mL). Secondary antibodies labeled with Alexa594 (Life Technologies) were used to detect primary antibodies. Samples were mounted in Prolong Gold anti-fade mounting media (Life Technologies). Results were confirmed in three independent transfections.
Immunohistochemistry
Ten μm microtome sections of paraffin embedded brain and kidney were mounted on Superfrost slides (Fisher). Slides were dried overnight in a 42°C oven. Paraffin was removed and the tissue rehydrated by incubation in washes of Citrisolv (Fisher) and isopropanol followed by rehydration in running distilled water. Antigen retrieval was performed in 10 mM citrate buffer in a rice cooker. Following slow equilibration to room temperature, sections were incubated in H2O2 and PBST as described above before block and incubation in primary antibody. In addition to antibodies as above, they were also exposed to anti-KL antibody (1:20, AF1819; R&D Systems) diluted in 0.5% PBST overnight at 4°C. Species-specific polymer was applied (ImmPRESS, Vector labs, Burlingame, CA) prior to detection with TSA-Plus reagent (1:400; Perkin Elmer, Melville, NY). Sections were washed in PBS containing DAPI and mounted in Prolong Gold anti-fade mounting media (Life Technologies). Samples were imaged on an Olympus BX53 fluorescent microscope. Results were confirmed in three to four independent adult mouse organs. Antibody specificity was tested by incubation of sections with no primary, whole serum diluted to the same concentration as the primary antibody, and utilizing knockout tissue.
Immunoprecipitation and mass spectrometry
Lysates were prepared from HEK293T cells transfected with human KL or control vector, as described above. Precleared lysate was incubated with 10 μL of one of the primary antibodies KL-115 (0.9 mg/mL) or KL-234 (1:100, 1.2 mg/mL) overnight. Antibody complexes were pulled out of solution by the addition of protein G agarose (Santa Cruz Biotechnology). SDS-sample buffer was added to the IP beads, boiled, and run into a 10% SDS Bis-Tris PAGE (Life Technologies), stained overnight with colloidal blue (Life Technologies), and destained with water. The entire band containing protein was excised and equilibrated in 100 mM ammonium bicarbonate, reduced, carbidomethylated, dehydrated, and digested with Trypsin Gold (Promega, Fitchburg, WI). Following digestion, peptides were concentrated under vacuum and resolubilized in 0.1% formic acid prior to 1D reverse phase nLC-ESI-MS analysis.
Digested peptides were injected onto a Surveyor HPLC plus (Thermo Scientific, San Jose, CA) using a split flow configuration on the back end of a 100 micron I.D. ×13 cm pulled tip C-18 column (Jupiter C-18 300 Å, 5 micron, Phenomenex, Torrance, CA). This is followed by a Thermo Orbitrap Velos Pro hybrid mass spectrometer equipped with a nano-electrospray source (Thermo Scientific). All data were collected in CID mode. Peptide fractions were directly sprayed into the mass spectrometer over a 1 h gradient set to increase from 0–30% acetonitrile in D.I. H2O containing 0.1% formic acid with a flow rate of 0.3 μL/min. Following each parent ion scan, fragmentation data were collected on the top most intense 18 ions. During data collection, the instrument is configured as follows: spray voltage 1.9kV, capillary temperature 170°C, 1 microscan for all scans, with a maximum inject time set for full MS scan in FT mode at 500 ms, and for fragmentation (MSn) in the Ion Trap at 50 ms. The parent ion scans were obtained at 60 K resolution in FT mode with a minimum signal threshold of 2000 counts for MSn activation. The activation settings were charge state 3, isolation width 2.0 m/z, normalized collision energy 30.0, activation Q 0.250, and activation time 25 ms. For the dependent scans, both charge state screening and dynamic exclusion were enabled with the following settings: repeat count 2, repeat duration 30 s, exclusion list size 500, and exclusion duration 60 s.
The XCalibur RAW files collected in profile mode were centroided and converted to MzXmL using ReADW, v. 3.5.1. The .mgf files were created using MzXmL2Search for all scans with a precursor mass between 300 Da and 1200 Da. The data were searched using SEQUEST set for three maximum missed cleavages, a precursor mass window of 20 ppm, trypsin digestion, variable modification C at 57.0293, and M at 15.9949. For the fragment-ion mass tolerance, 0.0 Da was used. Searches were performed with a Human subset of the UniProt100 database. The resulting list of peptide IDs generated was filtered using Scaffold (Proteomesoftware, Portland, OR). The filter cut-off values were set with peptide length (>5 AAs) and excluding peptides with a MH+1 charge state. Peptide probabilities were calculated and set to >90% C.I. with the number of peptides/protein set at 2 or more and protein probabilities set to >99% C.I.
Co-immunoprecipitation
For FGFR-1 immunoprecipitation, lysates were prepared using methods described by Kurosu and colleagues.(9) For Wnt7a IP, lysates were prepared as above. Precleared lysate was incubated with 10 μL of one of the primary antibodies: KL-115 (0.9 mg/mL), KL-234 (1:100, 1.2 mg/mL), myc (SC789, Santa Cruz Biotechnology), FGFR-1 (SC7945, Santa Cruz Biotechnology), or wnt7A (SC26361, Santa Cruz Biotechnology). Rat serum diluted to the same concentration as primary antibody was used as negative control. Protein G agarose was added to pull down protein complexes. Beads were washed two times in PBS. For Western blotting, washed beads were suspended in an equal volume of 5x SDS sample loading buffer, boiled, and supernatant loaded onto 10% polyacrylamide gels. Immunoblots were processed as above.
Results and Discussion
Generation of KL hybridoma lines and initial screening
To generate monoclonal antibodies, we constructed a plasmid to allow induction of recombinant protein for injection. This was accomplished by fusing the first half of the KL1 domain of KL in-frame with glutathione-S-transferase (GST) (Fig. 1A). Targeting this region of KL (amino acids 55-261) has produced specific antibody in the past(16) and will theoretically allow detection of all KL protein forms: transmembrane, shed, and secreted. Successful IPTG induction of the fusion protein was confirmed following Coomassie stain of the polyacrylamide gel. GST-KL1 fusion protein concentrations were determined by estimation to BSA standard proteins (Fig. 1B). Following immunization, fusion of sensitized lymph node cells with P3U-1 myeloma cells generated 384 hybridomas. Hybridoma supernatants were tested for reactivity to GST-KL and GST alone via ELISA. Forty-four supernatants ELISA, positive for GST-KL but negative for GST alone, were further screened in Western blot, ICC, and IHC applications. KL-234 and KL-115 were selected for scale up, antibody purification, and further validation.
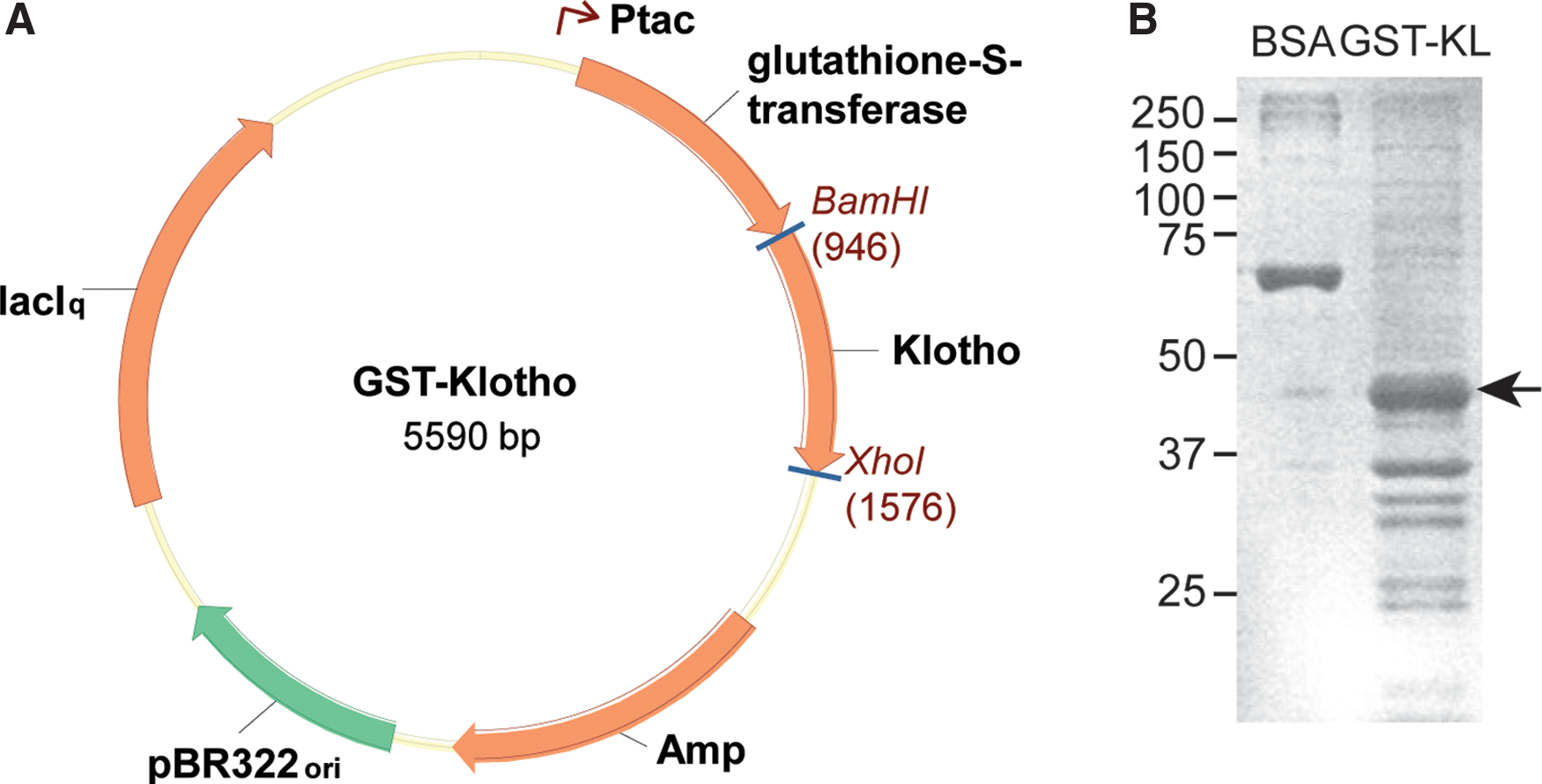
GST-KL1 cloning and recombinant protein production. (
Detection of KL protein across experimental applications
The most significant drawbacks in utilizing commercially available antibodies are restricted species reactivity, a lack of specificity when comparing wild-type to knockout tissue, and/or utility in only one experimental application. To validate KL-234 and KL-115 specificity, utility across species, and experimental applicability, we tested their function in Western blot, ICC, and IHC protocols. HEK293T cells were transfected with myc-tagged plasmid constructs for mouse KL, human KL, or a control plasmid (pcDNA 3.1). Following Western blot of lysates, myc-tagged mouse KL was detected, as anticipated, at 130 kDa using a myc antibody (Fig. 2A). When KL-115 was used as the primary antibody, transfection of human but not mouse KL was detected (Fig. 2A, panel 2). KL-234 detected both human and mouse KL from transfected cells (Fig. 2A, panel 3). Tubulin confirmed equal protein loading across transfected conditions (Fig. 2A). As cell transfection represents overexpression, detection of endogenous protein level was analyzed by Western blot of mouse kidney lysates using KL-234. KL-234 detected KL in wild-type but not knockout kidney lysates and detected equivalently to mouse KL transfection control lysate (Fig. 2B). Only after prolonged exposure times of more than 30 min were any non-specific bands appreciable (data not shown). We next compared use of KL-234 to two other antibodies typically used to detect KL, KM2076 and AF1819. Using KL-234 a band was detectible when 10 μg total protein were loaded. AF1819 did not detect KL until at least 50 μg of total kidney was loaded (Fig. 2C; 30 μg for KL-234; AF1819, 50 μg). Even at 100 μg, no band was detected with KM2076 (data not shown).
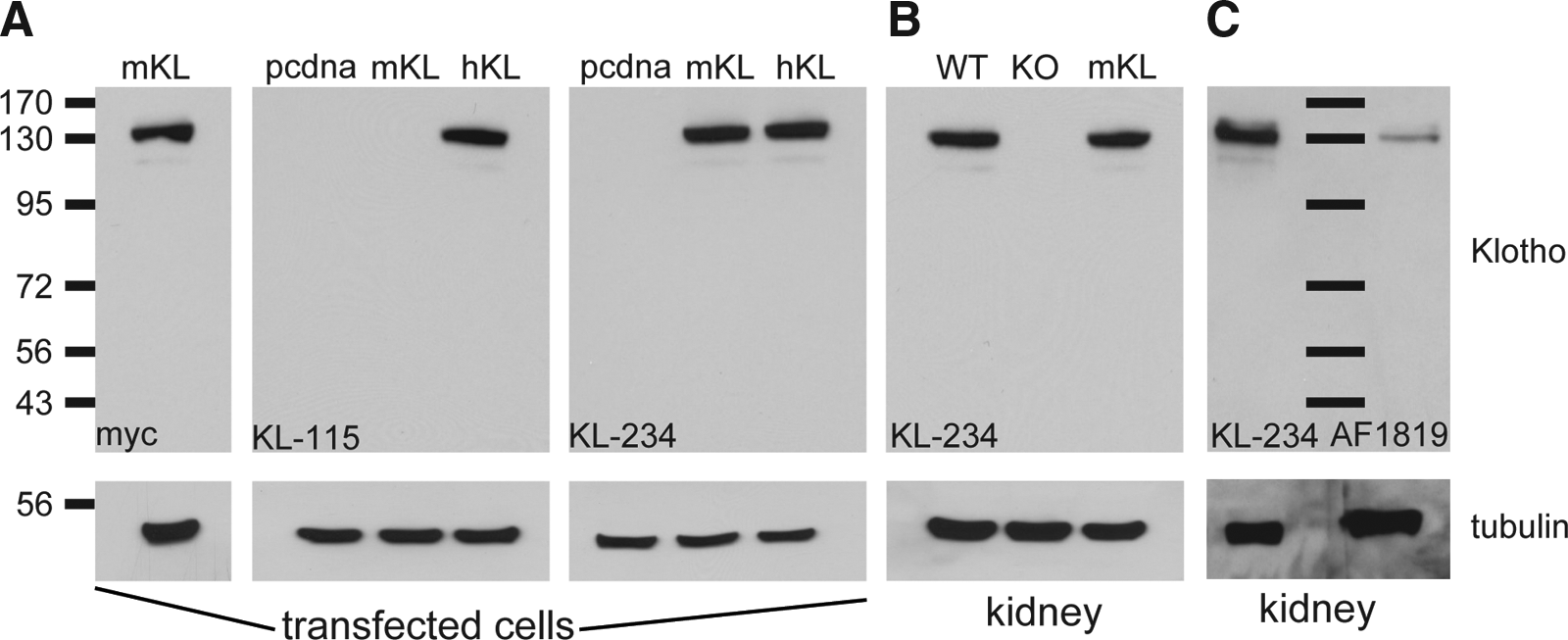
KL-115 and KL-234 differentially detect human and mouse KL protein by Western blot. Representative Western blots from: (
KL is detected most highly in the convoluted tubules of the kidney and the choroid plexus of the brain. In our hands, the only antibody able to detect endogenous levels of KL by IHC is AF1819 and detection requires multiple levels of TSA mediated antibody amplification, particularly in organs with lower protein expression levels.(19) ICC of transfected HEK293T cells revealed that both KL-234 and KL-115 detect mouse and human KL under overexpressing conditions (Fig. 3A). IHC using kidney and brain sections revealed that KL-234 detected endogenous protein equivalently to AF1819 and required less TSA amplification for this equivalent detection (Fig. 3B). No reactivity was noted in control conditions or in knockout kidney samples (Fig. 3B). Low level KL expression was detected in knockout choroid plexus consistent with observations of incomplete knockout in brain tissues (Fig. 3B).(1,19) KL-115 did not detect mouse KL in wild-type kidney or brain sections by IHC (data not shown). Although KL-115 may be capable of detecting mouse KL depending on expression level, KL-234 appears more versatile, detecting mouse and human in both Western and IHC applications.
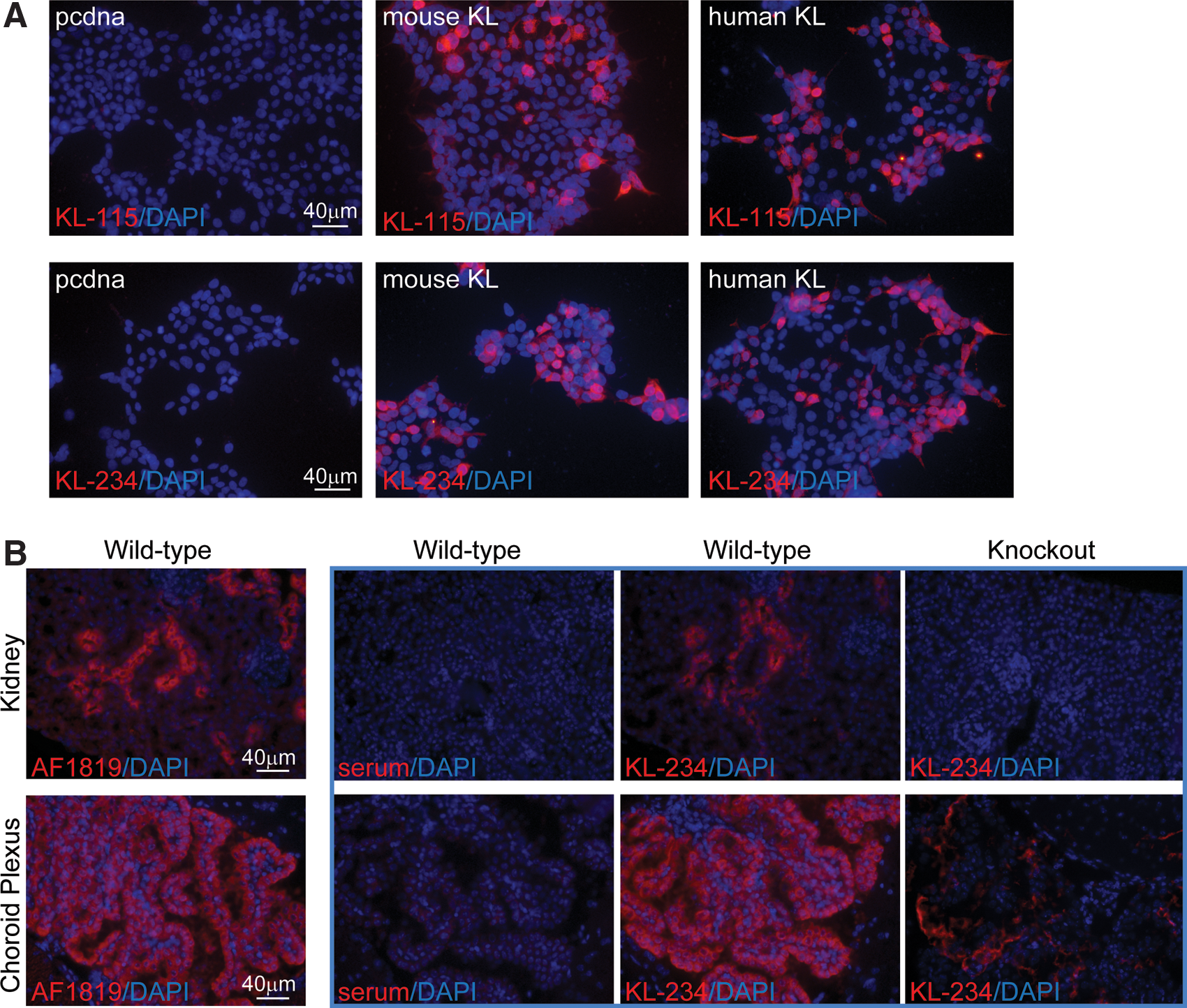
KL-115 and KL-234 detect human and mouse KL by immunocyto/histochemistry. (
While antibody specificity is generally accepted as valid when based upon Western blot and/or IHC comparing wild-type and knockout tissues, we further validated antibody specificity with immunoprecipitation and mass spectrometry. Both KL-115 and KL-234 were able to detect transfected KL following IP of myc-tagged KL, confirming our Western blot results (Fig. 4A). In reciprocal IP experiments, when either KL-115 or KL-234 antibody was used as the IP reagent, myc-tagged KL was pulled down (Fig. 4B). Utilizing lysates from cells transfected with human KL plasmid, when the products of IP KL-115 or IP KL-234 were subjected to mass spectrometry, human KL was confirmed to be pulled down with peptides identified covering, respectively, 41% and 56% of KL (Fig. 4D). Control IP did not detect any peptides identified as KL.
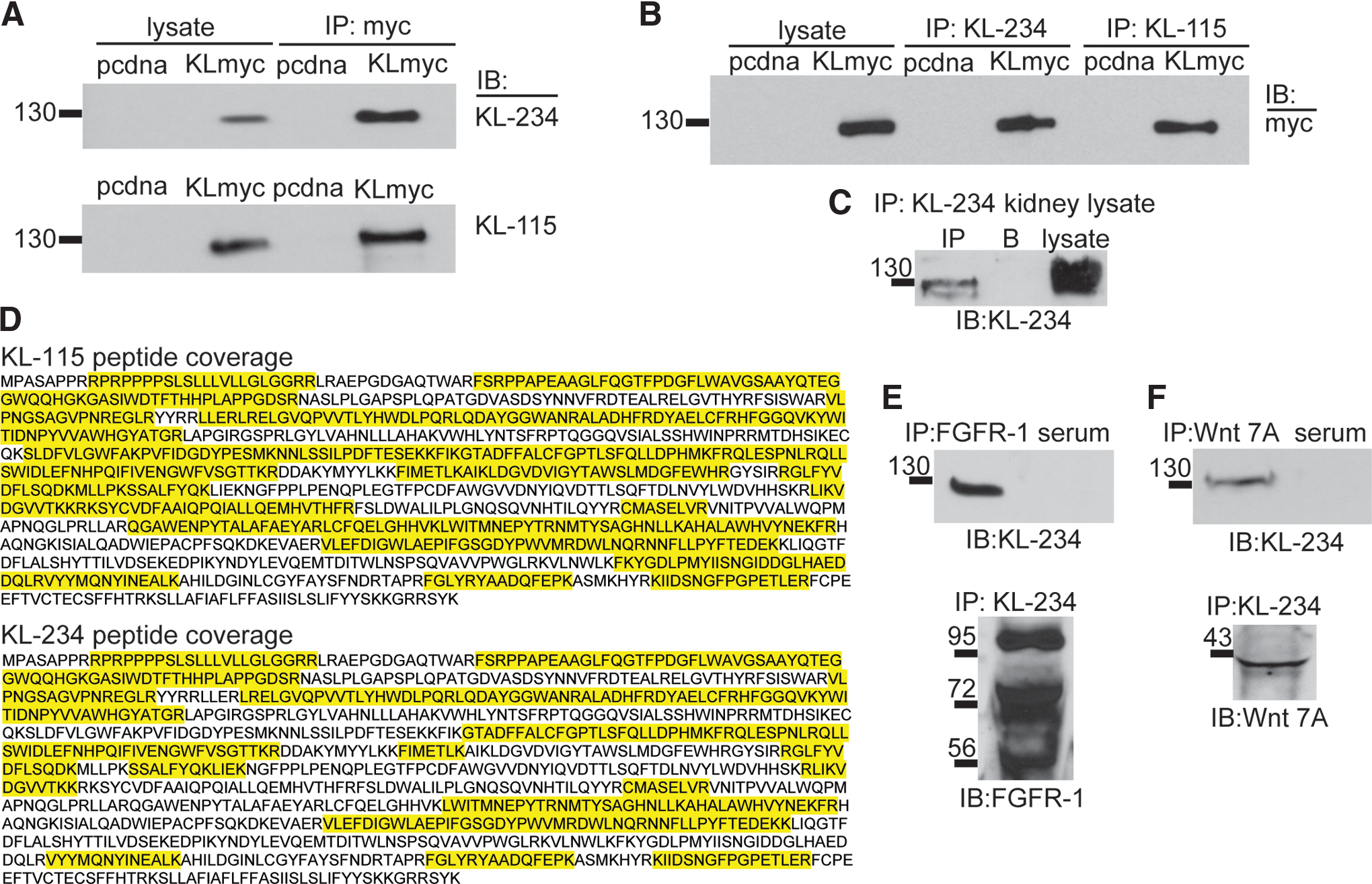
KL-115 and KL-234 immunoprecipitate KL. Both KL-115 and KL-234 antibodies were tested for use in immunoprecipitation (IP) protocols. Representative IP/Western blots are shown. (
KL functions by interacting with other protein partners; most notably KL is the co-receptor with FGFR to transduce FGF23 signaling and KL binding to wnt proteins inhibits wnt signaling.(7,9) Although a paper confirmed an in vivo interaction between KL/FGFR,(9) most identification and subsequent follow-up have required the use of protein tags. We verified that KL-234 could IP under endogenous conditions utilizing lysates from kidney. IP and detection with KL-234 allowed pull down of endogenous KL (Fig. 4C). Since KL-234 antibodies can IP endogenous levels of KL protein, we sought to confirm KL/FGFR and KL/wnt interactions from kidney lysates. FGFR-1 antibody IP pulled down KL protein and KL-234 antibody IP of KL pulled down FGFR-1 protein (Fig. 4E). Wnt7A antibody IP pulled down KL protein and KL-234 antibody IP of KL pulled down Wnt7A protein (Fig. 4F).
We have generated two new antibodies with unique KL detection profiles. KL-115 is specific to detection of human KL, although it can detect mouse KL by ICC when overexpressed. KL-234 detects human and mouse KL in Western blot, ICC/IHC, and immunoprecipitation applications enabling validation and expansion of our understanding of KL and its binding partners in vitro and in vivo.
Author Contributions
AM conducted initial hybridoma screening and participated in validation by Western blot, ICC, and IP. AN participated in initial characterization. AL participated in fusion protein production. TSD participated in GST-KL vector construction. MEB designed the GST-KL, oversaw production, and assisted in fusion protein production. MAL generated developed immunization schemes, generated hybridomas, and conducted initial ELISA screens. GDK oversaw all aspects of the project, conducted Western blot and IHC experiments, and wrote and edited the manuscript.
Footnotes
Acknowledgments
We gratefully acknowledge the support of core facilities at UAB, including the UAB NINDS Protein Interaction Core (P30NS047466), the UAB Comprehensive Cancer Center – Mass Spectrometry/Proteomics Shared Facility (P30CA13148-38), and the UAB Molecular Detection Core (P30 NS047466). This study was supported by NIH/NIA R00 AG034989 (to GDK).
Author Disclosure Statement
The authors have no financial interests to disclose.