
Abstract
Vasoactive intestinal peptide (VIP) is a vasodilator peptide present in cerebrovascular nerves. Vasoactive intestinal peptide can activate VPAC1, VPAC2 and the NPR-C receptor. This study sought to determine the receptors involved in VIP-induced vasodilation of porcine basilar arteries. Porcine basilar arteries contained the messenger ribonucleic acid of all three receptors. Immunocytochemical analysis of porcine basilar arteries revealed that the VPAC1 receptor is expressed on the endothelium, VPAC2 on the outer layers of the media and the NPR-C receptor throughout the artery, including nerves. Vasodilator responses to all receptor agonists showed that the receptors are functional. The vasodilator response to the VPAC1 receptor agonist was inhibited by
Introduction
Vasoactive intestinal peptide (VIP) is an important neurotransmitter in cerebral arteries, and has been located in cerebral perivascular nerves of rats, rabbits, pigs and humans (Nozaki et al, 1993; Dalsgaard et al, 2003; Larsson et al, 1976; Gulbenkian et al, 1995). Vasoactive intestinal peptide is a highly potent cerebral vasodilator both in vitro (Duckles and Said, 1982; Gaw et al, 1991; Matthew et al, 1997; Toda et al, 1997; Seebeck et al, 2002) and in vivo (Kitazono et al, 1993). Moreover, infusion of VIP into the carotid artery of baboons causes a marked increase in cerebral blood flow accompanied with electroencephalogram changes, indicative of the activation of cerebral neurones (McCulloch and Edvinsson, 1980). Vasoactive intestinal peptide also increases cerebral blood flow after intracerebroventricular administration in rabbits, goats, dogs and rats (Heistad et al, 1980; Larsen et al, 1981; Wilson et al, 1981; Tuor et al, 1990).
The cerebral vasodilator action of VIP involves the participation of nitric oxide (NO). Thus, inhibition of NO synthesis results in the attenuation of VIP-induced vasodilation of endothelium-denuded sheep middle cerebral arteries (Gaw et al, 1991) and bovine cerebral arteries (Gonzalez et al, 1997). In addition, the relaxant effect of VIP on isolated rat basilar arteries is inhibited by ω-conotoxin or 7-nitroindazole (7-NI), showing that nitrergic neurons are activated by VIP (Seebeck et al, 2002). Moreover, transmural nerve stimulation (TNS) of isolated cerebral arteries results in relaxation, which is susceptible to NO synthesis inhibition (Toda and Okamura, 1990a, b ; Gaw et al, 1991; Ayajiki et al, 1993; Matthew and Wadsworth, 1997; Matthew et al, 1997; Tanaka et al, 1999). Stimulation of the facial nerve of the cat results in the release of VIP into the superfusate of the cerebral cortex, showing that VIP is also released after vasodilator nerve stimulation (Goadsby and Shelley, 1990). Vasoactive intestinal peptide antiserum has been shown to inhibit TNS-induced relaxation of sheep middle cerebral arteries (Matthew et al, 1997) and different cat cerebral arteries (Brayden and Bevan, 1986). In addition, the nonspecific VIP receptor antagonist, VIP(7–28), significantly reduced the TNS-induced relaxation in the rabbit basilar artery (Olivar et al, 2000).
Three different receptor subtypes are known with which VIP is capable of interacting. Within the group II secretin receptor G protein-coupled receptor family, the VPAC1 and VPAC2 receptors have been identified, cloned (Harmar et al, 1998) and are considered to be the main physiological receptors that mediate the effects of VIP. Members of this family couple to the production of cAMP, and are capable of activating additional signalling pathways (McCulloch et al, 2002). Both VPAC1 and VPAC2 receptors have been shown to be expressed in equal amounts in human cerebral arteries regardless of endothelium; however, their location remains undetermined (Knutsson and Edvinsson, 2002). This finding concurs with previous experiments which have shown that VIP-induced vasodilation of cerebral arteries is independent of endothelium (Duckles and Said, 1982; Lee et al, 1984; Edvinsson and McCulloch, 1985; Nozaki et al, 1990; Gaw et al, 1991; Stojic et al, 2003). A third receptor has been identified to mediate VIP-induced relaxation of gastric smooth muscle: this is the natriuretic peptide clearance receptor (NPR-C) receptor (Akiho et al, 1995; Murthy et al, 2000). NPR-C receptor activation inhibits adenylate cyclase via coupling to Gi-1 and Gi-2 proteins (Murthy et al, 2000). The VIP-mediated stimulation of NPR-C receptors in rabbit gastric smooth muscle cells activates endothelial nitric oxide synthase (eNOS) via a Gi-1- and Gi-2-mediated Ca2+ influx (Murthy et al, 1998). However, it is not known whether the NPR-C receptor is present or has any vasoactive function in the cerebral circulation.
The objectives of the present study were to determine the VPAC1, VPAC2 and NPR-C receptor messenger ribonucleic acid (mRNA) expression in porcine basilar arteries and their localisation. Functional data, including the role of these receptors in neurogenic relaxation, was obtained with the use of recently developed ligands for these receptors and the role of NO was also investigated.
Materials and methods
Basilar arteries were isolated from pig heads obtained from an abattoir within 90 mins of slaughter and placed in Krebs-Henseleit solution (composition, mmol/L: 118.3 NaCl, NaHCO3, 11.1 glucose, 4.7 KCl, 1.2 MgSO4, 1.2 KH2PO4 and 2.5 CaCl2), gassed with 95% O2 and 5% CO2. Using light microscopy, connective and brain tissue was removed from the basilar artery and any remaining intraluminal blood was gently flushed out with Krebs-Henseleit solution.
Ribonucleic Acid (RNA) Isolation and Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
Five basilar arteries were immediately snap-frozen at −80°C and ground up in liquid nitrogen with a prefrozen (–80°C) mortar and pestle. For control reactions, RNA was extracted from tissue obtained from four separate brain regions of the pig: cortex, thalamus, hypothalamus and cerebellum. Total RNA extraction was performed using the RNAzol reagent (Biogenesis Ltd, UK). First-strand cDNA was synthesised from 10 μg of total RNA using the Superscript II RT First Strand Synthesis System (Invitrogen Life Technologies, UK) and random hexamers.
Oligonucleotide primers were designed to amplify the VPAC2 and NPR-C receptor sequences in pig based on the conserved regions identified through comparison of the published sequences of the mouse, rat and human genes, respectively. For the VPAC2 receptor, the primers used were VPAC2fp: 5'-TGCCTCTTCAGGAAGCTGCACTG-3' and VPAC2rp: 5'-GCAAACACCATGTAGTGGACGCC-3', and for the NPR-C receptor the primers used were NPRCfp: 5'-GG AGCGGAACTGCTACTTCAC-3' and NPRCrp: 5'-GTTTC; TCAACTGAACTTTTCACCTCC-3'. Specific primers for the pig VPAC1 receptor were derived from the published cDNA sequence accession number U49434, pVPAC1fp: 5'-GCACA CCCTACTTGCCGTG-3' and pVPAC1rp: 5'-CTCCTGACGGG TGCTGGTATTTG-3'. Specific primers for the pig PAC1 receptor were derived from the published gene sequence accession number AC091758, pPAC1.fp 5'-TTTGATGCCTGT GGGTTTGATGAG-3' and pPAC1.rp [5'-GAGAACAGCCACC ACGAAGCC-3'. PCR amplification for each receptor was performed with first-strand cDNA with 15 pmol of the respective forward and reverse primers using Taq Polymerase (Promega, UK) according to the manufacturer's instructions. The reactions were first heated to 95°C for 2 min, followed by 35 cycles of 94°C for 30 secs, 60°C for 30 secs and 72°C for 90 secs. Amplified DNA products were analysed by agarose gel electrophoresis, with positive and negative controls. Amplified products were subcloned and verified by sequence analysis.
Immunocytochemistry
The basilar artery was immediately fixed in neutral buffered saline (formalin) and dehydrated through graded alcohol to xylene. The artery section was then orientated for cross-section or longtitudinal sectioning, wax embedded, and 4-μm sections were cut using a rotary microtome (Leica RM2125 RTF, Leica Instruments GmbH, Germany). The sections were then mounted onto presilanated slides and rehydrated through graded alcohol to water. The sections were then placed in 0.3% H2O2 for 10 min to inhibit endogenous peroxidases and washed in Tris buffered saline (TBS) for 5 min. Antigen retrieval pretreatment was performed by microwave pressure-cooking for 5 min (0.37 g ethylenediamine tetraacetic acid (EDTA) and 0.55 g Tris were added to 1 L of distilled water). The sections were then washed in TBS for 5 min and 20% normal goat serum (in TBS) was added for 20 min to block nonspecific binding of the secondary antibody, a biotinylated goat anti-rabbit/anti-mouse antibody. The normal goat serum was then drained before addition of the primary antibody. Primary antibodies used were rabbit polyclonal, anti-human VPAC1 and VPAC2 antibodies (both 1:100, generous gifts from S Schulz, Otto-von-Guericke University Magdeburg, Germany – see Schulz et al, 2004), rabbit polyclonal, antihuman NPR-C antibody (1:1000, kind gift from K Omori, Tanabe, Seiyaku Co. Ltd, Osaka, Japan – see Fujishige et al, 1998), rabbit polyclonal, antiporcine VIP antibody (1:25, Europath, UK), mouse monoclonal, anti-human eNOS antibody (1:400, BD Biosciences, UK), mouse monoclonal, antihuman neuronal nitric oxide synthase (nNOS) antibody (1:1000, BD Biosciences, UK) and mouse monoclonal, antihuman neurofilament (1:400, Dako, UK). All of the primary antibodies used were applied for 60 min, then drained off, and the sections washed in TBS for 5 min. Each staining run contained a negative control slide, processed in the absence of a primary antibody. The biotinylated goat antirabbit/antimouse secondary antibody (LSAB2 kit, Dako, UK) was then added for 30 min; then the sections were washed in TBS for 5 min. The streptavidin—horseradish peroxidase complex tertiary antibody (LSAB2 kit, Dako, UK) was then applied for 40 min, then washed off in TBS for 5 min. The sections were then placed in 0.5% diaminobenzidine tetrahydrochloride (DAB) (activated with 700 μL of 30% H2O2) for 10 min, resulting in an insoluble brown precipitate formation at the site of the bound primary antibody. The sections were then rinsed in water, then counterstained in haematoxylin for 3 min, and then rinsed in water. Sections were then immersed in 0.5% acid alcohol (10 dips), washed in water, then placed in Scott's substitute tap water (SSTW) for 30 secs. After washing in water, the sections were then placed in 0.5% copper sulphate to intensify the stain. After completion of the staining, sections were then dehydrated by sequential immersion in water, through graded alcohols to xylene and then mounted in DPX (xylene-based mountant) with a glass coverslip. The sections were then viewed with a light microscope fitted with a daylight filter. Each staining run contained a negative control where the primary antibody was omitted from the protocol. All staining results were verified by duplicating the results in different arteries and in control tissues (gut, lung and brain, not shown, n = 3–5). Images were magnified with a light microscope (Nikon Eclipse E600) and captured with a digital camera (Fujix HC 300Z).
Myography Studies of Vascular Relaxation
Rings of porcine basilar arteries (2-mm long) were mounted on two 40-μm diameter wires in a 4-channel myograph (MultiMyograph 610, Danish Myo Technology, Aarhus, Denmark) in Krebs-Henseleit solution, gassed with 95% O2 and 5% CO2 at 36°C to reduce spontaneous activity of the arteries (Osol and Halpern, 1988). The responses were recorded via computer software (MyoDaq V.2.01, Danish Myo Technology, Aarhus, Denmark). A passive tension of 1.5 g was applied gradually over 60 min. The vessels were then allowed to equilibrate for 30 min before the addition of 40 mmol/L KCl to confirm contractile viability of each vessel. The chambers were then washed 3 times to ensure that the KCl was removed and the vessels were allowed to return to the passive tension of 1.5 g. The tension was readjusted if necessary. U46619 (0.1 μmol/L; was used to obtain a reproducible, stable contraction of the arteries before cumulative generation of relaxant responses to either VIP, the VPAC1 agonist ([K15,R16,L27]VIP(1–7)/GRF(8–27)), the VPAC2 agonist (Ro-25–1553) or the NPR-C agonist (cANP(4–23)). Any subsequent concentration response curves were generated after a 60-min equilibration period and only 2 concentration—response curves were generated per artery ring to avoid tachyphylaxis. Selective VPAC1 ([AcHis1,DPhe2,K15,R16,L27 VIP/GRF]), VPAC2 (PG-99–465) and NPR-C (ANP(1–11)) antagonists were also tested for their effect on the relaxation induced by the different agonists. Endothelium was removed in some experiments by gently rubbing the luminal surface with a roughened wire three times. Successful removal of the endothelium was confirmed by a subsequent lack of response to carbachol.
Transmural Nerve Stimulation
Pig basilar artery rings (2-mm long) were mounted in a 4-channel myograph as above using nonconducting nylon myograph jaws incorporating platinum electrodes. Guanethidine (5 μmol/L) and 1 μmol/L atropine were present throughout the experiment to eliminate the influence of adrenergic and cholinergic nerves on nitrergic nerves. During the contraction to U46619, the vessels were stimulated with square wave pulses delivered by an electrical stimulator (Grass S44, Quincy, MA, USA) via the platinum electrodes. Optimum stimulating voltage (10–150 V) for each vessel was then obtained and test responses were obtained in U46619-contracted vessels, causing a rapid, transient and submaximal (∼50%) relaxation of the artery smooth muscle. The frequencies studied were 8 and 20 Hz. These were applied for 10 secs, delivering square wave pulses, 0.2 ms in duration. The vessels were then washed free of U46619, allowed to equilibrate for 30 min, recontracted with U46619 and tested a second time to obtain time controls. Parallel experiments repeated the stimulation with selected pharmacological interventions. The relaxant responses were expressed as a percentage of the maximum relaxation, which was obtained with 1 μmol/L forskolin.
Drugs and Solutions
Formalin composition was 100 mL of 40% formaldehyde in 900 mL phosphate buffer (4 g NaH2PO4 · 2H2O and 6.5 g NaHPO4/L). Tris buffer: 0.038 mol/L (6.06 g/L) Trizma HCl and 0.011 mol/L (1.39 g/l) Trizma base dissolved in distilled water. Tris buffered saline: 1 part Tris buffer, 9 parts saline (0.9% (w/v) NaCl dissolved in distilled water). DAB: 1 vial of frozen DAB (30 mL aliquot, 0.5% w/v) dissolved in 300 mL Tris buffer. Scott's substitute tap water (SSTW): 0.08 mol/L (20 g/L) MgSO4 · 7H2O and 0.08 mol/L (3.5 g/L) NaHCO3 in distilled water. Copper sulphate solution (0.5%): 0.008 mol/L (5 g/L) copper sulphate in saline.
Trizma base, Trizma HCl, DAB, NaCl, NaHCO3, MgSO4, silane (3-aminopropyltriethoxy silane), U46619 (9,11-dideoxy-11α, 9α-epoxymethanoprostaglandin F2α), carbachol, guanethidine and atropine were purchased form Sigma, UK. Copper sulphate, KCl, glucose, KH2PO4, CaCl2 and DPX mounting media were purchased from VWR International, Poole, UK. H2O2 was bought from AAH Pharmaceuticals Ltd, UK. Paraffin wax was bought from TCS Microbiology, Boltof Claydon, UK. Xylene and industrial alcohol were purchased from Genta Medical, York, UK. Microscope slides were purchased from Menzel-Glazer, Germany. [K15,R16,L27]VIP(1–7)/GRF(8–27), Ro-25–1553, AcHis1,DPhe2,K15,R16,L27 (VIP/GRF) and PG-99–465 were kind gifts from Professor Robberecht (Department of Biochemistry and Nutrition, University of Brussels, Belgium). ANP(1–11) and cANP(4–23) were purchased from American Peptide Co. (Sunnyvale, CA, USA).
Statistical Analysis
Statistical analysis of concentration—response curves was performed by ANOVA with repeated measures (cross-over model), which determines a significant difference between two curves overall. A paired Student's t-test was used to compare TNS responses. n = number of arteries from different animals. Any statistical test yielding a P-value > 0.05 was considered significant.
Results
Since the vasodilator action of VIP on blood vessels could possibly be mediated by three different receptors, we first investigated which were expressed in porcine cerebral arteries by RT-PCR analysis. Polymerase chain reaction products of the expected sizes for the VPAC1 (550 base pair (bp)), VPAC2 (580 bp) and NPR-C (410 bp) receptors were amplified from porcine basilar arteries (shown in Figure 1). For the VPAC2 receptor, the 580 bp fragment was detected also in the thalamus and cerebellum and weakly in the cortex and hypothalamus, whereas the 550 bp fragment corresponding to the VPAC1 receptor was only very weakly detected in the cerebellum and hypothalamus. In addition, a smaller 300 bp product was amplified from all brain tissues with the pVPAC1 primers. The 410 bp fragment corresponding to the NPR-C receptor is expressed also in a rat SCN2.2 cell line used as a positive control in this study (Earnest et al, 1999). We next determined that the amplified DNA fragments corresponded to the correct receptor gene sequence by sequence analysis (data not shown). The small 300 bp amplification product amplified by the pVPAC1 receptor primers was also sequenced and determined to be nonspecific.
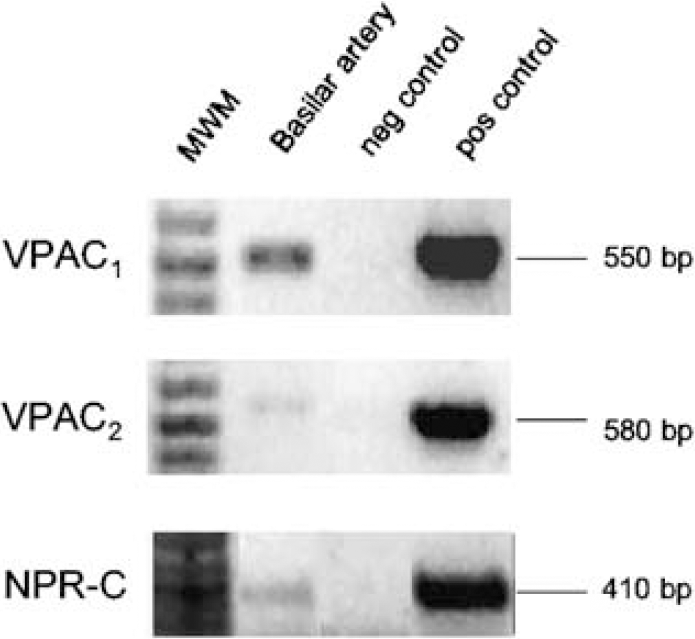
Reverse transcriptase-polymerase chain reaction results from porcine basilar arteries. The amplified products (35 PCR cycles) correspond to the mRNA encoding porcine VPAC1, VPAC2 and NPR-C receptors. The top panel shows the 550 bp product of the VPAC1 receptor from porcine basilar arteries. The middle panel shows the 580 bp product of the VPAC2 receptor found in the pig basilar artery. The bottom panel shows the 410 bp product of the NPR-C receptor found in the pig basilar artery. MWM = molecular weight marker, negative control = negative control DNA, positive control = pig VPAC1 cDNA, human VPAC2 cDNA and rat SCN for VPAC1, VPAC2 and NPR-C receptors, respectively.
The distribution of the receptor proteins was determined by immunocytochemical analysis. Porcine basilar arteries were found to be densely innervated by nerve bundles in the adventitia parallel to the lumen in addition to a network of fine nerve fibres (Figure 2A). Vasoactive intestinal peptide immunoreactivity was detected in the media/adventitial border, suggesting that this peptide is released from intramural nerves in the vicinity of the smooth muscle cells (Figure 2B). Endothelial cells stained positively for the VPAC1 receptor, but no immunoreactivity was observed in smooth muscle cells (Figure 2C). Immunoreactivity for the VPAC2 receptor is most prominent in the outer layers of the media, with immunoreactivity also detected in the endothelium (Figure 2D). The NPR-C receptor appears to have widespread cellular distribution, with immunoreactivity detected in the endothelium, smooth muscle, adventitia and nerve bundles (Figure 2E). The PACAP-specific receptor, PAC1, was not detected in porcine basilar arteries by RT-PCR or immunocytochemistry (data not shown). The constitutive NOS isozymes, eNOS and nNOS, were also detected on the endothelium and nerves, respectively (data not shown). Negative controls were obtained by omitting the primary antibody from the protocol and did not result in any staining (not shown).
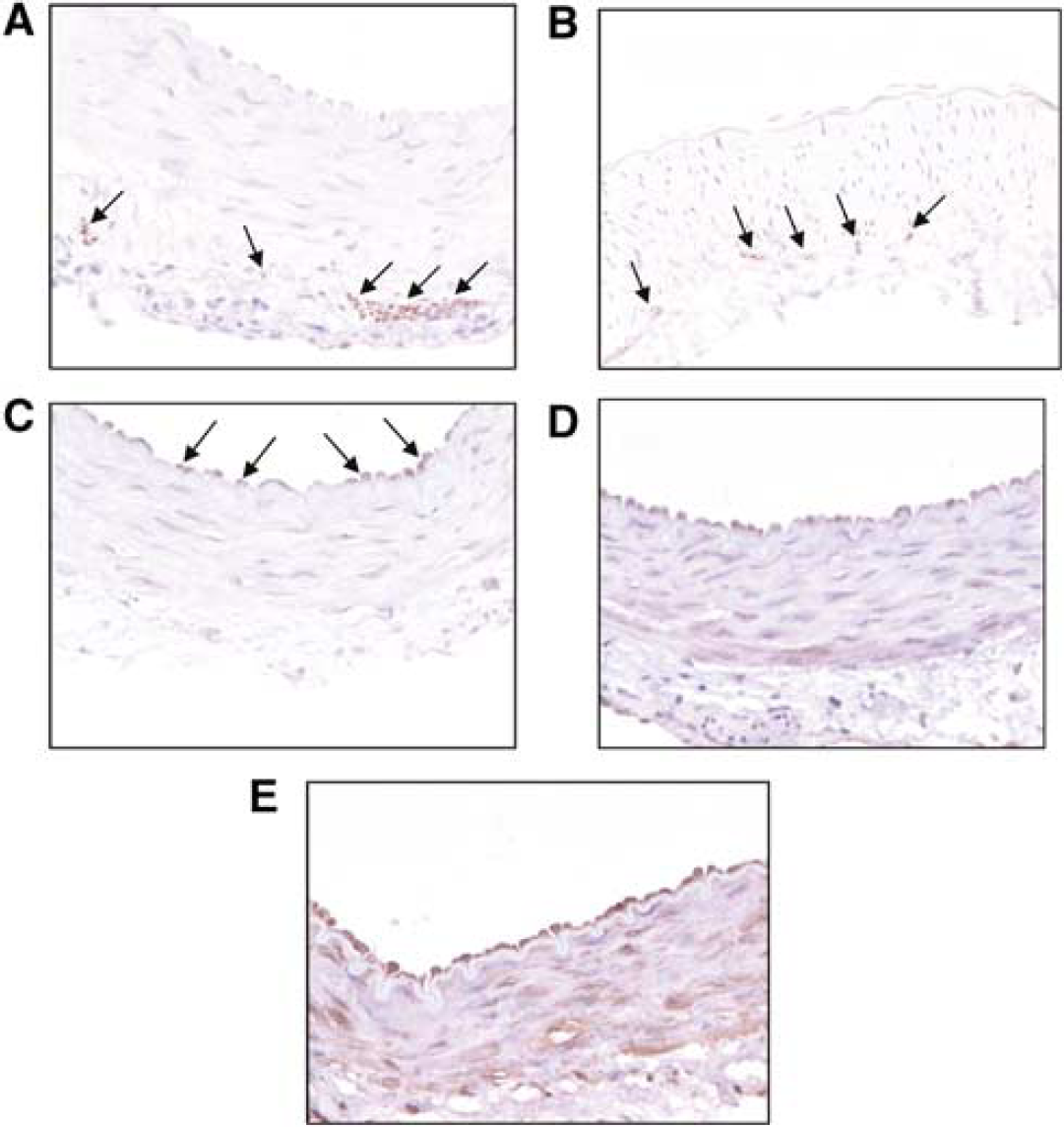
(
The functional activity of VIP and VIP receptor-specific agonists and antagonists was then investigated on intact basilar artery rings. Vasoactive intestinal peptide added cumulatively at a range of 10−9–3 × 10−7 mol/L produced concentration-dependent relaxation of the U46619-induced tone (maximum relaxation ∼100%). All generated concentration-dependent relaxations of U46619-contracted pig basilar arteries were reproducible. The NOS inhibitor NG-nitro-
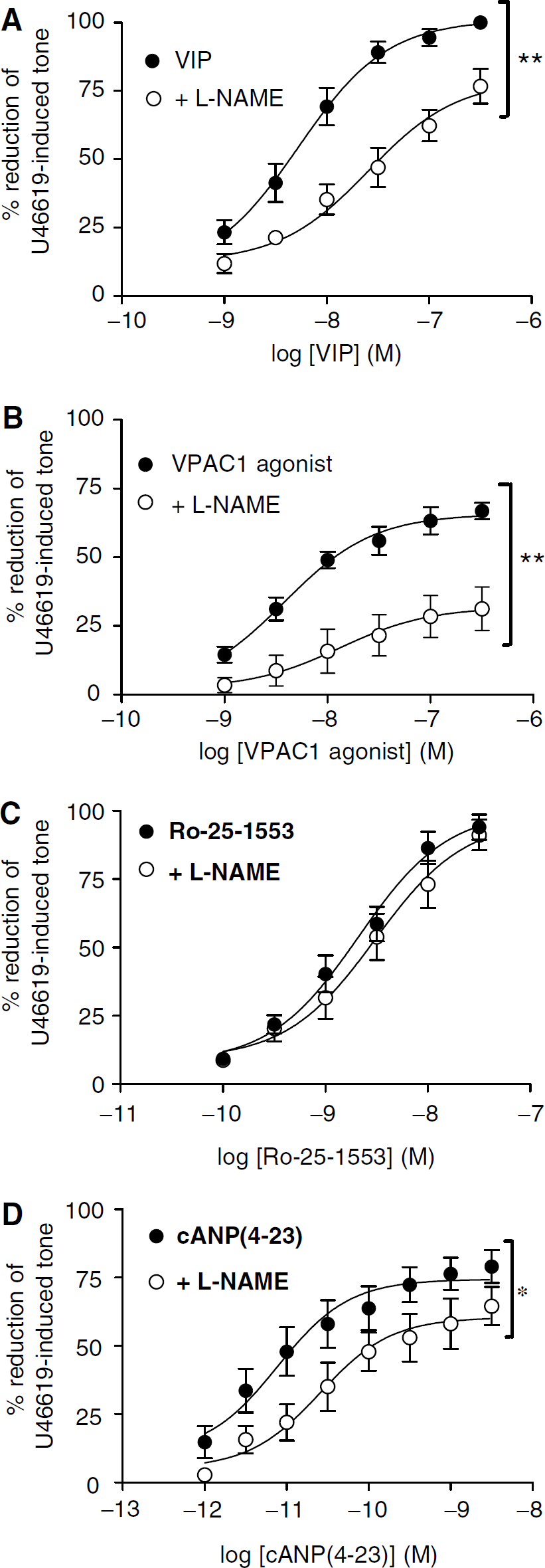
Porcine basilar arteries precontracted with 0.1 μmol/L U46619. (
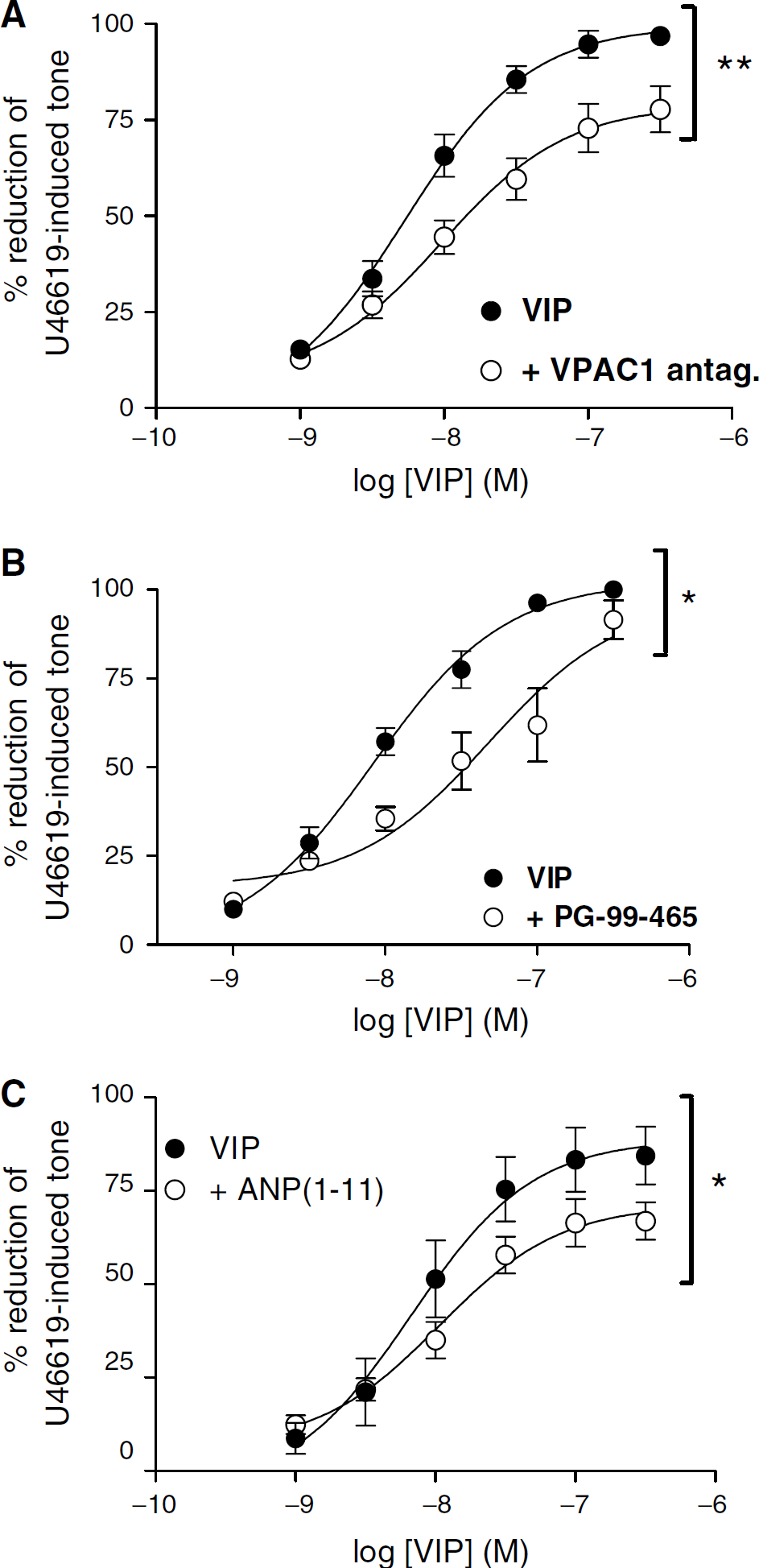
Porcine basilar arteries precontracted with 0.1 μmol/L U46619. Specific inhibition of VIP-induced relaxation of porcine basilar arteries by (
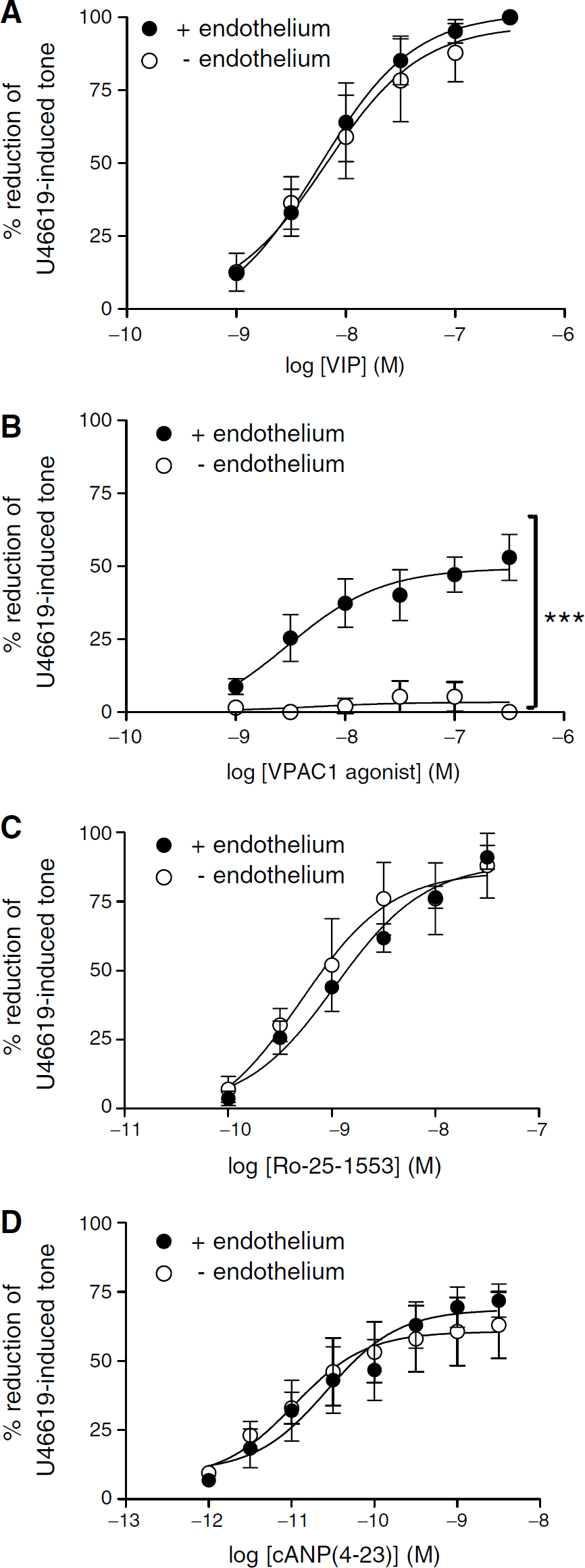
Precontracted porcine basilar arteries (0.1 μmol/L U46619). (
Transmural nerve stimulation responses at 20 Hz were reproducible when repeated after a 30-min interval (Figure 6A) and abolished by tetrodotoxin (TTX) or
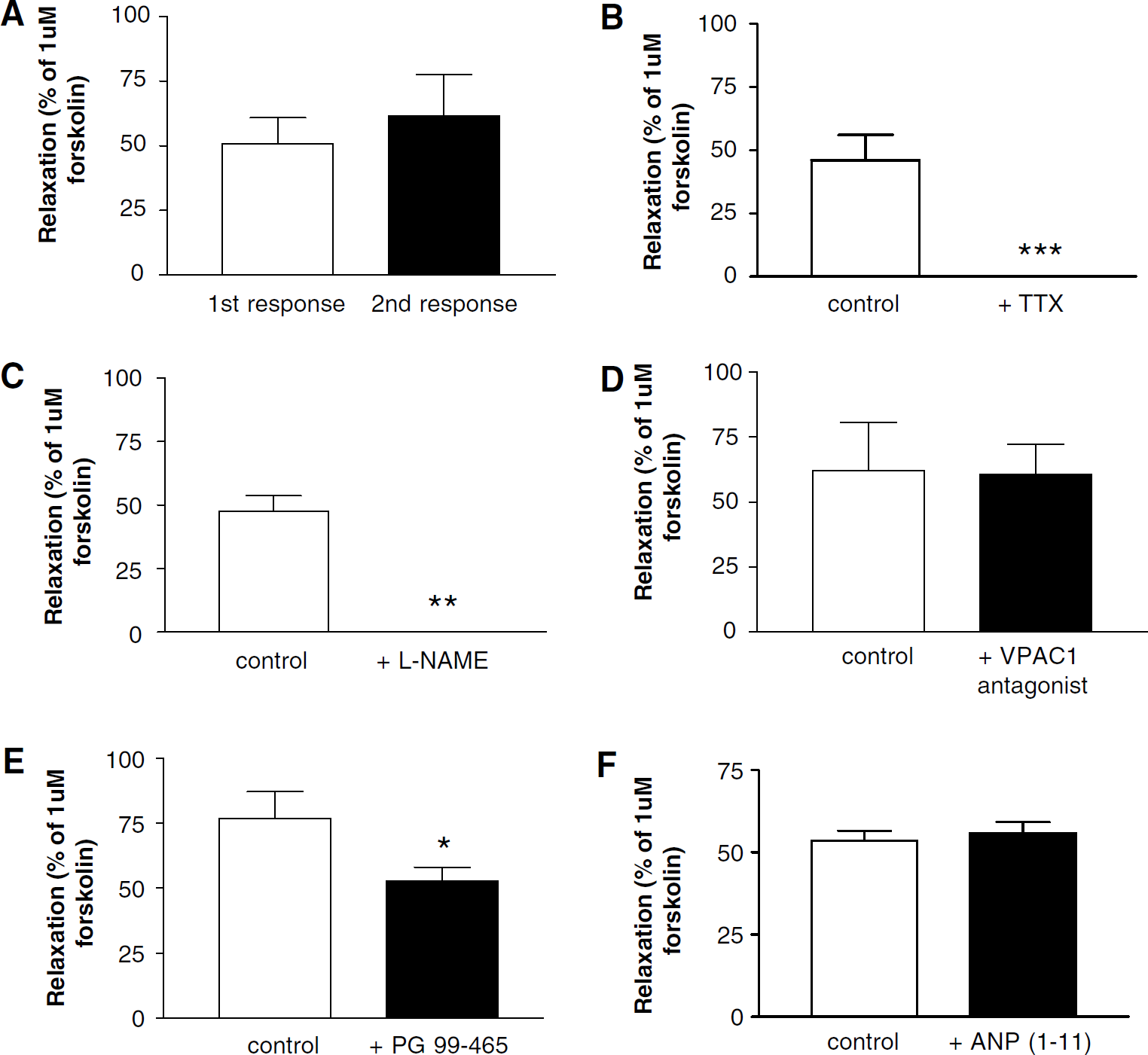
Porcine basilar arteries precontracted with 0.1 μmol/L U46619. (
Discussion
Vasoactive intestinal peptide produced a concentration-dependent relaxation of U46619-contracted porcine basilar arteries and the response was attenuated with
In the present study, the VPAC1 receptor was identified in the endothelium. The VPAC1 agonist induces NO- and endothelial-dependent vasodilation over the same concentration range as VIP, which is in agreement with the immunocytochemical findings that this receptor is located on the endothelium. This is in contrast to a previous study, which identified VPAC1 receptors on vascular smooth muscle cells of rat superficial cerebral arteries (Fahrenkrug et al, 2000). This highlights a possible species difference or a difference between the distributions of the VPAC1 receptor in basilar and superficial cerebral arteries (i.e. large versus small diameter arteries).
The VPAC2 receptor was most prominent in the outer layers of the media. Ro-25–1553, the VPAC2 agonist, is roughly 10 × more potent than VIP. This is due to the increased stability consequent on resistance to peptidases (Bolin et al, 1997; Xia et al, 1997). The response to VPAC2 stimulation is both NO- and endothelium-independent and probably induces relaxation via the cAMP pathway in smooth muscle cells. This finding is in agreement with the immunocytochemical detection of VPAC2 receptors in the outer layers of the smooth muscle. PG-99–465 significantly attenuated the neurogenic relaxation obtained by TNS at 20 Hz. This suggests that VIP released from intramural nerves (identified at the media-adventitial border) acts on VPAC2 receptors in the outer layers of the media to contribute to the relaxation. As NOS inhibition abolishes this response, NO generation is also implicated. The model of interaction between VIP and NO is not clear; however, it is possible that NO induces VIP release during high-frequency stimulation. This is a possible model of interaction as NOS inhibition completely inhibits the TNS response, but in the absence of NOS inhibition VPAC2 receptor antagonism attenuates the relaxation, suggesting that VIP might also be released by NO. Nitric oxide-induced VIP release has also been proposed in models of gastrointestinal smooth muscle relaxation. This was initially proposed after the observation that electrically induced VIP release from guinea-pig stomach smooth muscle strips was reduced by
The NPR-C agonist cANP(4–23) was vasoactive, showing that this receptor is functional. This response was obtained independently of endothelium, and was significantly inhibited by
The relaxant response to VIP is preserved in denuded porcine basilar arteries, suggesting that the contribution of the VPAC1 receptor (and the endothelium) is not essential for the vasodilator effect VIP. This suggests that the other receptors can compensate for the lack of the VPAC1 receptor. This would explain why there are multiple receptor types present (an in-built redundancy of some receptors due to the importance of VIP-induced vasodilation). This is in agreement with previous studies which have shown endothelium-independent VIP-induced vasodilation of cerebral arteries (Duckles and Said, 1982; Lee et al, 1984; Gaw et al, 1991; Stojic et al, 2003). Additional evidence that VIP acts through each of these receptors in porcine basilar arteries is provided by the significant attenuation of VIP-induced vasodilation by VPAC1, VPAC2 or NPR-C antagonists. The VPAC1 and VPAC2 agonists and antagonists used in the present study were developed using rat and human VPAC1 and VPAC2 receptors expressed in Chinese hamster ovary cells using binding studies and adenylate cyclase activation assays (Gourlet et al, 1997a—c; Moreno et al, 2000). The compounds have high potency and specificity as determined from inhibition curves of tracer ([125I]VIP) binding. Thus, the VPAC1 agonist had an IC50 = 1 nmol/L on hVPAC1 and ≥30,000 nmol/L on hVPAC2. The VPAC1 antagonist had an IC50 = 2 nmol/L on hVPAC1 and 3,000 nmol/L on hVPAC2. The VPAC2 agonist Ro-25–1553 had an IC50 = 1 ± 1 nmol/L on hVPAC2 compared with an IC50 = 800 ± 100 nmol/L on hVPAC1. The VPAC2 antagonist PG-99–465 had an IC50 = 2 ± 1 nmol/L on hVPAC2 compared with an IC50 = 200 ± 30 nmol/L on hVPAC1. The current study is the first to utilise selective VPAC ligands, and shows their selectivity for porcine VPAC1 and VPAC2 receptors. The NPR-C receptor agonist and antagonist used in this study also showed their selectivity for the porcine NPR-C receptor. These ligands have previously been shown to be selective for rabbit and guinea pig NPR-C receptors (Murthy et al, 1998, 2000).
In conclusion, the vasodilator effect of exogenously applied VIP in porcine basilar arteries involves NO generation and appears to be a mixture of the effects mediated by VPAC1, VPAC2 and NPR-C receptors. Neurally released VIP induces relaxation of smooth muscle cells by activation of VPAC2 receptors. The NO-dependent component of VIP-induced relaxation of porcine basilar arteries is mediated by VPAC1 receptors, which generate NO in endothelial cells and NPR-C receptors, which generate NO in nitrergic nerves. The characteristic location of each receptor suggests that each has a functional specialisation. These findings have relevance to the onset of cerebral vasospasm, as VIP levels have been reported to be decreased in the CSF of patients after subarachnoid haemorrhage (SAH; Juul et al, 1995). In addition, the relaxation of isolated rabbit basilar arteries in response to exogenous VIP is reduced after experimental SAH, suggestive of changes in the receptor(s) that VIP activates (Tsukahara et al, 1989). Therefore, administration of VIP or a VIP analogue may provide a novel therapeutic approach to reverse arterial narrowing after SAH.
Footnotes
Acknowledgements
We thank S Schulz for supplying the VPAC1 and VPAC2 antibodies and K Omori for the NPR-C antibody. We also thank P Robberecht for kindly supplying the VPAC1 and VPAC2 ligands. We are also indebted to AA Preston for his technical assistance.