
Abstract
Generally, L-amino acids are preferably transported into mammalian cells compared with their D-isomers, and only L-amino acids are incorporated into proteins. Former studies, however, indicated that D-[H]proline is accumulated in the brain of mice after injection, while L-[3H]proline is not. We investigated the differential cerebral uptake of the D- and L-isomers of the PET tracer cis-4-[18F]fluoroproline (D-/L-cis-FPro) and of D-/L-[3H]proline (D-/L-Pro) in rats by dual tracer autoradiography and the uptake of D-cis-FPro in two human subjects by PET. The standardized uptake value (SUV) of D-cis-FPro in the cerebral cortex of rats 2 h p.i. was 3.05±1.18 (n=9) versus 0.06±0.01 (n=4) for L-cis-FPro (P<0.001) and 1.29±0.27 (n=4) for D-Pro versus 0.30±0.14 (n=9) for L-Pro (P<0.001). Analysis of the rat brain tissue after injection of D-cis-FPro (n=3) revealed no radioactivity in the proteins but a relevant part in the form of L-trans-FPro. The PET studies yielded a four- to five-fold higher SUV and influx rate constant in the human cortex for D-cis-FPro than for L-cis-FPro. We conclude that D-cis-FPro and D-Pro are preferably transported at the blood–brain barrier compared with their L-isomers and isomerized to the L-form within the brain. Thus, D-Pro in the plasma might be a source of intracerebral L-proline, which has been shown to act as a modulator of excitatory neurotransmission.
Introduction
It was long assumed that only L-amino acids are present in higher animals and that the enantiomers, D-forms, are utilized only in the lower species such as microorganisms and bacteria (Corrigan, 1969). Generally, only L-amino acids are susceptible to enzyme-catalyzed polymerization (translation) to structural and functional peptides and proteins in mammals (Friedmann, 1999). Recent progress in analytical chemistry, however, has enabled the sensitive and selective determination of D-amino acids in mammalian tissues and revealed that substantial amounts of D-amino acids are present in mammals (Hamase et al, 2002). The function and origin of D-amino acids has been investigated in the last decades, but only for D-serine and D-aspartate relevant functional roles have been identified (Wang et al, 2000; Snyder and Kim, 2000).
In 1972, Garweg and Dahnke reported an intensive labelling of the cerebral cortex after intraperitoneal injection of D-[3H]proline (D-Pro) in mice, while after injection of L-[3H]proline (L-Pro) cortical uptake was negligible (Garweg and Dahnke, 1972). Further experiments demonstrated that radioactivity in the brain after injection of D-Pro was due to incorporation of L-Pro into proteins of the cerebral cortex (Garweg and Dahnke, 1973, 1974). The authors postulated the existence of a proline racemase in the cerebral cortex of the mouse converting D-proline to L-proline before utilization in protein synthesis.
Recently 4-[18F]fluoro-L-proline has been introduced as new tracer for PET imaging in humans (Hamacher, 1999; Wester et al, 1999a). Experiments in tumor-bearing mice showed that cis-4-[18F]fluoro-L-proline (L-cis-FPro) accumulates in experimental tumors and is incorporated into proteins (Wester et al, 1999a). Furthermore, it was shown that L-cis-FPro and L-Pro exhibit similar transport characteristics in F98 rat glioma cells, that is, mainly Na+-dependent transport via amino-acid transport systems A and B0,+ (Langen et al, 2002). A study with L-cis-FPro in F98 rat gliomas and human gliomas demonstrated uptake in the tumors but negligible uptake in the normal brain tissue (Langen et al, 2004). Stimulated by the observations of preferred uptake of D-Pro in the brain of rodents in relation to L-Pro, we investigated whether the D-isomer of cis-FPro is also preferably transported across the blood–brain barrier (BBB) in rats and humans in comparison to its L-isomer.
Materials and methods
Radiotracers
D-cis-FPro was prepared via cryptate-mediated n.c.a. nucleophilic 18F-fluorination starting from (2R,4R)-N-Boc-4-p-(toluolsulfonyloxy)proline methylester. N-deprotection and ester hydrolysis take place under acidic conditions in the presence of trifluoromethanesulfonic acid. HPLC purification combined with on-line solid-phase extraction yielded the diastereomerically pure D-cis-FPro with a radiochemical yield of approximately 30% within 90 mins, a radiochemical purity of >98% and a specific radioactivity of >150 GBq/μmol.
D-[3H]proline was obtained commercially with a specific radioactivity of 1.9 TBq/mmol (Biotrend, Cologne, Germany) and L-Pro with a specific radioactivity of 3.7 TBq/mmol (Amersham Buchler, Braunschweig, Germany).
Animal Experiments
In all, 16 male Fisher CDF rats (8 to 12 weeks, weight 250 to 300 g, Charles River Wiga, Sulzfeld, Germany) were examined in this study. The experiments were approved by the district government according to the German Law on the Protection of Animals (Cologne/Germany No. 23.203.2-KFA 10/98).
The animals were kept under standard conditions and free access to food and water. Animals were sedated in an isoflurane atmosphere (2% to 5%) and anesthetized with an intraperitoneal injection of ketamine (100 mg/kg)/xylazine (10 mg/kg) for tracer injection via a jugular vein. A first group of animals (n=4) was injected with a mixture of 80 MBq cis-D-FPro and 5 MBq D-Pro, a second group (n=5) with a mixture of 80 MBq cis-D-FPro and 5 MBq L-Pro and a third group (n=4) with a mixture of 80 MBq cis-L-FPro and 5 MBq L-Pro. At 2 h after tracer injection animals were killed, the brains were removed immediately and frozen in 2-methylbutane at −50°C. Brains were cut in coronal sections (thickness: 7 μm) with a cryostat microtome (CM 3050, Leica, Germany). In two animals blood was sampled, frozen, cut and coexposed with the brain sections to determine the radioactivity concentration in the blood.
Autoradiography
Coronal sections of the brain were placed on phosphor imaging plates (Raytest-Fuji, Straubenhardt, Germany) along with industrial tritium activity standards (Microscales, Amersham Biosciences, Freiburg, Germany) for 3H-studies and with in-house-made calibrated 18F liver paste standards. The first exposition to depict distribution of the 18F-radioactivity was started within 2 h after tracer injection. This exposition was performed on imaging plates insensitive to β−-particles of 3H (BAS-SR 2025, Raytest-Fuji, Straubenhardt, Germany). After decay of 18F (10 half-lives) the brain slices were exposed again to 3H-sensitive imaging plates to obtain 3H-distribution (BAS-TR 2025, Raytest-Fuji, Straubenhardt, Germany). On exposure, the imaging plates were scanned with a high-performance imaging plate reader (BAS 5000 BioImage Analyzer, Raytest-Fuji, Straubenhardt, Germany). Quantitative autoradiograms were generated (nCi/mg wet weight of the tissue) using the software provided by the manufacturer and the known radioactivity concentrations of the standards. Tracer uptake was quantified using a circular regions of interest (ROI) placed on the frontal cortex. Standardized uptake values (SUV) were calculated by normalization of the tissue concentrations in the ROIs to injected dose and body weight, and corrected for the signal of radioactivity from vascular space assuming a cerebral blood volume of 1.63% as determined in decapitated rats (Todd et al, 1993). The tissue sections were stained by cresyl violet, which demonstrates the Nissl-substance and nucleoli.
Analysis of Radiotracer in Brain
Three rats were killed 60 mins after injection of 150 MBq D-cis-FPro, the brains were removed immediately, weighed and their total radioactivity measured. The cerebrum was divided into two rostral and two caudal portions. For analysis of protein incorporation, one rostral and one caudal part were homogenized and lysed 1 h at 4°C in 1% NP40. The lysat was centrifuged and the pellet separated by gel electrophoresis. The second brain half was homogenized in 60% acetonitril, centrifuged, and the radioactivity in the supernatant and the pellet was determined. The supernatant was further analyzed using HPLC to identify unchanged D-cis-FPro and possible metabolites as previously described for L-cis-FPro (Hamacher, 1999).
D-cis-FPro PET in Humans
In a pilot study, two patients were investigated with PET and D-cis-FPro. The first patient (woman, aged 59) had previously been operated for a breast carcinoma. The MRI of the brain did not show signs of cerebral metastases. The second patient was suffering from an oligodendroglioma grade II. A recent MRI of the brain (T1-weighted images before and after Gd-DTPA application and T2-weighted images) was available. In the latter patient also a PET study using O-(2-[18F]fluoroethyl)-L-tyrosine (FET) was available, which demonstrated increased tracer uptake in the tumor (Pauleit et al, 2004). Both patients gave written informed consent for participation in the study. The subjects remained fasted for 12 h before the PET studies. After intravenous injection of 370 MBq D-cis-Fpro, dynamic PET scans of the brain were acquired. Samples of arterialized venous blood were taken every 15 to 30 secs at the beginning with increasing time intervals up to 120 mins. Aliquots of whole blood and plasma were measured in a well counter. To evaluate tracer stability, 2 to 5 μl probes of the blood plasma at different times after injection were spotted on a silica gel for thin-layer chromatography (TLC). The TLC was developed using a mixture of 70% phosphate buffer (pH 5.7) and 30% ethanol. The radioactive components were determined by imaging on a Hewlett-Packard Instant Imager. The identity of components was established by comparison to the reference value of intact D-cis-FPro.
The PET studies were performed using a Siemens ECAT Exact HR+ (optimum FWHM 4.5 mm, 15 cm transaxial field of view). Transmission scans were performed with 68Ge/68Ga rotating line sources and used for measured attenuation correction. The duration of the PET scans was 50 mins. After correction for random and scattered coincidences, dead-time and decay, data were FORE rebinned and reconstructed by filtered back projection.
A ROI was manually placed on the frontal cortex at the level of the basal ganglia. The SUV was calculated by dividing the decay corrected mean radioactivity of the measured ROI (kBq/mL) and the radioactivity injected per gram body weight. Standardized uptake values were corrected for radioactivity from the vascular space, assuming a cerebral blood volume of 3.8% (Ito et al, 2004).
Data were compared with those of a former study investigating the cerebral uptake of L-cis-FPro in patients with brain tumors after correction for radioactivity from the cerebral blood volume (Langen et al, 2004).
Kinetic Analysis
To get additional information on the metabolic behaviour of D-cis-FPro in the human brain, the linearization procedure of Patlak et al (1983) was applied to tissue and plasma time–activity data of the two human subjects investigated with D-cis-FPro and PET. If the Patlak plot becomes asymptotically linear, the rate constant of net tracer influx Ki can be estimated. The rate constant of net tracer influx Ki, which includes tracer transport and possible intracellular binding, is given by the slope of the linear part of this plot, the volume of distribution (Vd) by the y-intercept. The plasma input function was corrected for intact tracer and brain tissue data for radioactivity from the vascular space assuming a cerebral blood volume of 3.8% (Ito et al, 2004).
For comparison, the kinetic analysis was also applied to the tissue and plasma time–activity data of a patient investigated with L-cis-FPro within a former study (Langen et al, 2004). This patient (no. 2 in that study) had been operated for an anaplastic oligoastrocytoma WHO grade III, but was in tumor remission at the time of the PET study. The time–activity data were also corrected for radioactivity from the cerebral blood volume as mentioned above.
Statistical Analysis
Values are expressed as mean±standard deviation. Statistical methods used were t-test or Mann–Whitney rank sum test for group comparisons. Probability values less than 0.05 were considered significant.
Results
Autoradiography
D-cis-FPro and D-Pro exhibited significantly higher uptake in the brain of rats than the corresponding L-isomers. The SUV of D-cis-FPro in the cerebral cortex of the rat 2 h after intravenous injection was 3.05±1.18 (n=9) versus 0.06±0.01 (n=4) for L-cis-FPro (P<0.01) (Table 1). For D-Pro, the SUV was 1.29±0.27 (n=4) versus 0.30±0.14 (n=9) for L-Pro (P<0.001). An example of the differences of cerebral accumulation between D-cis-FPro and D-Pro as demonstrated by dual tracer autoradiography in different coronal sections of the rat brain is shown in Figures 1A and 1C. Uptake of both tracers in the gray matter was almost homogeneous, but D-Pro showed increased uptake in the pyramidal cell layer of the hippocampus corresponding to an area with a high density of nucleoli. Furthermore, D-Pro exhibited a high accumulation in the chorioid plexus, which corresponds to an area with a high protein synthesis rate. In contrast, D-cis-FPro did not accumulate in the pyramidal cell layer of the hippocampus and in the chorioid plexus. This finding indicates that D-cis-FPro is not incorporated into proteins.
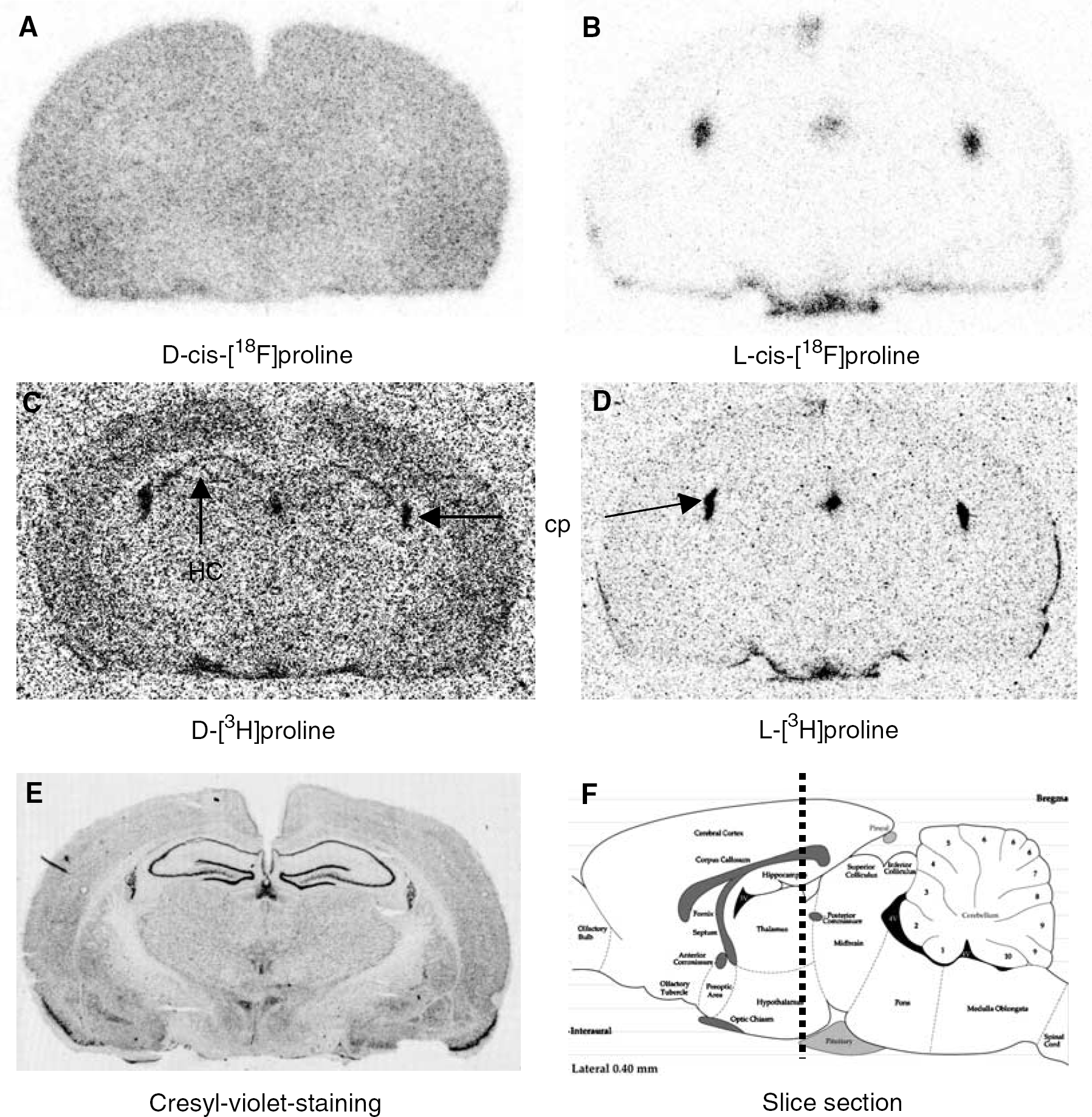
Coronal autoradiograms of the rat brain 2 h after injection of labelled prolines (
Tracer uptake in the frontal cortex of rats (SUV=standardized uptake value, tracer uptake normalized to injected dose and body weight)
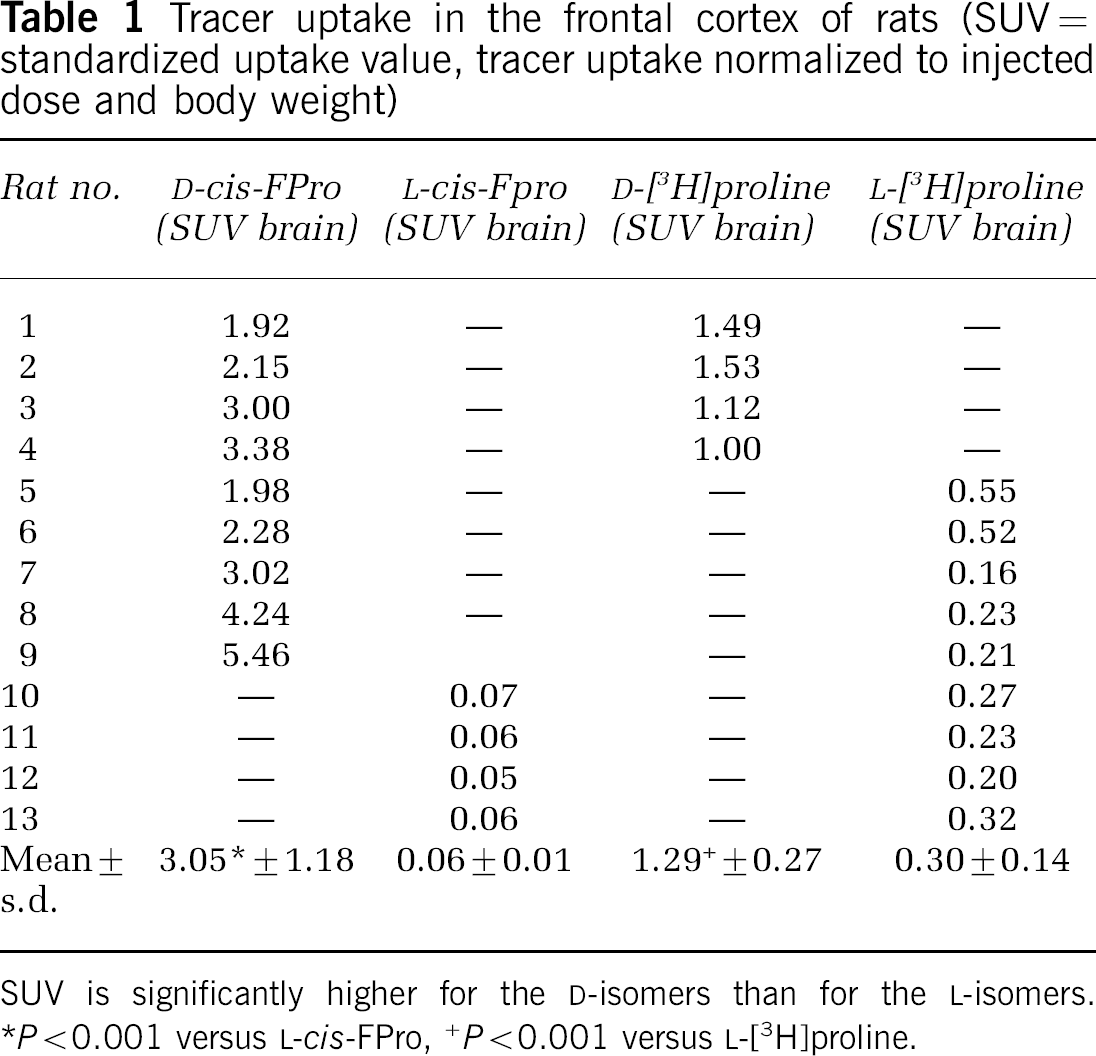
SUV is significantly higher for the D-isomers than for the L-isomers.
P<0.001 versus L-cis-FPro,
P<0.001 versus L-[3H]proline.
Corresponding autoradiograms of L-cis-FPro and L-FPro are shown in Figures 1B and 1D. Uptake of both tracers in normal brain tissue is negligible 2 h after intravenous injection indicating that the permeability of the BBB for both tracers is extremely low. The plexus chorioideus exhibits increased uptake for both tracers.
Analysis of Tracer in Brain
At 1 h after injection of D-cis-FPro, less than 20% of radioactivity was found in the pellet of the rat brain homogenate (n=4). Further separation of the pellet by gel electrophoresis could not identify significant amounts of tracer in the high molecular fraction, that is, no incorporation into protein. HPLC analysis of the supernatant demonstrated that significant amounts of the radioactivity in the rat brain consists of L-trans-FPro, indicating that D-cis-FPro is a substrate of an intracerebral racemase (Figure 2). No further metabolites could be identified.
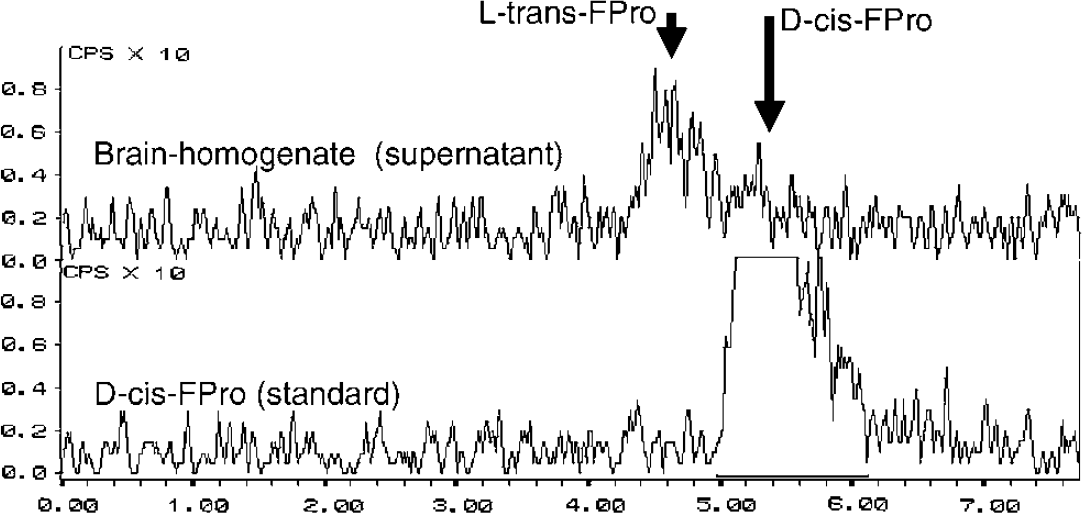
Example of a HPLC analysis of the supernatant of a rat brain homogenate (supernatant) 1 h after injection of D-cis-FPro. The lower curve shows the peak of the D-cis-FPro standard at 5.3 mins. The upper curve demonstrates that significant amounts of the radioactivity in the rat brain consist of L-trans-Fpro, indicating that D-cis-FPro is a substrate of an intracerebral racemase.
D-cis-FPro PET in Humans
The pet studies yielded a SUV of D-cis-FPro in the cerebral cortex of 0.67 in the first patient and 0.45 in the other (40 to 50 mins postinjection), while in a previous study we observed a considerably lower brain uptake for L-cis-FPro (SUV 0.12±0.03, n=7) (Langen et al, 2004). The PET scan of the first patient is shown in Figure 3. Uptake of D-cis-FPro in the normal brain tissue is clearly visible. The tracer concentration in the brain increases continuously from 5 to 50 mins after injection (Figure 3C). The Patlak Plot (Figure 3D) shows a linear slope indicating trapping of the tracer with rate constant of Ki=0.005 min−1 for unidirectional flow. An example of a PET study with L-cis-FPro is shown in Figure 4. This case is taken from a previous study (Langen et al, 2004), which investigated the cerebral accumulation of L-cis-FPro in human brain tumors. The measured tracer concentration in the brain is in the range of background (Figure 4A) and shows no increase up to 50 mins after injection (Figure 4C). Correspondingly, the rate constant for unidirectional flow (D) is by a factor 4 to 5 lower (Ki=0.001 min−1) than for D-cis-FPro. Figure 5 depicts the PET scan of patient 2, who suffered from an oligodendroglioma grade II. There is only minor uptake of D-cis-FPro in the tumor (Figure 5A), while in a corresponding PET study using FET (Figure 5B) the tumor exhibits increased uptake. No sign of BBB disruption within the tumor was found in the corresponding MRI study after injection of the paramagnetic contrast medium Gd-DTPA (Figure 5C). The Patlak analysis of D-cis-FPro kinetics in the frontal cortex and plasma showed a linear slope with a rate constant for unidirectional flow of Ki=0.004 min−1 (data not shown).
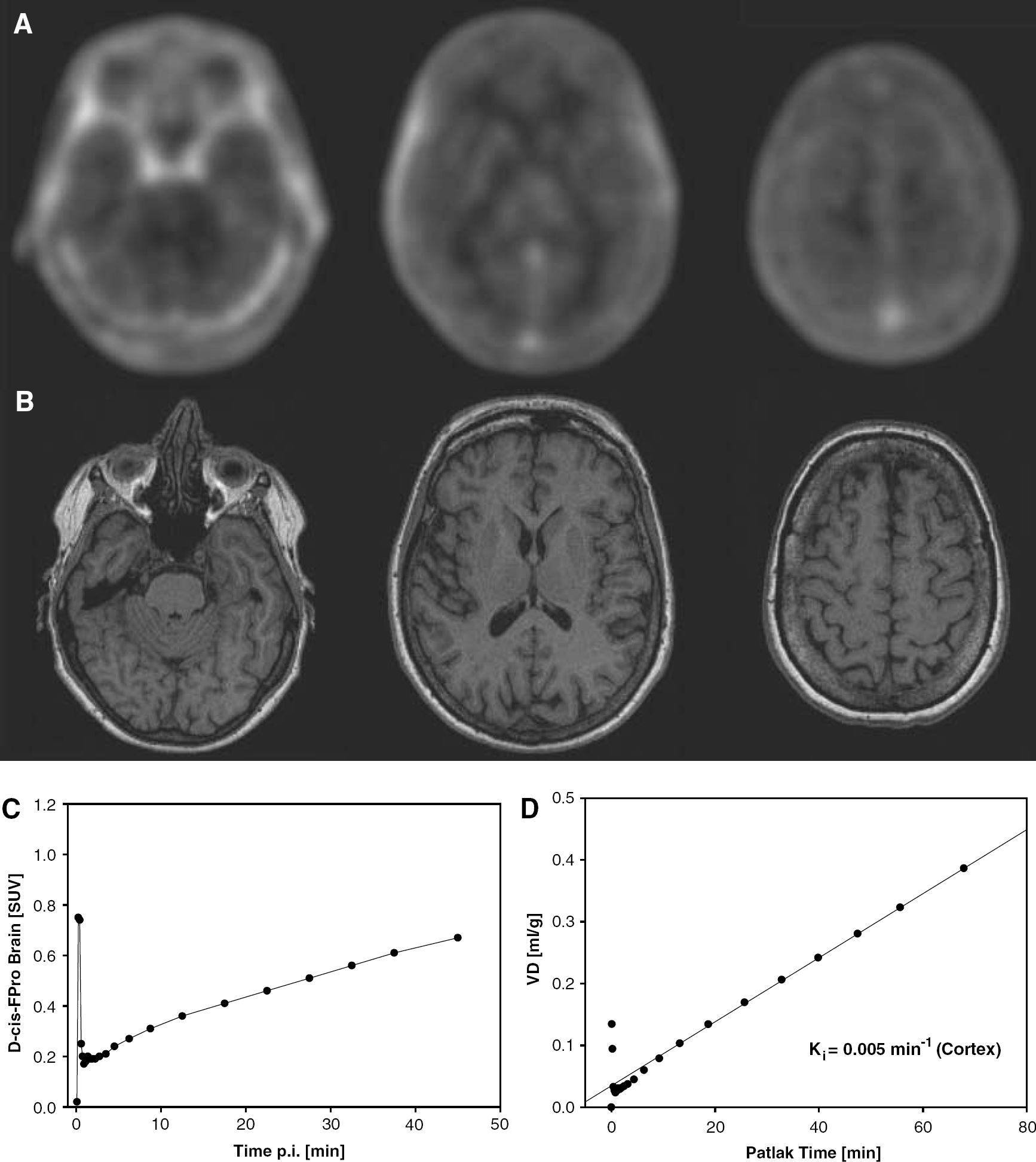
PET scans of patient no. 1 after injection of D-cis-FPro (40 to 50-mins p.i.) (
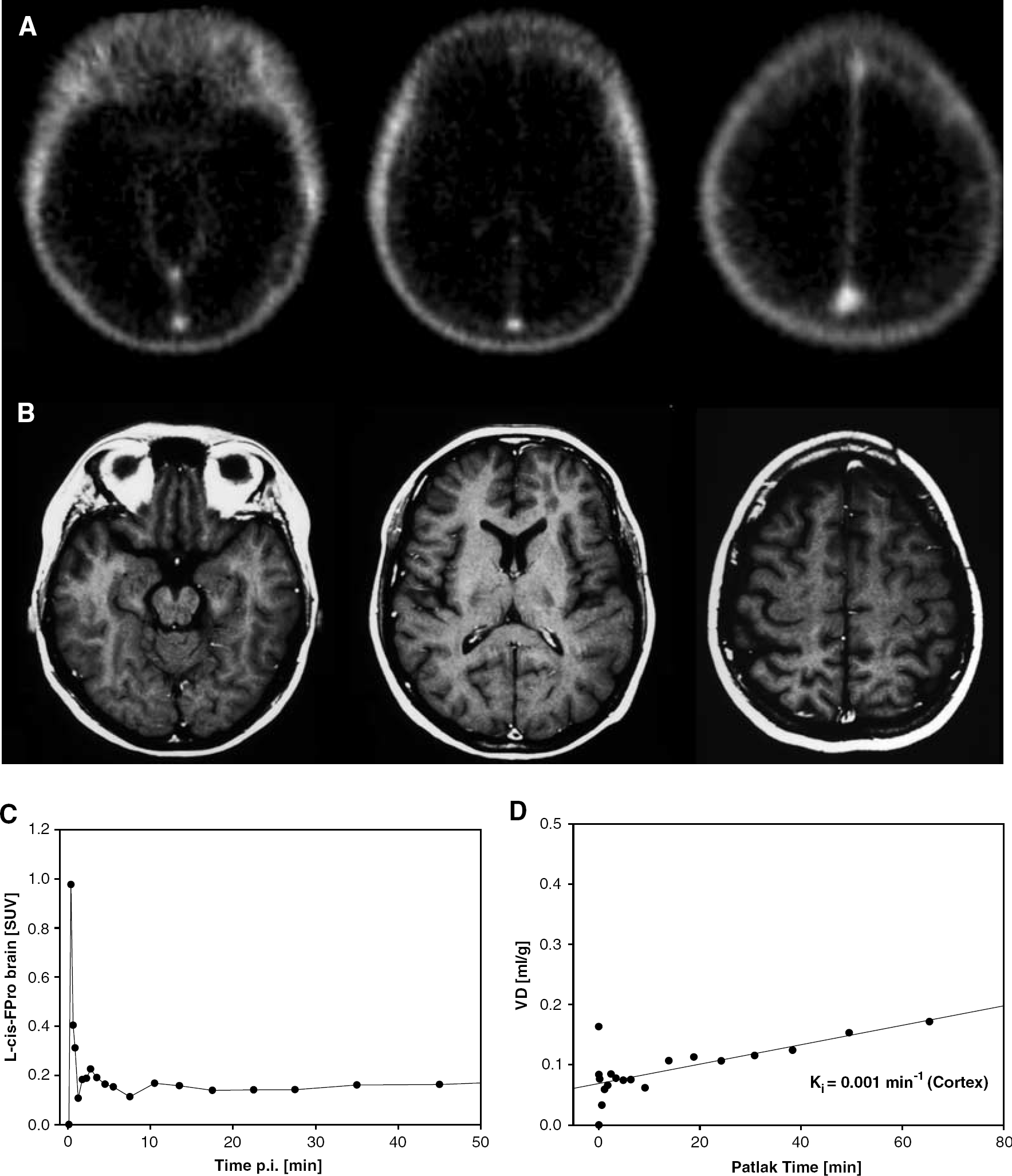
PET scans of a patient after injection of L-cis-FPro (40 to 50-min p.i.) (Langen et al, 2004) (
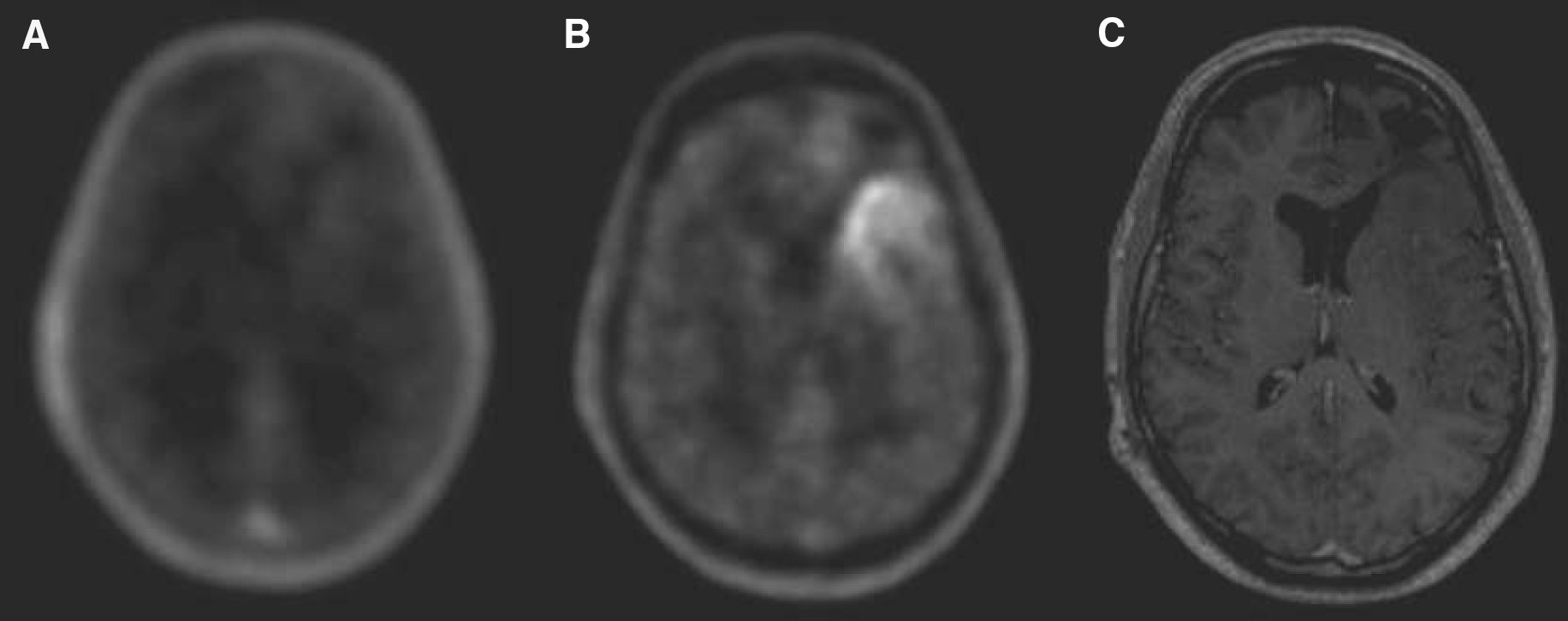
Brain scans of patient 2 suffering from an oligodendroglioma WHO grade II. (
Tracer Stability
The percentage of intact D-cis-FPro of total radioactivity in the plasma of the two patients as evaluated by TLC was found to be 83%/87% at 5 mins p.i., 74%/83% at 20 mins, 68%/58% at 60 mins and 77%/59% at 120 mins, indicating some metabolic degradation of the tracer. For the Patlak analysis, the plasma input curve of D-cis-FPro studies was corrected for intact tracer. L-cis-FPro data were not corrected, since the tracer has been shown to be metabolically stable within the time of measurement (Wester et al, 1999a).
Discussion
The nutritional value and possible toxic effects of D-amino acids have been investigated and discussed for about half a century. L-amino-acid residues in food proteins are subject to racemization to D-isomers under the influence of food-processing conditions, especially heat and alkaline pH. As a result of food processing, D-amino acids are continuously consumed by humans. In addition, D-amino acids are synthesized in vivo by microorganisms by transformation of L-isomers, which are enzymatically catalyzed by amino-acid oxidases, transaminases and racemases (Friedman, 1999). The concentration of D-proline in the human plasma is low and less than 1% of the concentration of L-proline, which ranges from 40 to 150 μmol/L plasma (Nagata et al, 1992; Bruckner and Hausch, 1993). D-proline is partly renally excreted in its unmetabolized form and effectively oxidized by D-amino-acid oxidase to α-keto-δ-amino-valeric acid, which is further metabolized (Schieber et al, 1996).
Data on the toxicity of D-proline are controversial. While Kampel et al (1990) observed fibrosis and necrosis of liver cells in rats after feeding of D-Pro (50 mg/kg of body weight/day), no evidence for subacute toxicity was found in another study under similar conditions (Schieber et al, 1997).
The penetration of L-Pro through the BBB has shown to be extremely low. Both Oldendorf (1971) and Sershen and Lajtha (1976) reported little or no L-Pro uptake into the brain when the amino acid is injected into the rat carotid artery. Until today, it is believed that extracellular L-proline in the brain most likely originates from some combination of local biosynthesis and diffusion from the plasma (Cohen and Nadler, 1997).
In the early 1970s, Garweg and Dahnke reported an intensive labelling of the cerebral cortex of mice after intraperitoneal injection of [3H]D-Pro, while after injection of [3H]L-Pro uptake in the cerebral cortex was negligible (Garweg and Dahnke, 1972). High accumulation was found in the cerebral cortex and in the pyramidal cells of the hippocampus. Further experiments showed that all of the protein bound radioactivity after injection of D-Pro was owing to L-Pro. Stimulated by these observations, the authors compared the uptake of 2-[3H]D-proline and 2,3-[3H]D-proline. The first compound, 2-[3H]D-proline, loses its 3H-label when racemized to L-proline. Indeed, no uptake of radioactivity was observed in the mouse brain. The second, 2,3-[3H]D-proline, which keeps the 3H-label in the 3-position after racemization to 3-[3H]L-proline, exhibited high uptake in the cerebral cortex and in the pyramidal cells of the hippocampus. The authors postulated the existence of a proline racemase in the cerebral cortex of the mouse converting D-proline to L-proline before utilization in protein synthesis.
The observations of Garweg and Dahnke (1972) dating more than 30 years ago have not been taken into consideration by the scientific literature, although the search for L-proline transport systems in the brain and the possible role of L-proline in the modulation of excitatory synaptic transmission has attracted major interest in recent years.
The results of our autoradiographic study comparing the cerebral uptake of L-Pro and D-Pro in rats confirm the results of Garweg and Dahnke in mice that the D-isomer of proline is preferably taken up into the brain and consequently preferably transported across the BBB in contrast to its L-isomer (Garweg and Dahnke, 1972, 1973, 1974). Our data with the D- and L-isomers of the PET tracer cis-FPro suggest that the same holds true for the human brain. To the best of our knowledge, this presents the first report on the preferred transport of a D-amino acid in comparison to its L-isomer across the human BBB.
Analysis of the patients' plasma indicated some metabolic degradation of D-cis-Fpro, but the analysis of the brain homogenates of rats revealed that all of the radioactivity in the brain was owing to free fluoroproline, indicating that the tracer signal in the human brain is not caused by metabolites. The HPLC analysis of the brain homogenate, however, revealed a significant part of the radioactivity in the form of L-trans-FPro, indicating that D-cis-FPro is accepted by a racemase in the human brain and converted to the L-isomer similar to its natural parent. The pattern of cerebral radioactivity distribution after injection of D-cis-FPro in rats, however, was different from that after injection of D-Pro. A labelling of the pyramidal cell layer of the hippocampus was not observed with D-cis-FPro, indicating that D-cis-FPro or its racemate L-trans-FPro is not incorporated into cerebral proteins. This finding was confirmed by gel-electrophoretic analysis of proteins.
In the patient studies, tracer uptake increased linearly from 5 to 50 mins after injection of D-cis-Fpro, indicating trapping of the tracer (Figure 3). This observation was confirmed by the Patlak analysis, which yielded a linear slope with a rate constant of unidirectional flow of Ki=0.005 min−1 for patient one and Ki=0.004 min−1 for patient two. A dynamic study in a patient after injection of L-cis-FPro taken from a previous study (Langen et al, 2004) showed no relevant uptake in the normal brain tissue and the slope of corresponding Patlak analysis was close to zero. It has to be considered that the data for tracer influx are directly influenced by competing substrates in the plasma. The observed differences between the tracers, however, should not be influenced by this factor, provided that the enantiomers share a common carrier system and transport is inhibited by the same substrates. Another interesting aspect was provided by the comparative study with FET in a patient with an oligodendroglioma. While the tumor exhibited increased FET uptake, the accumulation of D-cis-FPro in the tumor was negligible. The tyrosine derivative FET is transported via system L similar to methionine and other large neutral amino acids, and exhibits increased uptake in gliomas (Wester et al, 1999b, 2000; Weber et al, 2000; Langen et al, 2003; Pauleit et al, 2004). The low uptake of D-cis-FPro in this tumor indicates that system L is not involved in the uptake process of D-cis-FPro at the BBB.
The nature of the amino-acid carriers at the BBB which transport the D-isomer of proline remains to be elucidated. The transport of L-proline in the brain has attracted considerable interest. More than 30 years ago, it was discovered that mammalian synaptosomes express a high-affinity, Na+-dependent L-proline uptake activity similar to the synaptosomal uptake activities identified for recognized neurotransmitters, suggesting that L-proline might be a neurotransmitter or neuromodulator in the mammalian central nervous system (Bennett et al, 1972; Peterson and Raghupathy, 1972; Balcar et al, 1976; Hauptmann et al, 1983; Nadler, 1987). In 1992, cDNAs were isolated from rat and human brain that code for a high-affinity Na+-dependent transporter designated PROT (Fremeau et al, 1992, 1996; Shafqat et al, 1995; Renick et al, 1999; Crump et al, 1999). The physiological properties clearly distinguished this carrier from the other widely expressed mammalian Na+-dependent plasma membrane carriers that transport L-proline, that is, systems A and ASC. The isoform ATA2 of system A has been shown to be predominantly expressed at the BBB, acting as brain-to-blood efflux transporter for L-proline (Takanaga et al, 2002). The brain-specific expression of PROT in subpopulations of putative glutamatergic neurons in rat brain supported the consideration of a specialized role for this transporter and its presumed natural substrate L-proline in the modulation of excitatory synaptic transmission. Within the rat hippocampal formation, high-affinity uptake of L-proline is confined to a subset of glutamatergic nerve terminals. The Schaffer collateral–commissural projections take up L-Pro in vitro, whereas the mossy fibers and medial perforant path do not (Nadler et al, 1992). An L-proline-induced potentiation of NMDA-receptor activation in the Schaffer collateral–commissural synaptic transmission to CA1 pyramidal cells of the rat hippocampus at a concentration typical of normal human cerebrospinal fluid has been shown (Cohen and Nadler, 1997).
Recently, two novel mammalian amino-acid cotransporters have been characterized by expression in Xenopus oocytes which induce a pH-dependent electrogenic transport actvity for the small amino acids glycine, alanine and proline (Boll et al, 2002). The mRNA of the transporter designated mPAT1 was highly expressed in the brain. The transporter was found to be not very enantioselective and in oocytes expressing mPAT1 D-proline evoked currents which were almost as high as those of glycine. PAT2 was found to be expressed in neurons positive for the NMDA subtype of glutamate receptor subunit NR1 (Rubio-Aliaga et al, 2004). A preferred selective transport of D-proline, however, has not yet been described, suggesting that the amino-acid transporter at the BBB which preferably transports the D-isomers might still be unknown.
The experiments of Garweg and Dahnke (1972, 1973, 1974) as well as our results in rats and humans suggest that D-Pro might be a source of L-Pro in the brain. The biological meaning of the preferred uptake of the D-isomer of proline remains to be elucidated. Assuming an important role of L-Pro in excitatory neurotransmission, a free diffusion of this amino acid through the BBB would be hazardous to the brain similar to glycine or glutamate. The free diffusion of D-proline, which is much lower concentrated in the blood than L-proline, could guarantee a constant but low supply of a substrate from which L-proline is generated in brain tissue by racemases. This discovery opens new aspects concerning the physiological role of D-amino acids in the central nervous system. Disturbances of glutamatergic excitatory neurotransmission are involved in many brain diseases such as ischemia and trauma (Arundine and Tymianski, 2004), multiple sclerosis (Groom et al, 2003), schizophrenia (Harrison et al, 2003), Alzheimer's disease (Doraiswamy, 2003; Hynd et al, 2004) and cerebral gliomas (Takano et al, 2001).
It is tempting to speculate that the intracerebral concentration of L-proline and its precursor D-proline might be affected in some of these brain diseases and D-cis-FPro might be an interesting new tracer for PET imaging.
Footnotes
Acknowledgements
We thank Mr Markus Cremer and Mr Norbert Hartwigsen for assistance in animal experiments, Mrs E Theelen, Mrs S Schaden and Mr Markus Lang for assistance in PET studies, Mrs Silke Grafmüller, Mrs Bettina Palm, Mrs Erika Wabbals and Mr Sascha Rehbein for assistance in the radiosynthesis of D-cis-FPro.