
Abstract
Metallothioneins (MTs) are a group of intracellular metal-binding and cysteine-enriched proteins and are highly inducible in many tissues in response to various types of stress. Although it mainly acts as a regulator of metal homeostasis such as zinc and copper in tissues, MT also acts as a potent antioxidant and adaptive (or stress) protein to protect cells and tissues from oxidative stress. Diabetes affects many Americans and other populations, and its development and toxic effect on various organs have been attributed to increased oxidative stress. Studies showed that zinc-induced or genetically enhanced pancreatic MT synthesis prevented diabetes induced by chemicals such as streptozotocin and alloxan, and zinc pretreatment also prevented spontaneously developed diabetes. Since diabetic complications are the consequences of organ damage caused by diabetic hyperglycemia and hyperlipidemia through oxidative stress, whether MT in nonpancreatic organs also provides a preventive effect on diabetic toxicity has been recently investigated. We demonstrated that overexpression of cardiac MT significantly prevented diabetes-induced cardiomyopathy. Likewise, overexpression of renal MT also prevented diabetes-induced renal toxicity. In addition, we also found that MT as an adaptive protein is overexpressed in several organs in response to diabetes. Therefore, the biological importance of diabetes-induced MT in diabetic complications and subsequent other pathogenesis was further explored. We found that diabetes-induced hepatic and renal MT synthesis was accompanied by a significant prevention of endotoxin-induced hepatic toxicity and cisplatin-induced renal toxicity. These studies suggest that MT as an adaptive protein can prevent both diabetes development and its complications or subsequent suffered other pathogenic injury.
INTRODUCTION
Metallothioneins (MTs) are a group of intracellular metal-binding and cysteine-enriched proteins. Four isoforms of MTs are characterized, and MT-I and MT-II exist as the major isoforms in various human and animal organ tissues including pancreas (Thornalley and Vasak, 1985; Klaassen and Liu, 1998; Cai et al., 1999; Kang, 1999). Zinc (Zn) is the major metal to which MT binds under physiological conditions, but is can be replaced by other metals such as copper (Cu), cadmium (Cd), and iron (Fe). Functionally, MT mainly acts as a regulator of metal homeostasi in tissues, especially trace element metals, and also acts as a potent antioxidant to protect cells or tissues from oxidative damage (Klaassen and Liu, 1998; Cai et al., 1999; Kang, 1999). Compared with other antioxidants that can protect specifically against certain damage—SOD against superoxide radical, catalase against hydrogen peroxide, and glutathione peroxidase against hydrogen peroxide and lipid peroxides—MT is a potent antioxidant against a wide range of free radicals including the most active radicals: hydroxyl radical and peroxynitrite (Thornalley and Vasak, 1985; Miura et al., 1997; Cai et al., 1999, 2000a; Cai and Cherian, 2003).
Diabetes affects so many Americans and other populations around the world. Cardiovascular injury from diabetes contributes greatly to various diabetic complications including renal, retinal, and cardiac dysfunction and even stroke (Cai and Kang, 2001a; Rosen et al., 2001). In addition, both diabetes onset and its complications have been considered to be due to the generation of oxidative stress caused by diabetes; therefore, developing a strategy for prevention of diabetic onset and its complications through suppression of oxidative stress has received investigative attention (Cai and Kang, 2001a; Rosen et al., 2001). A pilot study by Dr. Cherian has demonstrated that Zn-induced MT synthesis in rat pancreas prevented chemical streptozotocin (STZ)-induced diabetes (Yang and Cherian, 1994), and we have shown that overexpressed MT in the mouse heart significantly prevented diabetes-induced cardiomyopathy (Cai and Kang, 2001b; Kang and Cai, 2001). These preliminary studies have been further confirmed and characterized by later studies. In this article, the summarized data based on our own and other studies for the prevention of diabetes onset, its complications, and subsequent suffered pathogenic injury were presented to extend our insight into MT prevention of diabetes and its toxicity.
MATERIALS AND METHODS
Early published data on the preventive effect of MT on diabetes onset and diabetic complications were obtained by searching MEDLINE using MT and diabetes and selected after studying the content of these publications. Our own studies were performed in a mouse diabetic model which was produced by giving a single injection of 150 mg/kg STZ as described by Cai et al. (2002a).
RESULTS AND DISCUSSION
MT Prevents Diabetes
In the study by Yang and Cherian (1994), diabetes was produced by a single injection of STZ (75 mg/kg) in Sprague Dawley rats, and the synthesis of pancreatic MT was induced by subcutaneous Zn injection 12 hr before injection of STZ. Results, summarized in Figure 1, indicate that both Zn and MT levels increased in the pancreas of rats on day 1 after STZ treatment. Serum MT was also significantly increased in the Zn + STZ group on days 1, 21, and 45 after STZ treatment. More importantly, serum glucose levels in both STZ and Zn + STZ groups increased compared with the control group, but the level of glucose in the Zn + STZ group was significantly lower than that in the STZ group (Figure 1D). This result suggests that MT induction by Zn pretreatment could significantly prevent STZ-induced diabetes. Subsequent experiments supporting the case for prevention against diabetes development by MT are summarized in Table 1 with the following characteristics.
Evidence for the Preventive Effect of MT on Diabetes

Notes: ND, no detected; IS, immunohistochemical staining; WB, Western blotting; w, week; Cd-hem, 109Cadmium-hemoglobin method to measure MT protein; 1 (12) w, 1 week prior to STZ and continued 12 weeks after STZ.

Changes of Zn and MT in tissues, and serum MT and glucose. Rats were treated by Zn for 12 hr and then treated with STZ. At day 1 (30 hr) after STZ, (A) Zn and (B) MT levels were measured. In addition, (C) serum MT and (D) glucose levels were also measured on days 1, 21, and 45 after STZ treatment. a: p < 0.05 or 0.01 vs control; b: p < 0.05 or 0.01 vs STZ alone. This graph was modified based on the results of a study by Yang and Cherian (1994) with permission from the authors.
Zn supplement was the most commonly used agent to induce pancreatic MT synthesis for prevention of the diabetic development. Supplementation of Zn by subcutaneous or intraperitoneal injection, drinking water and dietary food were all effective in inducing pancreatic MT, which was confirmed by measuring MT protein level using biochemical assay (such as Cd-hem), Western blotting or immunohistochemical localization, and mRNA level using the RT-PCR method. By in situ immunohistochemical methods, MT induction has been specifically identified in islet β-cells. Cu is another well-known MT inducer (Thornalley and Vasak, 1985; Klaassen and Liu, 1998; Cai et al., 1999; Kang, 1999). Like Zn, supplementation of Cu also provided significant prevention of diabetes induced by multiple low doses of STZ (Sitasawad et al., 2001). In addition, a transgenic mouse model was developed in which MT is pancreatic-specifically expressed about 20-fold higher than the wild type without alteration of other antioxidants, and a significantly high resistance to STZ-induced diabetic development was observed in this mouse model (Chen et al., 2001), suggesting the direct prevention by MT of STZ-induced diabetes.
Induced MT synthesis prevents diabetes development in a wide range of diabetic models. It has been recently reported that multiple low doses of STZ induce a diabetic model mechanistically different from that induced by a single high dose of STZ (Ohly et al., 2000; Ho et al., 2001; Sitasawad et al., 2001). Induced MT synthesis in pancreas not only prevented diabetes induced by a single dose of STZ or alloxan, but it also prevented diabetes induced by multiple low doses of STZ, and even genetically prodiabetic models such as the BB Wister rat and db/db or od/od mouse diabetes development. This suggests that MT can prevent both type I and type II diabetes.
The preventive effect of T on diabetes could be seen in different strains of rats and mice. The preventive effect of induced MT synthesis in pancreas was not affected by species difference for the diabetic development in response to STZ. For example, C57BL/6 mice typically start severe hyperglycemia 2 weeks after the first injection of STZ, whereas B6SJL/F1 mice show a long latent period for developing severe hyperglycemia about 10 weeks after the first injection of STZ (Ohly et al., 2000). However, the preventive effect of induced MT synthesis in pancreas was significantly observed in both diabetic models.
MT Prevents Diabetic Complications
Increasing evidence in both experimental and clinical studies suggests that free radicals are formed disproportionately in diabetes by glucose oxidation, nonenzymatic glycation of proteins, and the subsequent oxidative degradation of glycated proteins. These consequences of oxidative stress play a major role in the pathogenesis of diabetic complications (Cai and Kang, 2001a; Rosen et al., 2001). We recently have demonstrated that high levels of glucose induce cardiac cell death along with significant ROS formation (Cai et al., 2002a). In diabetic rats, abnormal spermatogenesis was found due to germ cell death (Cai et al., 2000b). Therefore, we hypothesized that diabetes-caused oxidative stress may play a critical role in inducing cell death in diabetes-targeted organs leading to severe organ dysfunction, and MT as a potent antioxidant may prevent diabetic complications through suppression of diabetic oxidative damage.
First we have preliminarily explored the preventive effect of MT in the heart on diabetes-induced cardiac toxicity (Cai and Kang, 2001b; Kang and Cai, 2001). We found that diabetes-caused cardiac toxicity, measured by serum creatine phosphokinase (CPK) and cardiac cell death, could be prevented in cardiac-specific MT-overexpressing transgenic mice (Cai and Kang, 2001b; Kang and Cai, 2001). Table 2 summarizes the features obtained from subsequent studies on the preventive role of MT in diabetes-induced cardiac toxicities. These studies suggested that MT in the hearts showed significant prevention against diabetes-induced serum cardiac enzyme activity, myocardial cell death, oxidative damage, and abnormal morphology and function.
MT Prevents Diabetic Toxicity in Cardiac Tissues


Note: The author previously produced an OVE26 diabetic mouse model (type 1 diabetes). They used this mouse model to crossbreed with a cardiac-specific MT-overexpressing transgenic mouse model. The OVE26MT mice are those in which cardiac MT is overexpressed about 20-fold and spontaneously develops diabetes (Liang et al., 2002).
A different transgenic mouse model in which MT is whole-body overexpressed was also used to investigate the protective effect of MT on diabetic toxicity. MT expression in the heart and kidney of transgenic (MT-TG) mice was significantly higher than that of their wild-type (WT) counterparts (Figure 2A, Cai et al., unpublished data). Our preliminary study (Cai et al., unpublished data) showed that diabetes was developed by administration of a single dose of STZ (150 mg/kg) in both MT-TG and WT mice. Impaired cardiac and renal function were observed in the WT (C57BL/6) diabetic mice, showing increases in cardiac enzyme activity (CPK) and BUN or creatinine levels, but these changes were not observed in the MT-TG diabetic mice (Figures 2B and 2C). These studies, therefore, strongly indicate that MT as a potent antioxidant significantly prevents the diabetes-induced cardiac and renal toxicities, respectively, and results in a prevention of these organ dysfunctions.

MT concentrations in the tissue of transgenic mice and its prevention from diabetic toxicity. Adult (8–10-week-old) male transgenic mice in which MT-I/II genes are overexpressed in whole-body and their wild-type (C57BL/6J) mice were used to measure cardiac and renal MT by Cd-hem methods (Cai et al., 2002b). Serum CPK as an index of hepatic function and serum BUN or creatinine (mg/dl) as an index of renal function were measured in WT and MT-TG diabetic mice 1 month after STZ-treatment. a: p < 0.05 or 0.01 vs control; b: p < 0.05 and 0.01 vs WT diabetic mice.
MT Induced by Diabetes Prevents Subsequent Pathogenic Injury
MT is also up-regulated in various organs in response to various types of stress, making it a so-called adaptive response (stress) protein (Klaassen and Liu, 1998; Cai et al., 1999). In our previous study, we demonstrated that MT expression was significantly enhanced in the liver and kidney of 1- and 6-month diabetic rats (Cai et al., 2002b) and in organs of 2-month diabetic mice (Figure 3). However, what biological importance diabetes-induced MT plays in diabetes and other coexisting pathogenesis has received limited investigative attention.
Enhanced expression of MT in tissues in response to diabetes. MT was immunostained in the heart, kidney, and liver of diabetic mice 2 months after STZ treatment.
We recently investigated the effect of diabetes on endotoxin-induced hepatic toxicity by measuring serum hepatic enzymes in normal and 2-month diabetic mice after endotoxin treatment. We found that 2-month diabetes animals exhibited a significant increase in hepatic MT expression compared to nondiabetic mice (Figure 3), and endotoxin further enhances the diabetes-induced MT expression (Song et al., unpublished data). More important, diabetic animals exhibited significantly reduced endotoxin-induced hepatic injury, showing significantly lower serum ALT activity and hepatic lipid peroxidation (TBARS) than one would expect from LPS-treated diabetic mice (Figure 4).

MT induced by diabetes prevents endotoxin-induced hepatic injury. Serum ALT and hepatic lipid peroxidation (TBARS) were measured in LPS-treated nondiabetic and diabetic mice 2 months after STZ treatment. a: p < 0.05 or 0.01 vs control; b: p < 0.05 or 0.01 vs the expected effect from diabetic and LPS-treated mice (diabetes + LPS − control = expected effect).
In support of this observation, an early study demonstrated that renal MT synthesis was increased in diabetic mice and was accompanied by a significant prevention of cisplatin-induced renal dysfunction (Jin et al., 1996). In addition, earlier studies have shown the preventive effects of diabetes on subsequent ischemia-induced cardiac injuries (Bouchard and Lamontagne, 1998; Tatsumi et al., 1998; Ravingerova et al., 2000; Schaffer et al., 2000). This may also be related to the induction of cardiac MT in response to diabetes as indicated in Figure 3. In summary, these results indicated that diabetes at certain stages of its pathogenesis can provide a resistance to subsequent insult-induced toxicity, and this enhanced resistance may be due to up-regulated MT expression.
Mechanisms by Which MT Prevents Diabetes, Diabetic Complications, and Subsequent Other Pathogenic Injury
MT as Antioxidant
Oxidative stress is the critical initiator for diabetic onset and complications, and antioxidants can prevent both diabetic onset and complications (Rosen et al., 2001). T-cell-mediated inflammatory autoimmune reaction and STZ- or alloxan-induced diabetes are all considered to be due to ROS and RNS formation leading to β-cell destruction. In addition, very low concentrations of antioxidants found in animal pancreatic islets and hearts make these organs vulnerable to STZ or alloxan-induced diabetes through the generation of ROS and RNS (Lenzen et al., 1996; Tiedge et al., 1997).
Pancreas contains relatively high levels of MT compared to other tissues such as liver and heart (Onosaka et al., 1988; Andrews et al., 1990; Tomita and Matsubara, 2000). The high expression of MT in pancreas may suggest that it is required for normal physiological function. Indeed, mice with the MT-I and -II gene null show obesity and hyperlipidemia (Beattie et al., 1998). Furthermore, although isolated islets from MT-null and WT mice had similar insulin content (168 ng insulin/five islets for WT mice vs 170 ± 16 ng/five islets for MT-null mice), the islets from the MT-null mice showed much lower levels of basal and maximal insulin releisletsase (Laychock et al., 2000). These results suggest the requirement of MT for physiological function of pancreatic β-cells in which MT may act as an antioxidant in scavenging ROS and RNS to preserve cell membrane integrity, protein sulfhydryls or nucleic acids, and secretary responsiveness. Overexpression of MT induced by Zn in pancreatic β-cells has been shown to provide significant protection against STZ-induced diabetes in rats and mice (Table 1), suggesting that MT plays an important role in the prevention of oxidative injury which otherwise initiates diabetes. The possibility that the prevention of the diabetes was due to other effects of the inducer (such as Zn) was eliminated by use of genetically manipulated pancreatic-specific MT-overexpressing transgenic mice (Chen et al., 2001). In these mice, both spontaneously developed and STZ-induced diabetes were prevented. In cultured islets from control mice and MT-TG mice, MT decreases STZ-induced islet disruption, DNA breakage, and depletion of NAD+. These results demonstrate that MT can reduce diabetes through inhibition of STZ-induced oxidative DNA damage (Chen et al., 2001). Using cardiac MT-TG mice, significant prevention of diabetic cardiomyopathy was also found along with significant suppression of diabetes-induced ROS and RNS accumulation and oxidative damage (Table 2).
Activation of NF-κB (a ROS-sensitive transcription factor) plays a critical role in initiating β-cell death leading to diabetic development (Darville et al., 2001; Heimberg et al., 2001; Ho et al., 2001). MT plays a role in the negative regulation of NF-κB (Sakurai et al., 1999; Papouli et al., 2002), and dietary Zn supplementation with overexpressing pancreatic MT was found to significantly attenuate hyperglycemia in prodiabetes db/db mice and chemically induced diabetes by alloxan or STZ, accompanied by significant inhibition of NF-κB activation (Ho et al., 2001; Simon and Taylor, 2001).
In further support of the antioxidant action of MT, there is evidence indicating that up-regulation of SOD made mice resistant to diabetes development (Kubisch et al., 1997). Thioredoxin (TRX), a redox (reduction/ oxidation)-active protein, has recently been shown to protect cells from oxidative stress. Transgenic mice with specific expression of TRX in pancreatic islets showed a significantly lower incidence of spontaneously developed and STZ-induced diabetes as compared to their WT counterparts (Hotta et al., 1998). In addition, low-dose radiation has been known to up-regulate antioxidant capacity leading to a wide range of adaptive response (Cai, 1999). It has also been reported that low-dose radiation prevents spontaneous or chemically induced development of diabetes through enhanced pancreatic antioxidant capacity (Takehara et al., 1995; Takahashi et al., 2000).
MT as Regulator of Zn Homeostasis
MT is a regulator of Zn homeostasis. When the target tissues are exposed to oxidative stress, Zn-binding MT releases Zn that in turn plays an important role in the prevention of oxidative damage (Maret, 2000). High Zn content was found in islets. For example, Zn content of Sprague Dawley rat pancreatic islets was found to be 27.9 ± 2.0 ng/mg islet cells (Kim et al., 2000). This may suggest the requirement of Zn for insulin function (Kim et al., 2000; Lukowiak et al., 2001; Tang and Shay, 2001). Several physiological roles of Zn in insulin function have been recently indicated (Salgueiro et al., 2001): (a) Zn is required for insulin-forming hexameric crystals which are stored in β-cells and released into the portal venous system at the time of β-cell degranulation. Furthermore, pancreatic Zn level in db/db mice was significantly lower than in lean mice, and Zn supplementation restored both pancreatic Zn concentration and serum glucose of db/db mice (Simon and Taylor, 2001). (b) The Zn-to-insulin ratio within these crystals determines the ternary structure, antigenic properties of insulin, and the binding capacity to its receptor. (c) Zn pretreatment not only accelerates and increases the magnitude of insulin binding to the receptor, but also simultaneously inhibits the degradation of insulin by the liver plasma membranes. (d) Zn is closely involved in insulin-dependent metabolism of protein, carbohydrate, and lipids, and chelation of Zn is able to induce diabetes (Epand et al., 1985). (e) Zn is an important factor in preventing cell death. Zn can inhibit caspase-3 activation to prevent cell death that is initiated via Fas/FasL pathway or mitochondrial cytochrome c pathway (Yamada et al., 1999). β-cell death under exposure to different insults plays a critical role in the development of diabetes (Chandra et al., 2001). Therefore, Zn plays important and wide-ranging functions in regulation of insulin function and prevention of spontaneous and chemical-induced diabetes.
However, whether these functions of Zn are MT-dependent is unclear because MT is both normally found in high concentration in pancreas and also highly inducible by Zn. MT-null mice were used to investigate the effect of Zn supplement against STZ-induced diabetes in order to dissect the protective role against STZ-induced diabetes either by MT or Zn (Apostolova et al., 1997), but the results are still inconclusive. In that experiment, Zn pretreatment was given at two dose levels: 1 mg/kg and 10 mg/kg body weight as ZnSO4. The high dose of Zn fully suppressed development of hyperglycemia in both MT-null and WT mice after STZ treatment. However, the low-dose Zn pretreatment had a definite inhibitory effect on STZ-induced diabetes development in MT-null mice, but no marked effect in WT control. This study seems to suggest that Zn itself can play an important role in the prevention of diabetic pathogenesis under certain conditions (Apostolova et al., 1997). However, glucose-stimulated insulin secretion in MT-null mice is less than in WT mice, suggesting the requirement of pancreatic MT for the insulin secretion under normal condition (Laychock et al., 2000).
The different effects between low-dose and high-dose levels of Zn may be due to MT induction in nonpancreatic tissues. In C57BL/6J mice, low-dose Zn supplement induced hepatic and renal MT, which binds more Zn leading to less free Zn available to pancreas β-cells, whereas in MT-null mice the lack of MT induction, or subsequent Zn binding, in nonpancreatic tissues causes sufficient free Zn to be available for pancreatic β-cells. It has been documented that MT induction by low-dose Zn supplementation is lower in the pancreas than in the liver, whereas high concentration of Zn supplementation could induce a higher proportion of pancreatic MT synthesis compared to liver (Zimny et al., 1993). This study suggests that liver is the target organ for initial storage of Zn although pancreas is highly responsible for Zn-induced MT synthesis. This explanation may also explain the failure of MT to demonstrate protection in the alloxan-induced diabetes study by Minami et al. (1999).
The aforementioned studies strongly demonstrate that MT prevents diabetes through its antioxidant action, Zn regulation, or both. Enhanced expression of MT by inducers other than Zn provides significant prevention of diabetes. For example, cytokines (Laychock et al., 2000) and Cu (Vinci et al., 1995; Tobia et al., 1998; Sitasawad et al., 2001) significantly prevented diabetes in rats and mice. These results support the antioxidant action of MT in the prevention of diabetes. In addition, although Zn is required for insulin physiological function and provides preventive effects from spontaneous and chemical-induced diabetes, high levels of free Zn also cause pancreatic β-cell damage resulting in diabetes (Kim et al., 2000). The fact that the free-Zn-induced pancreatic β-cell damage can be prevented by Zn chelator (Kim et al., 2000) suggests that MT plays an important role in maintaining Zn homeostasis by binding free Zn and releasing Zn when it is required. Therefore, MT plays the critical role in coordination with Zn for preventing diabetes development, as outlined in Figure 5.
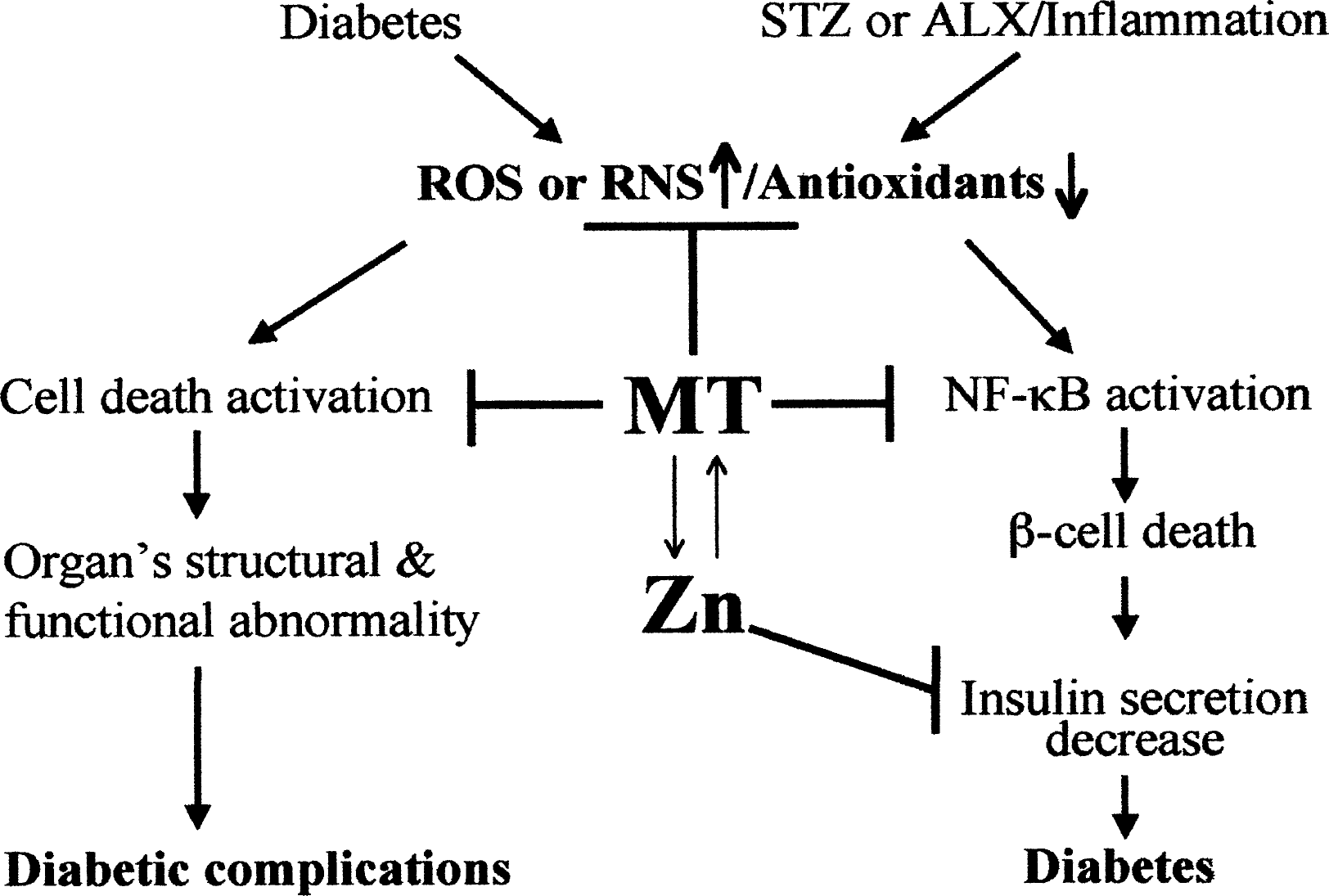
Outline of mechanisms by which MT coordinately with Zn prevents diabetes development and diabetic complications.
Regarding the relationship between MT and Zn in nonpancreatic tissues and their function in preventing diabetic complications, to date no investigation has been documented. With the exception of Zn's role in insulin regulation and secretion, it would be expected that the relationship interaction between Zn and MT would be the same as in the pancreas (Figure 5).
In summary, MT as a potent antioxidant, adaptive protein, and Zn homeostasis regulator plays important roles in the prevention of diabetes development, complications, and subsequent suffered pathogenic toxicity. Development of strategies for up-regulation of MT would be of interest for pharmaceutical and medical applications.