
Abstract
Abstract
Bianba, Sveinung Bernsten, Lars Bo Andersen, Hein Stegum, Ouzhuluobu, Per Nafstad, Tianyi Wu, and Espen Bjertness. Exercise capacity and selected physiological factors by ancestry and residential altitude—Cross-sectional studies of 9–10-year-old children in Tibet. High Alt Med Biol. 15:162–169, 2014.—Aim: Several physiological compensatory mechanisms have enabled Tibetans to live and work at high altitude, including increased ventilation and pulmonary diffusion capacity, both of which serve to increase oxygen transport in the blood. The aim of the present study was to compare exercise capacity (maximal power output) and selected physiological factors (arterial oxygen saturation and heart rate at rest and during maximal exercise, resting hemoglobin concentration, and forced vital capacity) in groups of native Tibetan children living at different residential altitudes (3700 vs. 4300 m above sea level) and across ancestry (native Tibetan vs. Han Chinese children living at the same altitude of 3700 m).
Methods:
A total of 430 9–10-year-old native Tibetan children from Tingri (4300 m) and 406 native Tibetan- and 406 Han Chinese immigrants (77% lowland-born and 33% highland-born) from Lhasa (3700 m) participated in two cross-sectional studies. The maximal power output (Wmax) was assessed using an ergometer cycle.
Results:
Lhasa Tibetan children had a 20% higher maximal power output (watts/kg) than Tingri Tibetan and 4% higher than Lhasa Han Chinese. Maximal heart rate, arterial oxygen saturation at rest, lung volume, and arterial oxygen saturation were significantly associated with exercise capacity at a given altitude, but could not fully account for the differences in exercise capacity observed between ancestry groups or altitudes.
Conclusions:
The superior exercise capacity in native Tibetans vs. Han Chinese may reflect a better adaptation to life at high altitude. Tibetans at the lower residential altitude of 3700 m demonstrated a better exercise capacity than residents at a higher altitude of 4300 m when measured at their respective residential altitudes. Such altitude- or ancestry-related difference could not be fully attributed to the physiological factors measured.
Introduction
A
Most previous studies have shown that native Tibetan adults (Sun et al., 1990; Groves et al., 1993; Zhuang et al., 1993; 1996) and adolescents (Chen et al., 1997) achieve a superior exercise capacity compared to Han Chinese immigrants. However, a few studies have shown that native Tibetan adults attained a higher exercise capacity, but a similar (Niu et al., 1995) or lower (Ge et al.,1994) peak oxygen uptake (
A person who moves from sea level to Lhasa (3700 m) will acclimatize after some days, but even after decades he or she will not reach a sea level performance of exercise or work. A person arriving in Lhasa from sea level can expect to reach an exercise performance or work that is approximately 70% of his or her performance at sea level, which will increase to 80% after a few weeks of acclimatization (Moore et al., 1994). Similar to the immigrant Han Chinese in Tibet, if a person stays at 3700 m for years, there may be a further small increase to nearly 85% of the initial sea level value (Moore et al., 1994). After generations of residence, Tibetans at 3700 m have an exercise performance or work that is 92% of sea level values for sedentary populations (Moore et al., 1994).
In relation to both a decrease in arterial oxygen content and a limitation in maximal cardiac output, moving lowlanders to higher altitudes is associated with a decreased aerobic exercise capacity (Fulco et al., 1998), while a higher pre-exposure residential altitude modulates the negative effect (Fulco et al., 1998). High altitude populations are continuously challenged in an environment of hypobaric hypoxia with a low ambient partial pressure of oxygen, thus influencing the capacity for work and exercise. Moreover, the capacity is reduced with an increasing residential altitude (Fulco et al., 1998). However, if high altitude natives move to a lower altitude, they will exhibit a better exercise capacity than natives living at the same, lower altitude, as demonstrated by Curran et al. (1998). The relatively better maximal exercise capacity in high altitude natives may primarily be due to their exposure to a high altitude environment during childhood (Frisancho et al., 1973). Independent of being a child or an adult, or of a low or high socioeconomic status, everyone is exposed to the same ambient hypoxia at a given altitude. As reviewed by Simonsen et al. (2012), native highland populations have genetic adaptations that enable them to live at high altitudes. Three out of several genes associated with natural selection among Tibetans, EPAS1, EGLN1, and PPARA, are associated with their relatively low hemoglobin concentration (Simonson et al., 2012).
In a review by Wu and Kayser (2006), they concluded that among adults, Tibetans were better adapted than other populations to live and work at high altitudes. In addition to a lower Hb concentration, as compared with lowlanders and Andeans living at the same altitude, several other physiological factors contributed to this adaptation, including: maintaining a higher arterial oxygen saturation at rest and during exercise, less loss of aerobic performance with an increasing altitude, a greater hypoxic and hypercapnic ventilatory responsiveness, larger lungs, a better lung function, and a greater lung diffusing capacity than lowlanders (Wu and Kayser, 2006). Lastly, Tibetans develop only minimal hypoxic pulmonary hypertension (Groves et al., 1993), and have higher levels of exhaled nitric oxide than lowlanders or Andeans (Wu and Kayser, 2006). Studies of physiological aspects of Tibetan children's capacity to work and live at high altitude are scarce, although differences similar to those seen in adults may be expected to occur. Children living at altitudes higher than 3000 m above sea level show several adaptive mechanisms in the uptake of oxygen in terms of increased ventilation and pulmonary diffusion capacity (de Meer et al., 1995). The latter depends on the diffusion rate per unit of the surface area and the total surface area in the lungs available for diffusion of oxygen, which is reflected in part by total lung capacity (de Meer et al., 1995). Oxygen transportation in the blood is highly dependent on blood flow, hemoglobin concentration ([Hb]), and arterial oxygen saturation (Sa
The objectives of the present study were to compare exercise capacity (maximal power output) and selected physiological factors (arterial oxygen saturation and heart rate at rest and during maximal exercise, resting hemoglobin concentration, and forced vital capacity) in groups of high altitude native Tibetan children across residential altitudes (3700 vs. 4300 m above sea level) and across ancestry (native Tibetan vs. Han Chinese children living at the same altitude of 3700 m). We also aimed to investigate the association of residential altitude and ancestry with exercise capacity, and the possible impact of selected physiological factors on this association.
Materials and Methods
Nine primary schools out of 20 were randomly selected from Lhasa City (3700 m), the capital of TAR, while all five primary schools were included from the rural Tingri district (4300 m) located under the North Face of Mt. Everest. An identical procedure for selecting participants was applied in both areas, and all 9–10-year-old children, 817 children from Lhasa and 460 children from Tingri, were invited. In Lhasa, one child refused to participate and four children on the list were excluded due to respiratory health problems, yielding 812 eligible children, which gave a participation rate of 99%. A total of 406 native Lhasa Tibetan children (207 boys and 199 girls) and 406 Lhasa Han Chinese children (235 boys and 171 girls) participated in the urban area. Among the Han Chinese, 313 (77%) were born at low altitude (lowland-born Han Chinese) and then migrated to Lhasa with their parents, while the remaining 93 (23%) were born in Lhasa (highland-born Han Chinese). In Tingri, 16 did not go to school on the days of the data collection and four children refused to participate, whereas 10 children participated but did not give permission to use their data for publication, thus yielding a response rate of 93%. A total of 215 girls and 215 boys participated from Tingri.
Ethical considerations
The Health and Education Office and the Tibet University Medical College in TAR approved the study, and information and consent forms were given to parents through the school leaders. Information about the study procedure was also given to the children, participation was voluntary, and the children could withdraw from the study for any reason at any time with no negative consequences.
Data collection and variables
Data were collected through a questionnaire, a clinical examination, and a maximal exercise test, and the same research team and methods were used in the studies in Lhasa and Tingri. The data collections were performed in a classroom at each school from August to November 2005 in Lhasa, and from September to October 2007 in Tingri. In addition, there was no vigorous physical activity the day before the test, no food was consumed less than 2 hours prior to the test, and no feast meals were eaten less than 4 hours before each test.
Study group
“Study group” is a variable that combines ancestry and residential altitude, which includes Tingri Tibetans, Lhasa Tibetans, and Lhasa Han Chinese. It is a measure of the effect of altitude when comparing Lhasa Tibetans (3700 m) and Tingri Tibetans (4300 m), and a measure of ancestry effect when comparing Lhasa Tibetans and Lhasa Han Chinese. Lhasa Han Chinese were further divided into lowland- and highland-born for selected analyses.
Maximal power output and heart rate
Maximal power output (Wmax) was assessed using an electronically braked cycle ergometer (Monark Ergomedic 839, Varberg, Sweden) and presented as exercise capacity according to a previously validated protocol (Hansen et al., 1989). We lack data on 350 children from the cycle ergometer test: 279 children could not fully reach the pedals, 49 children could not complete the test, and for 22 no reason was given.
Children were requested to have 5 minutes of practice on the ergometer cycle before the test to ensure that all inexperienced children had some cycling practice. A variable “owning a bicycle,” with answers of yes or no, was used to distinguish between experienced and inexperienced cyclists. Heart rate (Polar Electro OY, Kempele, Finland) was measured throughout the test, and recorded at the end of each step in the progressive cycle test. We applied methods similar to previous studies in children (Aandstad et al., 2006; Riddoch et al., 2005), and more details regarding the procedures and test criteria can be found elsewhere (Bianba et al., 2010). The children cycled at a pedaling rate of 70–80 revolutions per minute (rpm), and the power output was increased by 20 W (weight<30 kg) or 25 W (weight>30 kg) every third minute. The test was stopped when the child could no longer continue with a pedaling rate above 30 rpm.
Wmax was determined as the number of watts in the last fully completed step (Wl), plus the increment in watts (Wd) of the last step, multiplied by the number of seconds completed at the last step (t) and then divided by 180 seconds (Hansen et al., 1989): Wmax=Wl + (Wd · t /180). The absolute Wmax and Wmax relative to body mass (Wmax/body mass) are presented.
Arterial oxygen saturation
Arterial oxygen saturation at rest (Sa
Hemoglobin concentration
A HemoCue Hb 201+ analyzer (Ängelholm, Sweden) was applied to measure [Hb] in capillary blood, and we calibrated the analyzer every testing day using a HemoCue Hemoglobin Calibrator (12.0±0.2 g•dL−1).
Lung volume
As a measure of total lung capacity, forced vital capacity (FVC) was measured using maximal expiratory flow volume maneuvers with Spiro USB (Micro Medical Limited, Rochester, Kent, UK), according to standardized international guidelines (Miller et al., 2005). The highest of the recorded FVC values was reported in the present study.
Anthropometrics
The body mass of the children was measured to the nearest 0.1 kg (without shoes and wearing light clothes) using an Electronic Scale (OMRON, HN-281, Shanghai, China), while height was measured to the nearest 0.5 cm using a stadiometer (TZG, Shanghai, China). Chest and waist circumferences were measured to the nearest 0.5 cm with children standing and breathing normally, and the average value both before and after expiration was recorded. The Body Mass Index (BMI) was calculated as the body mass (in kg) divided by the height squared (in meters).
Statistical methods
Data are reported as the mean plus or minus one standard deviation or as the mean with a 95% confidence interval. Differences in mean values between groups were tested using a one-way ANOVA. Three linear regression models were constructed for determining the relationship between the outcome variable (maximal power output) and the selected covariates. The association between the “study group” (i.e., residential altitude and ancestry) and Wmax was estimated in Model 1, with adjustment for sex, body mass, and having a bicycle. In Model 2, we estimated the association between physiological covariates ([Hb], Sa
Results
The sex-specific descriptive data of Tingri Tibetan, Lhasa Tibetan, and Lhasa Han Chinese children are presented in Table 1. On average, the Lhasa Tibetans were 32% and 7% heavier than Tingri Tibetans and Lhasa Han Chinese, respectively. Furthermore, they were 10% and 1% taller, and had 8% and 4% larger chest circumferences than Tingri Tibetans and Lhasa Han Chinese, respectively.
p<0. 05 Tingri Tibetans vs. Lhasa Tibetans, **p<0. 001 Tingri Tibetans vs. Lhasa Tibetans.
p<0. 05 Tingri Tibetans vs. Lhasa Chinese,##p<0. 001 Tingri Tibetans vs. Lhasa Chinese.
p<0. 05 Lhasa Tibetans vs. Lhasa Chinese,□□p<0. 001 Lhasa Tibetans vs. Lhasa Chinese.
Physiological factors
The [Hb] was 5% higher in Lhasa Han Chinese compared with Lhasa Tibetans, and 4% higher in Lhasa Tibetans than in Tingri Tibetans. The lung volume (FVC) and peak heart rate (HRpeak) were higher in Lhasa Tibetans than both Lhasa Han Chinese and Tingri Tibetans. Lhasa Tibetan achieved 46% and 10% higher levels of absolute Wmax (W) than Tingri Tibetan and Lhasa Han Chinese, respectively, and 20% (21.8% in boys and 18.5% in girls) and 4% (5.8% in boys and 2.1% in girls) higher levels of relative Wmax (W/kg), respectively (Table 1).
There was no difference in arterial oxygen saturation at rest (Sa
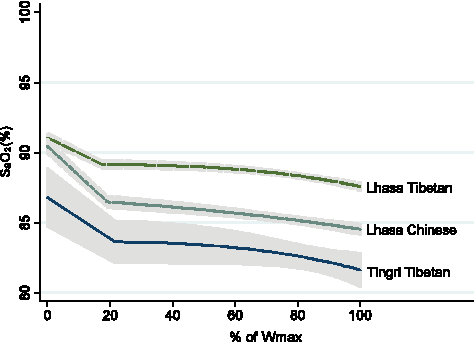
Alterations in arterial oxygen saturation (Sa
Relationship between maximal power output (Wmax) and selected physiological factors
Coefficients from linear regression analyses are presented in Table 2, and in Model 1, Wmax for Lhasa Tibetans was 86 W (95% confidence interval: 83.2, 88.2). The effect of residential altitude on expected Wmax was estimated by contrasting the two study groups, Tingri vs. Lhasa Tibetans, yielding a 16 (−18.8, −14.0) W lower Wmax in Tingri Tibetans. The effect of ancestry was estimated by contrasting Lhasa Han Chinese vs. Lhasa Tibetans, yielding a 5 (−6.4, −2.9) W lower Wmax in Lhasa Han Chinese. These results indicate that Tibetans living at a higher altitude had a lower Wmax, and Han Chinese had a lower Wmax than the Tibetans at the same altitude. Model 2 shows that all the included physiological variables were associated with Wmax. For example, for every increase in maximal heart rate of one beat per minute, the exercise capacity (maximal power output, Wmax) increased by 0.4 W, and for a one liter increase in lung volume (FVC), the Wmax increased by 13.2 W, whereas a one kg body mass increase was associated with a 1 W increase in Wmax (Table 2, Model 2). Girls had an 8.8 W lower Wmax than boys (Table 2, Model 2), and the explained variance (R2) of Model 2 was 54%. The estimated Wmax was 79 W when the following were set as a reference: a boy with a body mass=27 kg; [Hb]=14.6 g/L; HRpeak=195 beats/min; Sa
No bivariate association was found between HRrest and a maximal power output, thus not included in Models; ▴Own a bicycle (vs. no bicycle) was set as reference; ▵when boy is set as reference and body mass=27 kg, [Hb]=14.6g/L, HRpeak=195 beats/min, SaO2rest=90%, SaO2peak=86%, FVC=2.00; *0.001<p<0.01; **p<0.001;
 The explained variance (R2) is 54%;
The explained variance (R2) is 63%.
The explained variance (R2) is 54%;
The explained variance (R2) is 63%.
Discussion
In the present study we have shown that native Lhasa Tibetan 9–10-year-old children presented a higher exercise capacity than both native Tingri Tibetans and Lhasa Han Chinese when measured at their respective residential altitudes. [Hb], Sa
Compared to Lhasa Tibetan children living at 3700 m, the lower maximal power output in native Tingri Tibetans living at 4300 m may be due to the reduction in the inspired partial pressure of oxygen (PiO2). Furthermore, children's growth and development are closely correlated with their work and exercise performance, as well as altitude adaptation (Frisancho et al., 1975; 1985). The present study reveals that Tingri Tibetan children were shorter and lighter than Tibetan children from Lhasa of the same age, and such findings are consistent with a previous comparative study of Tibetan children and adolescents aged 7–18 years living at various altitudes from 2261 m to 4040 m in Qinghai-Tibet. The children from the highest altitude (4040 m) presented a slower growth (stature), lagging behind their counterparts at 2261 m by approximately 2 years. It was also indicated that hypoxia may be the main agent responsible for this delayed growth (Zhang et al., 1985), and Greksa et al. (1984) suggested that socioeconomic factors slowed the growth of high-altitude children (Greksa et al., 1984). In the present study, we have no valid information on socio-economic status (SES). It is a challenge to identify an indicator of SES that is valid for both Lhasa and Tingri. For example, in Tingri, parents generally have a low or no education and no income, but they may have many Yak or sheep, thus indicating a high SES. In contrast, in Lhasa, parents seldom own animals, and most males have an education, as well as income.
We speculate that the present finding of a lower [Hb] among Tingri Tibetan children compared with Lhasa Tibetan children, which stands in contrast to previous reports of an increase in [Hb] with an increasing altitude (Wu et al., 2005), may be due to poor nutrition, including less iron-rich foods at high altitude.
The present finding of a higher maximal power output in native Tibetan- compared to Han Chinese children at the same altitude may be due to a better oxygen transport system. The difference in oxygen concentration between arterial- and mixed-venous blood is a measure of tissue oxygen extraction, which can be inferred from [Hb] and the difference between the oxygen saturation in arterial- and mixed-venous blood (Sa
Although the present Lhasa Tibetan children also had a lower [Hb] than Han Chinese, they sustained a higher Sa
A large sample size and high participation rates are strengths of the present study, while a weakness is that the study could have underestimated the exercise capacity for children who were less accustomed to cycling. However, the adjustment by the variable “owning a bicycle” in Models 1–3 may have partly compensated for differential cycling practices across the three groups of children. A selection of healthier children for the schools in rural Tingri may have taken place because of the need to be sufficiently physically healthy to walk the long way to school, and to live at the school dormitory for long periods. This is in contrast to urban Lhasa children, who are staying with their families, and have short travel distances. As a result, this possible selection would most likely have distorted the results towards a smaller difference between Tingri- and Lhasa children.
Conclusion
The superior exercise capacity in native Tibetan children vs. Han Chinese immigrant children living at the same altitude of 3700 m may reflect a better adaptation to life at high altitude. The lower exercise capacity at a higher altitude in native Tibetans may partially be attributed to residential altitude differences. In addition to residential altitude and ancestry, selected physiological parameters explained some of the variance in exercise capacity, including higher arterial oxygen saturation and a higher forced vital capacity, but these physiological factors did not fully account for the associations between residential altitude and ancestry with a maximal power output. Future studies comparing Tibetan and Han Chinese children living at the same altitude should also take genetic factors into account, in addition to potentially unmeasured confounders such as SES and food habits. These confounders should also be included in future studies comparing native Tibetans living at different altitudes.
Footnotes
Acknowledgments
We are very thankful to the Network for University Cooperation Tibet-Norway for supporting this study. Thanks too to all the children who participated in this study and to our colleagues at the Tibet University Medical College, who gave great support in the data collection.
Author Disclosure Statement
Drs. Bianba, Bernstsen, Andersen, Stigum, Ouzhuluobu, Nafstad, Wu, and Bjertness have no conflicts of interest or financial ties to disclose.