
Abstract
The platelet-derived lysophospholipid sphingosine-1-phosphate (S1P) is present in blood plasma and is one of the most potent growth factors displaying proangiogenic activity towards endothelial cells (EC) derived from various tissues. The paracrine regulation of brain angiogenesis by platelet-derived growth factors is, however, poorly understood. In the present study, we assessed the role of S1P on brain EC migration and tubulogenesis, using rat brain-derived (RBE4) EC as an in vitro model. We show that S1P inhibits brain EC migration and tubulogenesis, while it displays proangiogenic activity towards noncerebral EC. Overexpression of the S1P receptor S1P-1 in RBE4 cells potentiated all of the S1P-mediated events. We also show that the lack of expression of MT1-MMP, a membrane-bound matrix metalloproteinase that is thought to cooperate with S1P in tubulogenic processes, may explain the antiangiogenic activity of S1P on brain vasculature. Altogether our results support the hypothesis of a tissue-specific, antiangiogenic role of S1P in the brain, which may help to stabilize the cerebral vasculature and thus have crucial impact on the setting and regulation of normal brain vascularization.
Introduction
Brain vascularization is performed through two different mechanisms, vasculogenesis (Mato et al, 1989) and angiogenesis (Plate, 1999). The latter is a physiologic process that is fundamental to normal healing, reproduction, and embryonic development (Zadeh and Guha, 2003). However, angiogenic processes have also been implicated in the pathogenesis of a wide variety of disorders, including primary and metastatic brain tumors, aneurysms, arteriovenous malformations, and cavernous malformations (Harrigan, 2003). Angiogenesis is initiated by the release of proteases from activated endothelial cells (EC), leading to the degradation of the basement membrane, migration into the interstitial space, with subsequent EC proliferation and differentiation into mature blood vessels. Each of these processes is tightly regulated through the complex interplay of endogenous factors that promote and inhibit angiogenesis (Folkman, 2003). Among them, vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF) have been shown to promote strong proangiogenic activity in EC (Distler et al, 2003).
The specific process of brain angiogenesis has been shown to be sustained by several growth factors, such as VEGF, bFGF, platelet-derived growth factor (PDGF) and transforming growth factor-β (TGF-β) (Harrigan, 2003; Niimi, 2003). Numerous bioactive lipids have also been identified as potent angiogenesis modulators, including platelet-derived lysophospholipids (English et al, 2002). The prominent members of the family of lysophospholipids (LPL) are the structurally related molecules sphingosine-1-phosphate (S1P) and lysophosphatidic acid (LPA) that are secreted by activated platelets (Xu et al, 2003). Several studies have documented the effect of LPL on the proliferation properties of cell types such as EC or fibroblasts (Goetzl et al, 2000; Panetti, 2002). LPL act through homologous G protein-coupled receptors (GPCRs), known as the Edg (endothelial differentiation genes) receptors, but recently named S1PR and LPAR (Goetzl and An, 1998). Sphingosine-1-phosphate preferentially activates EDG-1/S1P-1, EDG-5/S1P-2, EDG-3/S1P-3, EDG-6/S1P-4, and EDG-8/S1P-5, whereas LPA activates EDG-2/LPA-1, EDG-4/LPA-2, and EDG-7/LPA-3 (Anliker and Chun, 2004; Chun et al, 2002). Moreover, S1P may also specifically act as an intracellular second messenger (Spiegel and Milstien, 2003a).
Platelet-derived lysophospholipid S1P is a lysosphingophospholipid present in blood plasma, largely in albumin- and lipoprotein-bound forms (English et al, 2000; Yatomi et al, 2000). The S1P serum concentration ranges from approximately 0.2 to 1 μmol/L (Okajima, 2002). This bioactive sphingolipid metabolite is generated through the phosphorylation of sphingosine by sphingosine kinase (Ancellin et al, 2002) and has been shown to elicit a great variety of responses, including stimulation of cell proliferation, survival, and cytoskeletal reorganization (Payne et al, 2002; Spiegel and Milstien, 20030b). One of the most widely studied functions of S1P is the regulation of the cell migration process and its role in angiogenesis, as it stimulates migration and promotes tubulogenesis in EC (English et al, 2000; Panetti, 2002).
However, little is known about the role of S1P in regulating the angiogenic process in brain. Because EC migration and tubulogenesis represent crucial steps of angiogenesis, we investigated how the motility of brain-derived EC can be modulated by S1P. In this work, we show that S1P acts as a specific inhibitor of brain EC migration and tubulogenesis in vitro. Moreover, we show that the S1P-1 receptor specifically mediates the S1P antiangiogenic effects on brain endothelium. We also show that membrane-type 1-matrix metalloproteinase (MT1-MMP), a membrane-bound matrix metalloproteinase (MMP) that is highly involved in the angiogenic process, is not expressed in brain EC. Finally, we establish that the absence of MT1-MMP in brain EC, which usual function is to cooperate with S1P to trigger tubulogenic processes in noncerebral EC (Langlois et al, 2004), can explain the antiangiogenic role of S1P observed in vitro.
Materials and methods
Antibodies and Chemicals
TriZOL reagent, trypsin, geneticin (G418), penicillin, and streptomycin were from Invitrogen (Burlington, Ontario, Canada). Fibronectin was from Roche (Mississauga, Ontario, Canada). Type 1 collagen was extracted from rat tail tendon by standard method (Silver and Trelstad, 1980). Vitronectin was prepared as previously described (Yatohgo et al, 1988). Recombinant PDGF, VEGF, and granulocyte–macrophage colony-stimulating factor (GM-CSF) were from R&D Systems (Minneapolis, MN, USA). S1P, LPA, concanavalin A (ConA), cytochalasin D (CytoD), EDTA, agarose, gentamycin, laminin-1, sodium dodecyl sulfate (SDS), trypsin for EC and bovine serum albumin (BSA) were purchased from Sigma (Oakville, Ontario, Canada). bFGF was from Upstate Biotechnology (Lake Placid, NY, USA). Epithelial growth factor (EGF) and Matrigel™ were from BD Biosciences Labware (Bedford, MA, USA). Anti-human MT1-MMP polyclonal antibody (raised against the hinge region), anti-TIMP-2 polyclonal antibody, and the enhanced chemiluminescence-Western blot kit were from Chemicon International (Temecula, CA, USA).
Cells and Culture Media
Dulbecco's modified Eagle's medium (DMEM), MEM, α-MEM medium, MDCB 131 medium, Ham's F12 Nutrient Mixture and
Morphogenic Differentiation Assay
Matrigel™ was plated in 96-well plates and allowed to gel for 2 h at 37°C before cell seeding. RBE4 cells (5 × 104) were added to the Matrigel™ in serum-free media in the presence or absence of S1P and incubated at 37°C for 18 h. Capillary-like structures were analyzed microscopically and photographs (50 x) taken with a Retiga 1300 camera connected to a Nikon Eclipse TE2000-U microscope. The capillary-like structures formed in the gel were quantified by analysis of digitized images to determine the segment length of the capillary-like network, using the Northern Eclipse analysis software (Empix Imaging Inc., Mississauga, Ontario, Canada). Results are expressed as percentage of control cells, and are the means of at least three independent experiments. Error bars represent SD of values.
cDNA Constructs and Cell Transfection
A plasmid encoding S1P-1-GFP chimera was generously provided by Dr Timothy M. Palmer. The cloning of human MT1-MMP cDNA has been described previously (Annabi et al, 2001). RBE4 cells were transiently transfected with cDNA plasmids by using the nonliposomal formulation FUGENE-6 transfection reagent (Boehringer Mannheim, Laval, Quebec, Canada). Transfection efficiency was confirmed by Western blotting. All experiments involving transfected cells were performed 36 h after transfection. Mock transfections of RBE4 cells with pcDNA (3.1 +) expression vector alone were used as controls.
Cell Adhesion Assays
Adhesion assays were performed as previously described (Rigot et al, 1998). Briefly, wells were coated with purified extracellular matrix (ECM) protein solutions for 2 h at 37°C, then blocked with a solution of PBS/BSA 0.5%. Matrix concentrations used were as follows: Type-I collagen, fibronectin, laminin-1, vitronectin, elastin (10 μg/mL), fibrin (0.25% w/v in NaOH 1 N), hyaluronan (1 mg/mL) and casein (0.25% w/v in PBS). Cells were then harvested in single cell suspension by brief trypsinization, added to wells precoated with purified ECM proteins, and allowed to adhere to the substrata for 2 h at 37°C. After washing, adherent cells were stained with a solution of 0.1% crystal violet/MetOH 20% and lysed with 1% SDS. Absorbance was then measured at 600 nm. All assays were performed in triplicate and data are the means of at least three independent experiments. Error bars represent s.d. of values.
Migration Assays
To assess the EC migratory potential, the lower face of Transwells (8 μm pore size; Costar, Acton, MA, USA) was precoated with type-1 collagen, laminin-1, vitronectin or fibronectin, at a concentration of 10 μg/mL, for 2 h at 37°C. The Transwells were then assembled in a 24-well plate (Fisher Scientific Ltd, Nepean, Ontario, Canada), the lower chambers were filled with 600 μL of serum-free medium, and 100 μL of cells (106 cells/mL) were inoculated onto the upper chamber of each Transwell. Various concentrations of growth factors in serum-free medium were added to the lower chambers of the Transwells. The plate was then placed at 37°C in 5% CO2/95% air for 1 h. Cells that had migrated to the lower surface of the filters were fixed and stained with 0.1% crystal violet/20% (v/v) MeOH. All assays were performed in triplicate. The migration was quantified using computer-assisted imaging and data were analyzed using the Northern Eclipse software. Migration data are expressed as a mean value of, at least, three independent experiments. Error bars represent s.d. of values.
Isolation of EC Fractions from Brain Tissue
The isolation of EC from normal male Lewis rat (Charles River, Quebec, Canada) brains has been performed as previously described (Regina et al, 2003). All animal experiments were evaluated and approved by the Institutional Comity for Good Animal Practices (UQAM, Montréal, Quebec, Canada). Magnetic cell sorting (MACS) was performed using microbeads, a MidiMacs separation unit and positive selection MACS columns from Miltenyi Biotec (Auburn, CA, USA). Mouse anti-PECAM-1 antibody that was linked to microbeads was from Cedarlane Laboratories (Hornby, Ontario, Canada).
Total RNA Isolation and Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
Total RNA was extracted from cells using the TriZOL Reagent. RT-PCR reactions were performed using the SuperScript™ One-Step RT-PCR with Platinum® Taq Kit from Invitrogen (Burlington, Ontario, Canada). Polymerase chain reaction conditions have been optimized so that the gene products were examined at the exponential phase of the amplification. Polymerase chain reaction products were resolved on 2% (w/v) agarose gels containing 1 μg/mL ethidium bromide.
Immunoblotting Procedures
Cells were homogenized in lysis buffer. An amount of 20 μg of protein from control or transfected cells were separated by SDS–polyacrylamide gel electrophoresis (PAGE). After electrophoresis, proteins were electrotransferred to polyvinylidene difluoride membranes, which were then blocked overnight at 4°C with 5% nonfat dry milk in Tris-buffered saline (150 mmol/L NaCl, 20 mmol/L Tris–HCl, pH 7.5) containing 0.3% Tween 20 (TBST). Membranes were further washed in TBST and incubated with the primary antibodies (1/1,000 dilution) in TBST containing 3% BSA, followed by a 1 h incubation with horseradish peroxidase-conjugated anti-rabbit IgG (1/5,000 dilution) in TBST containing 5% nonfat dry milk. Immunoreactive material was visualized by enhanced chemiluminescence.
Gelatin Zymography
Gelatinolytic activity in culture media from monolayer cultures was detected by gelatin zymography as previously described (Beaulieu et al, 1999). Briefly, an aliquot (20 μL) of the culture medium was subjected to SDS/PAGE using a 7.5% (w/v) acrylamide gel containing 0.1 mg/mL gelatin. The gels were then twice incubated for 30 mins at room temperature (25°C) in 2.5% (v/v) Triton X-100 to remove SDS and rinsed five times in double-distilled water. The gels were further incubated at 37°C for 20 h in 20 m
Treatment with Antisense Oligonucleotides
Antisense phosphorothioate oligodeoxyribonucleotides (PTO) against S1P-1, S1P-3, and their respective scrambled controls were custom synthesized based on published sequences (Lee et al, 1999a; Liu et al, 2001). The sequences of the oligonucleotides used are the following: S1P-1 antisense, 5'-GAC GCT GGT GGG CCC CAT-3'; scrambled S1P-1, 5'-TGA TCC TTG GCG GGG CCG-3'; S1P-3 antisense, 5'-CGG GAG GGC AGT TGC CAT-3'; scrambled S1P-3, 5'-ATC CGT CAA GCG GGG GTG-3'. To reduce S1P-1 and S1P-3 gene expression, RBE4 cells were grown to approximately 60% of confluence and incubated with 200 nM of either the scrambled controls or the S1P-1 or S1P-3 antisense oligonucleotides using Fugene-6 transfection reagent. After 72 h of incubation, cells were used for migration and morphogenic assays.
Semiquantitative Real-Time PCR Analysis
Real-time PCR analysis was performed in a two-step manner. cDNA synthesis and real-time detection were performed in a Perkin-Elmer Gene Amp PCR system and with a M x 4000 system (Strategene, La Jolla, CA, USA) respectively. Total RNA was extracted as described above. S1P-1 mRNA (5 μg) was reverse transcribed in a final volume of 20 μL containing 1 μL of 10 mmol/L dNTP, 4 μL BRL 5 x buffer, 0.5 μL oligo-(dT)12–18 primer (0.5 μg/μL, Invitrogen), 2 μL of 0.1 mol/L DTT, and 1 μL of 200 U/μL M-MLV reverse transcriptase (Invitrogen) for 1 h at 37°C. The reaction was stopped by heating at 95°C for 5 min. The QuantiTect SYBR Green PCR kit (Qiagen, Mississauga, Ontario, Canada) was used in subsequent PCR assays according to the manufacturer's protocol. Gene expression was compared by the determination of target quantities in experimental samples relative to a calibrator sample (control), and expression levels were corrected for a normalizer gene, 16S rRNA. Single-strand cDNA (2 μL) was used for real-time PCR to amplify a 546-bp fragment of S1P-1 cDNA with the complementary sense primer 5'-GTCCGGCATTACAACTACAC −3’ and the antisensee primer 5'-TATAGTGCTTGTGGTAGAGC-3'. For a 107-bp fragment of the housekeeping gene ribosomal protein S16 cDNA, the antisense primer was 5'-GCTACCAGGGCCTTT***GAGATG-3', the sense primer, 5'-AGGAGCGATTTGC***TGGTGTGG-3'. Thus, 2 μL of cDNA was amplified with specific primers at a final concentration of 1 μmol/L. All real-time RT-PCR experiments were performed in duplicate and were conducted with an initial denaturing interval (95°C, 15 mins) and then with 40 sequence cycles for S1P-1 and S16: 94°C (30 secs), 55°C (45 secs), and 72°C (30 secs). Melting-curve analysis was also performed to control for the absence of primer dimers.
Results
Dependency of RBE4 Cell Adhesion and Migration on Substrate
Although RBE4 cells have been extensively used as an in vitro model for the blood–brain barrier (Chishty et al, 2002; Lagrange et al, 1999; Mroczkowska et al, 2000), relatively little data are available concerning their motility properties. Since we were interested in several distinct processes involved in angiogenesis, we first characterized the adhesive and migratory properties of these cells on various purified ECM proteins. We determined that RBE4 cells exhibited approximately seven fold higher adhesion to fibronectin than on any other matrix (Figure 1A). We used modified Boyden chambers to assess the ability of RBE4 to migrate towards various purified ECM proteins including type I collagen, laminin, vitronectin, and fibronectin. In accordance with the adhesion test results, RBE4 cell migration was significantly higher towards fibronectin (10-fold higher) than towards any other ECM protein tested (Figure 1B). All subsequent migration experiments were consequently performed on fibronectin-coated filters.
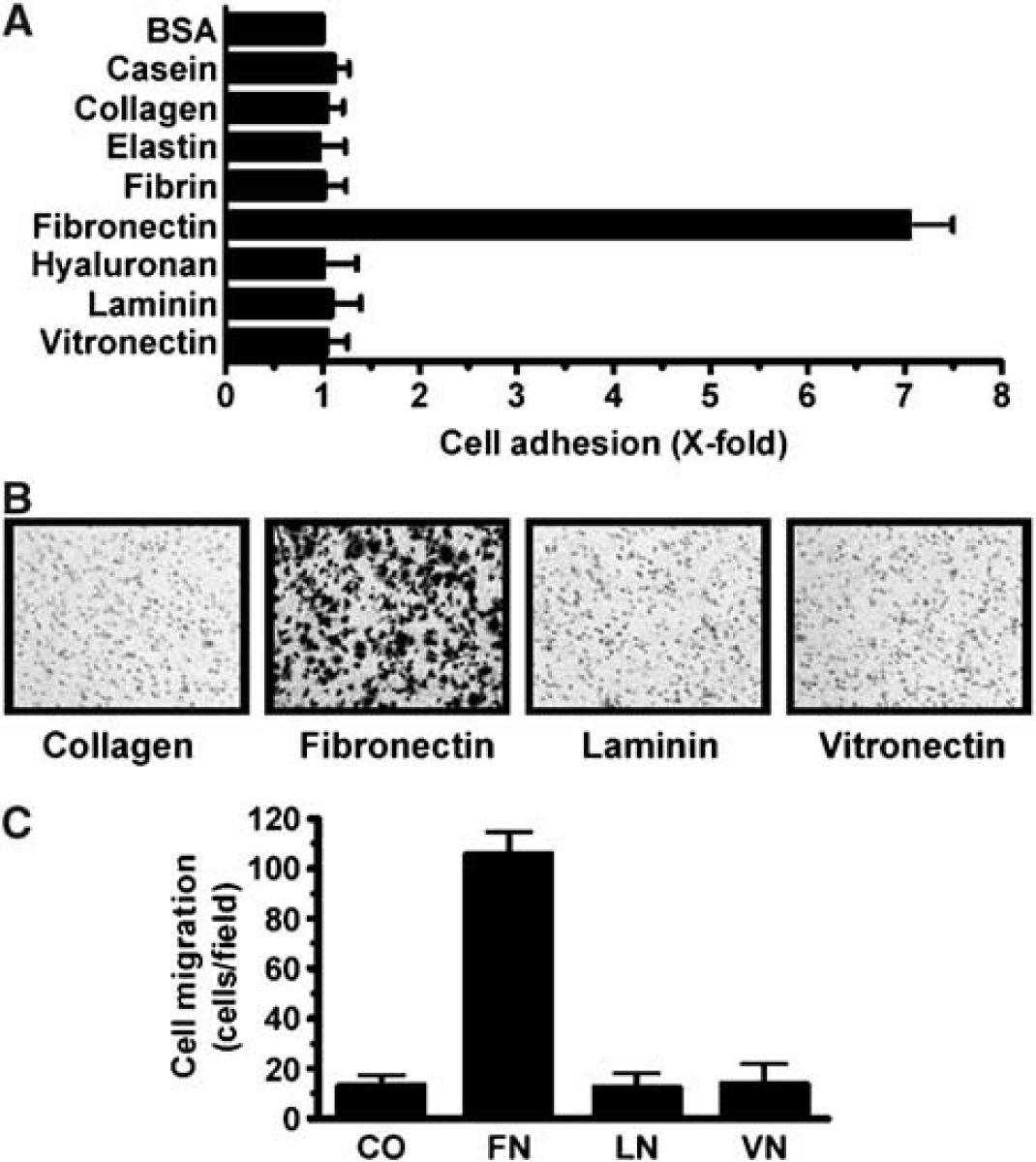
Fibronectin is the most potent ECM substrate for RBE4 cell adhesion and migration. (
Modulation of RBE4 Cell Migration by Various Growth Factors
Because brain EC are exposed to the effects of various growth factors in vivo, we assessed whether some of these factors can affect RBE4 cell migration. RBE4 were thus seeded on fibronectin-coated Transwells, and allowed to migrate for 1 h towards serum-free medium supplemented with either serum (FBS 10%), S1P, PDGF, VEGF, TNFα, bFGF, or GM-CSF. RBE4 displayed a strong chemotactic migration towards serum, VEGF, bFGF, PDGF, and GM-CSF (Figure 2). In contrast, S1P inhibited RBE4 cell migration by approximately 50%. Because this result is the first to report a potent role for S1P on the inhibition of brain EC migration, we decided to further characterize the effect of S1P on RBE4 cell migration and on tubulogenesis, another crucial event in brain angiogenesis.
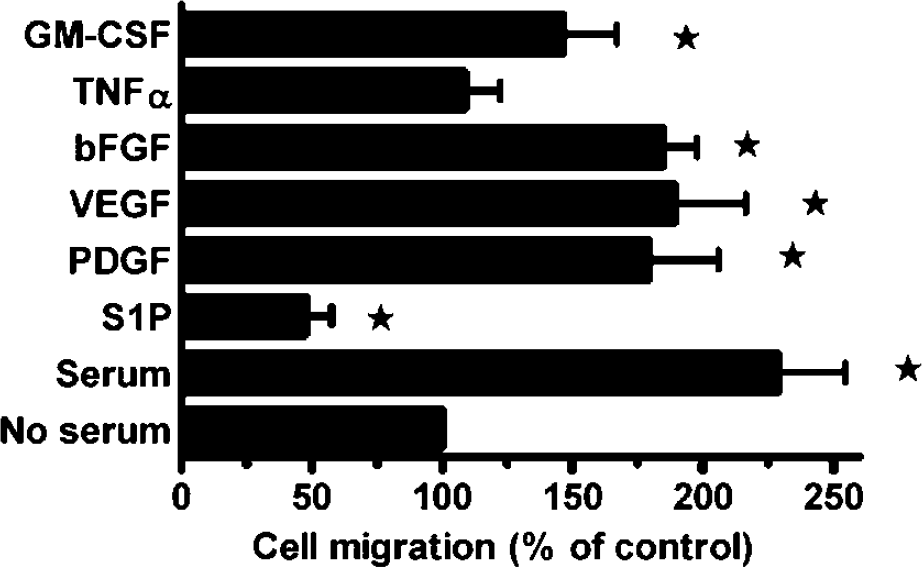
RBE4 cell migration on fibronectin is inhibited by S1P The lower faces of the Transwells were previously coated with 10 μg/mL fibronectin. Cells (105) were seeded on the upper face of Transwells and allowed to migrate for 1 h in a serum-free medium containing one of the following growth factors: S1P (10 μmol/L), PDGF (10 ng/mL), VEGF (10 ng/mL), bFGF (10 ng/mL), TNFα (50 ng/mL) and GM-CSF (50 ng/mL). As a positive control, cells were allowed to migrate towards 10% FBS-containing medium. Cells that migrated to the underside were counted. * indicates statistically significant differences (P<0.05) compared with the control condition (serum-free medium) using Student's t-test.
Modulation of Angiogenic Properties of RBE4 Cells by S1P
Varying S1P concentrations were used to assess RBE4 cell migration and tubulogenesis. As shown in Figure 3A, S1P inhibits RBE4 cell migration in a dose-dependent manner. RBE4 cells were also plated on Matrigel™ with varying S1P concentrations and incubated for 18 h. S1P was also found to inhibit RBE4 in vitro tubulogenesis in a dose-dependent manner (Figure 3B). The unexpected results obtained with RBE4 cells were the opposite of those reported for noncerebral EC. We thus investigated whether the precise tissue origin of EC could determine the different effects of S1P on angiogenesis.
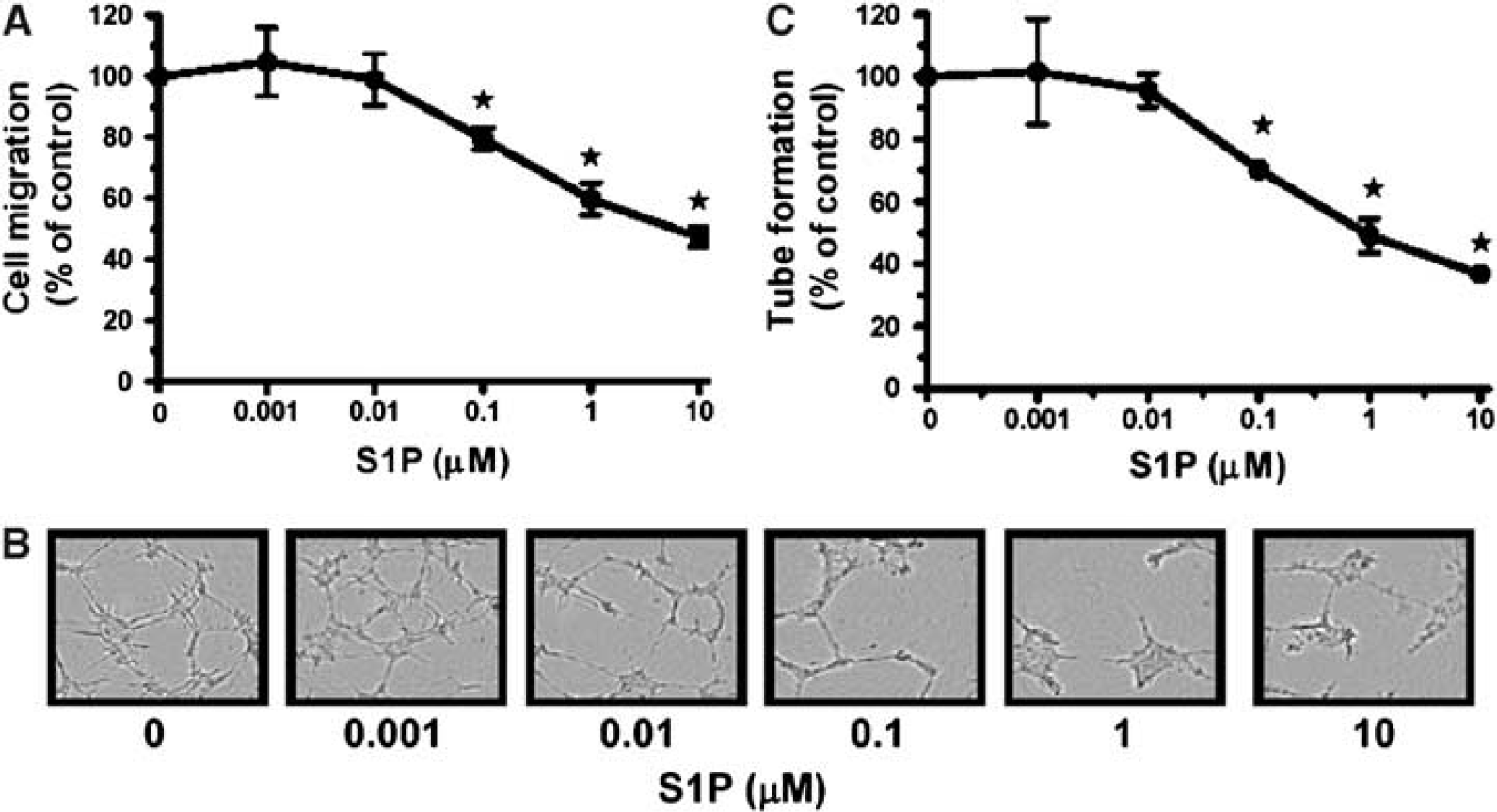
S1P inhibits RBE4 cell migration and in vitro tubulogenesis. (
Effect of S1P on the Migration of Cerebral and Noncerebral EC
We tested the chemotactic activity of S1P on the migratory potential of various EC lines: HUVEC (Umbilical Vein), HMEC-1 (Dermal Microvascular), BAEC (Aortic), RBE4 (Cerebral), GP8.39 (Cerebral) and BBCEC (Cerebral). As shown in Figure 4, S1P stimulated the migration of all EC lines that originated and were isolated from outside the brain. However, brain-derived EC (RBE4, GP8.39 and BBCEC) migration was inhibited by S1P (between 50% and 55%). In light of these results, and because it has already been shown that S1P promotes noncerebral angiogenesis, we can conclude that S1P is a specific inhibitor of brain angiogenesis.
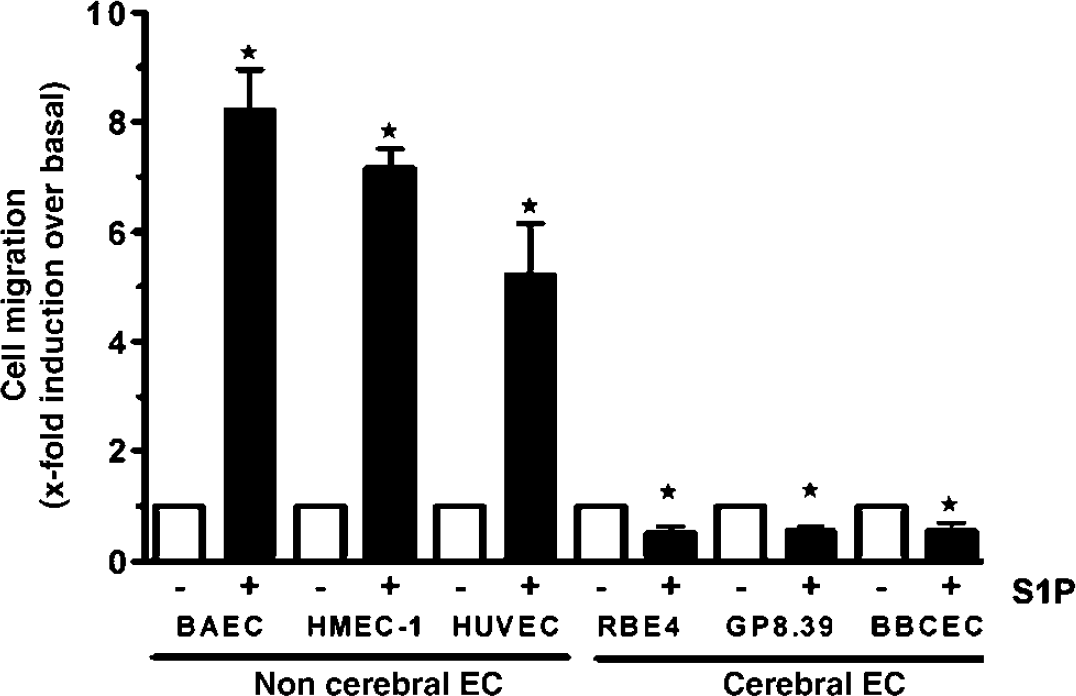
S1P specifically inhibits the migration of brain derived-EC. Noncerebral EC (BAEC, HMEC-1, and HUVEC) and cerebral EC (RBE4, GP8.39, and BBCEC) were harvested by brief trypsinization. Migration assays were performed on fibronectin-coated Transwells. Cells (105) of each cell line were added to the wells and allowed to migrate in a serum-free medium containing or not 10 μmol/L S1P Results are expressed as the fold-induction of cell migration over basal migration of untreated cells.
Role of the Receptor S1P-1 in the Antiangiogenic Effect of S1P in Brain EC
Because S1P-1 and S1P-3 are the two main receptors that mediate S1P angiogenic activity in EC, we wished to assess their contribution to the S1P-mediated inhibition of RBE4 cell migration and tubulogenesis. We used antisense oligonucleotides to downregulate S1P-1 and S1P-3 gene expression in RBE4 cells. Downregulation of S1P-1 and S1P-3 gene expression was confirmed by RT-PCR (Figure 5A). Interestingly, S1P-1 downregulation antagonized the S1P-mediated inhibition of RBE4 cell migration (Figure 5B). However, no significant difference on the S1P-inhibitory effect was observed between the control RBE4 cells and the S1P-3-lacking RBE4 cells. Consequently, we concluded that the inhibition of RBE4 cell migration by S1P is mainly mediated by the S1P-1 receptor. Semiquantitative real-time PCR experiments were performed to determine the level of S1P-1 mRNA in RBE4 cells after incubation with antisense oligonucleotides. A mean decrease of 48% of the S1P-1 mRNA in RBE4 treated with the antisense oligonucleotides, was observed when compared with the untreated cells (data not shown). This correlates and confirms the downregulation of the S1P-1 mRNA observed by RT-PCR (Figure 5A). Furthermore, capillary-like tube formation of S1P-1-downregulated cells was also less effective (Figure 5C). Thus, we concluded that S1P-1 also regulates the antitubulogenic properties of S1P on brain EC.
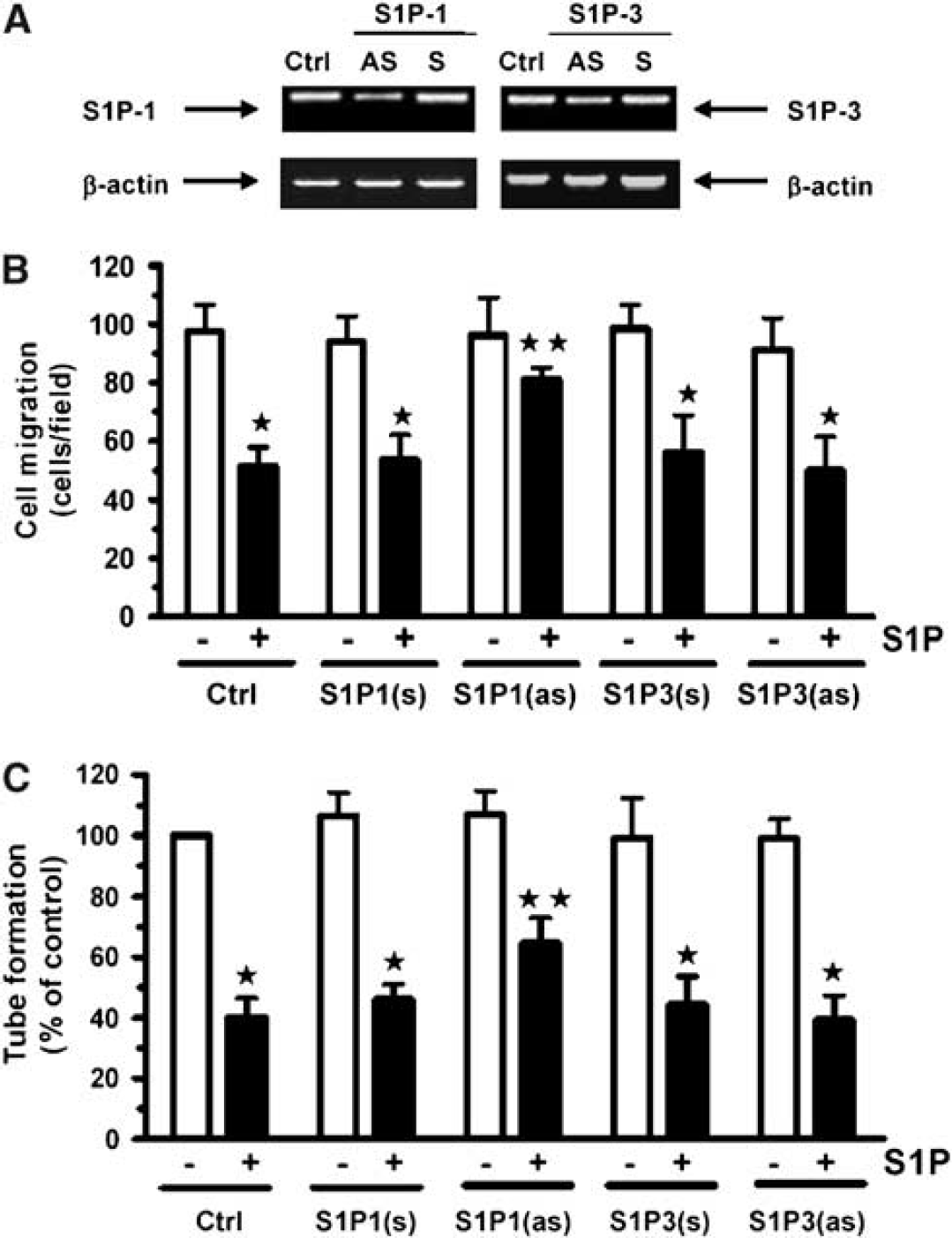
S1P-1 transduces the antiangiogenic effects of S1P in brain endothelial cells. RBE4 cells were transiently treated with 200 nmol/L of antisense (AS) oligonucleotides against S1P-1, S1P-3, or with their scrambled controls (S), and used 72 h after transfection. (
Effect of the Overexpression of S1P-1 on the Response of Brain EC to S1P
Because S1P-1 appeared to be the main S1P receptor involved in the S1P-mediated inhibition of RBE4 cell migration, we overexpressed S1P-1 in RBE4 by transfection with a plasmid encoding for the fusion protein S1P-1-GFP and tested whether overexpression of S1P-1 could potentiate the S1P effect on RBE4 cell migration and tubulogenesis. Using fluorescence microscopy, we observed broad expression of the S1P-1-GFP fusion protein in RBE4 cells (Figure 6A). S1P-1-transfected RBE4 cells were allowed to migrate on fibronectin with varying concentrations of S1P. As expected, cells that overexpressed S1P-1 underwent S1P-mediated migration inhibition at a decreased S1P concentration than did control cells (Figure 6B). Indeed, S1P-1-transfected RBE4 cell migration exhibited 50% inhibition at a S1P concentration of 0.027 μmol/L compared with 0.164 μmol/L for the control cells. Thus, S1P-1 overexpression sensitized RBE4 cells to the inhibitory effect of S1P. The same result was observed in tubulogenesis assays where the capillary-like structure formation of S1P-1-transfected RBE4 cells was more inhibited than in control cells (Figure 6C), as quantified in Figure 6D. This clearly shows that S1P-1 triggers a specific transduction pathway leading to S1P-mediated brain antiangiogenic activity.
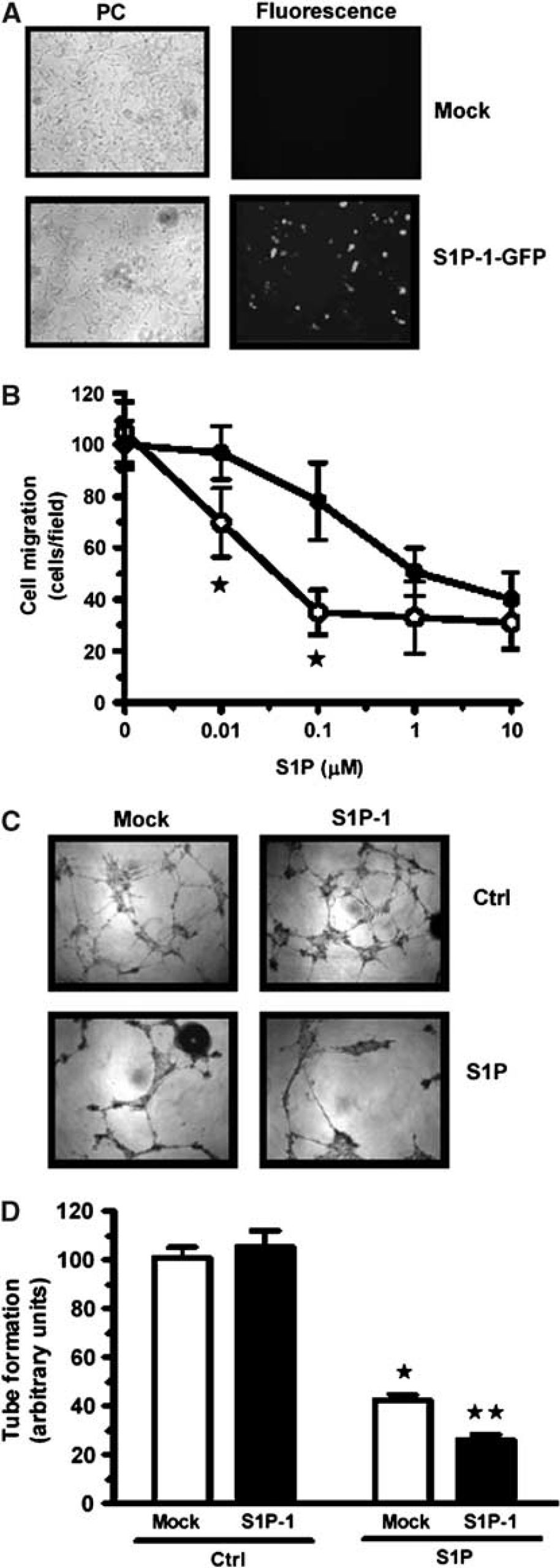
Overexpression of S1P-1 in RBE4 cells potentiates the S1P antiangiogenic effect. RBE4 cells were transiently transfected with a cDNA plasmid encoding S1P-1-GFP or the pcDNA (3.1 +) empty plasmid (mock). (
Expression of MT1-MMP in Rat Brain-Derived EC
MMPs have been shown to play a crucial role during angiogenesis. In particular, it has been shown that MT1-MMP, the best-characterized member of the MT-MMP family, is functionally involved in EC migration and tubulogenesis, as well as in the proMMP-2 activation process (Lafleur et al, 2002, 2003). MT1-MMP-dependent pro-MMP-2 activation in RBE4 cells was tested with concanavalinA (ConA) or cytochalasinD (CytoD) (Ailenberg and Silverman, 1996; Yu et al, 1997), in parallel with U-87 glioblastoma cells. The extent of proMMP-2 activation in the conditioned media was assessed by gelatin zymography, as described in the Materials and methods section. As shown in Figure 7A, ConA and CytoD treatments clearly induced proMMP-2 activation in U-87 cells, but failed to do so in RBE4 cells. Because the MT1-MMP-dependent proMMP-2 activation mechanism also involves TIMP-2, a tissue inhibitor of MMPs, we assessed the presence of this molecule in the conditioned media of various cell lines. As shown by Western blotting, TIMP-2 was found to be expressed in all cell lines tested, including RBE4 (Figure 7B). We then assessed transcription from the MT1-MMP gene by RT-PCR in various cell lines. MT1-MMP mRNA expression was absent in all the brain EC lines tested (RBE4, GP8.39), while MT1-MMP was clearly expressed in human glioma cells, rat CNS-1 glioma cells and non-cerebral human EC lines (HUVEC, HMEC-1) (Figure 7C).
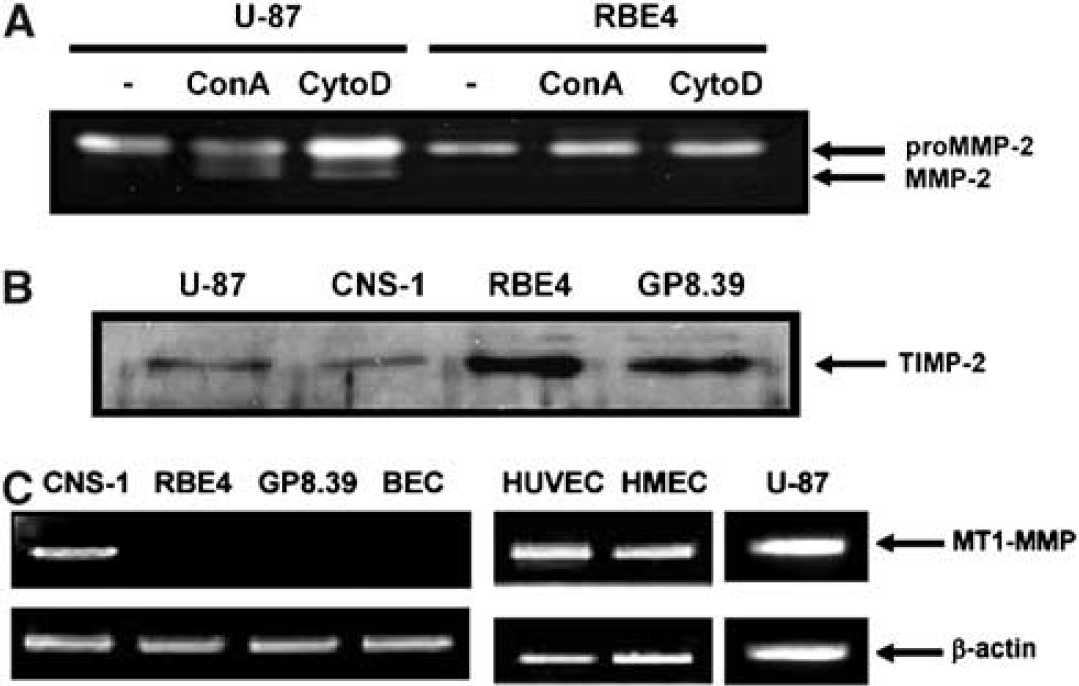
Pro-MMP-2 activation, TIMP-2, and MT1-MMP expression in rat brain endothelial cells. (
Effect of the Expression of MT1-MMP on the Response of Brain EC to S1P
We hypothesized that the absence of MT1-MMP in brain EC could explain the antiangiogenic activity of S1P since intracellular crosstalk has been reported by us between S1P signaling and MT1-MMP expression (Annabi et al, 2003). Later, we have also shown that MT1-MMP cooperates with S1P to induce EC migration and tubulogenesis (Langlois et al, 2004). To test this possibility, we transiently overexpressed MT1-MMP in RBE4 cells. The transfection efficiency was measured by Western Blot detection of MT1-MMP in RBE4 cell lysates (Figure 8A). The overexpression of MT1-MMP in RBE4 did not change the basal migratory potential of these cells (Figure 8B), which suggests that RBE4 cell migration does not depend on the presence of MT1-MMP. However, when transfected RBE4 cells migrated in the presence of S1P, we observed that the overexpression of MT1-MMP reversed the S1P-mediated inhibition of RBE4 cell migration (Figure 8B). The same result was observed for capillary-like structure formation ability on Matrigel™ (Figure 8C). Consequently, we conclude that the lack of MT1-MMP in brain EC explains the antiangiogenic effect of S1P.
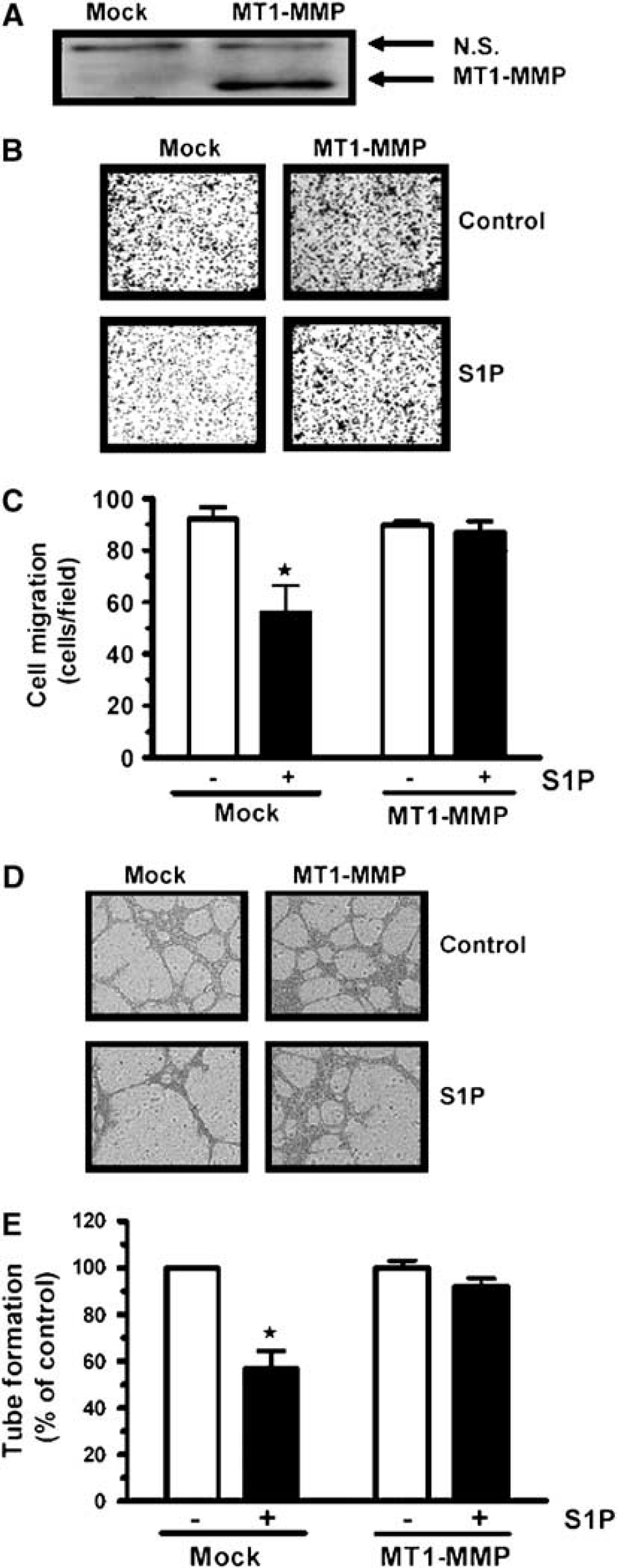
Overexpression of MT1-MMP antagonizes S1P-mediated antiangiogenic effects on RBE4 cells. RBE4 cells were transfected with MT1-MMP cDNA or the corresponding empty vector (mock). (
Discussion
Numerous molecules have been identified as playing crucial roles in the control of brain angiogenesis (Zadeh and Guha, 2003). These include proangiogenic factors such as VEGF, bFGF, and PDGF, as well as antiangiogenic factors (Distler et al, 2003). All these factors have been characterized in terms of modulating crucial cellular processes implicated in angiogenesis such as cell growth, cell differentiation or cell migration (Liekens et al, 2001). Lysophospholipids have also been identified as key modulators of the angiogenic process (Hla et al, 2001). They have been shown to regulate numerous cellular mechanisms leading to angiogenesis (Panetti, 2002). For instance, all of the studies of S1P activity in EC have indicated a proangiogenic role for this molecule (Lee et al, 1999b). However, we present molecular and cellular evidences establishing that S1P can display tissue-specific antiangiogenic activities. Indeed, we show that S1P inhibits in vitro brain-derived EC migration and tubulogenesis. The specificity of the brain EC response to S1P stimulation could be explained by their unique function in the setting of the blood–brain barrier. In light of our results, the specific response of brain EC towards S1P clearly confirms that EC display phenotypical and molecular variability depending on their tissue location. We have previously provided molecular evidence of phenotypic distinction between tumoral and normal brain vasculature as well as between EC isolated from intracerebral and subcutaneous brain tumor models (Regina et al, 2003). These results indicated that the EC phenotype depends on interactions with surrounding cells but also with the ECM microenvironment. Thus, in the brain, EC experience the influence of various growth factors, which leads to the setting of a specific cerebral phenotype.
One pathophysiologic consequence of the brain vascular phenotype might be found in inflammation, during which platelets secrete high quantities of S1P (English et al, 2000; Yatomi et al, 2000). The first event of the inflammatory response is a strong vasodilatation. In peripheral vessels, S1P triggers the activation of EC, the release of NO and the relaxation of pericytes, leading to vasodilation (Dantas et al, 2003; Nofer et al, 2004). However, S1P triggers a specific vasoconstriction of brain vessels, and could thus block the inflammatory process in the brain (Coussin et al, 2002; Salomone et al, 2003; Tosaka et al, 2001). Thus, it appears that the cerebral microenvironment strongly influences the endothelial response to S1P. Interestingly, S1P has been quantified in numerous tissues of various species (Edsall and Spiegel, 1999; Murata et al, 2000; Yatomi et al, 1995) and a recurrent characteristic is its high quantity in brain. The specific localization of S1P in brain suggests that it could have a role during neurogenesis and brain angiogenesis (McGiffert et al, 2002). Our study presents strong evidence suggesting that S1P affects cerebral EC angiogenic properties, and indicates that S1P could stabilize the brain vasculature through its antiangiogenic effect on brain EC. This raises an attractive hypothesis of a possible protective role for S1P in brain, as already described in lung (Peng et al, 2004; Schaphorst et al, 2003).
At the molecular level, we determined that the lack of MT1-MMP at the brain EC surface was responsible for the antiangiogenic activity of S1P. The role of MT1-MMP in the S1P-mediated angiogenic activity has already been documented by us (Annabi et al, 2003; Langlois et al, 2004). Sphingosine-1-phosphate has been shown to be proangiogenic for numerous cell lines. It has also been shown that S1P increases HUVEC, HMEC-1, BAEC, and bone marrow-derived stem cell (BMSC) cell migration and tubulogenesis (Annabi et al, 2003; Lee et al, 1999b). One common feature in S1P-responsive EC is that they all express MT1-MMP. The brain EC, which do not express MT1-MMP, are characterized by their antiangiogenic response to S1P. It is thus tempting to suggest that the expression of MT1-MMP in EC modulates the angiogenic potential of EC in response to S1P.
S1P has already been shown to affect capillary-like tube formation in various noncerebral EC (Annabi et al, 2003; Bayless and Davis, 2003; Lee et al, 1999b). We have shown that BAEC, which express low levels of MT1-MMP, display a strong migration in response to S1P. Using the same cells, we have shown that BAEC do not undergo morphogenic differentiation in response to S1P (Langlois et al, 2004). Thus, it seems that low expression of MT1-MMP is sufficient to trigger a promigratory response to S1P but is not sufficient to induce endothelial tubulogenesis in response to S1P. Accordingly, overexpression of MT1-MMP in BAEC triggered the EC morphogenic differentiation in response to S1P (Langlois et al, 2004). Then, we can safely state that MT1-MMP expression dictates the angiogenic response to S1P in EC.
The tissue-specific expression of MT1-MMP described here leads to reconsideration of the role of MT1-MMP in endothelial morphogenic differentiation. Numerous papers have shown that MT1-MMP plays a major role in endothelial tubulogenesis (Chiarugi et al, 2000; Hotary et al, 2000; Lafleur et al, 2002). In the present study, we clearly observe that brain EC display potent MT1-MMP-independent tubulogenesis. We cannot, however, preclude that other MT-MMP isoforms may compensate for the absence of MT1-MMP (Lafleur et al, 2002). It is well known that a variety of MT-MMPs exist, with different tissue distributions. Our study supports previous results concerning the specialized location of certain MT-MMPs. The absence of MT1-MMP could be one of the molecular characteristics of the brain EC phenotype that may regulate some of the blood–brain barrier functions. Other MT-MMPs have been shown to be expressed in a tissue-dependent manner. For instance, MT5-MMP have been described as being mainly expressed in brain (Hayashita-Kinoh et al, 2001; Sekine-Aizawa et al, 2001). Thus, assessing the roles of other MT-MMPs, and more particularly MT5-MMP, in cerebral EC should certainly provide new insights in our understanding of the major role that these proteins play in brain angiogenesis.
We have shown that the expression of the MT1-MMP in brain EC antagonizes the specific S1P antiangiogenic activity. The differential effect observed between control and MT1-MMP expressing brain EC clearly shows the ability of MT1-MMP to transduce intracellular signaling pathways regulating angiogenic processes. However, the specific MT1-MMP-mediated transduction pathway, leading to the proangiogenic response to S1P, remains to be elucidated. Altogether the present data revealed the antiangiogenic role of S1P in brain. According to the major role of cerebral endothelium in the setting of the blood–brain barrier, we suggest that S1P, in physiologic conditions, acts as a stabilizing agent of the blood–brain barrier by inhibiting brain EC migration. Because of its dose-dependent activity, S1P should be more active on brain EC in case of inflammation than in physiologic conditions. Consequently, S1P may also participate to a global brain protection process during pathologic conditions through a stronger stabilization of the blood–brain barrier.
Footnotes
Acknowledgements
Borhane Annabi holds a Canada Research Chair in Molecular Oncology (Tier-II).