
Abstract
The first molecular insights into how prodrugs modified with ethanolamine-related structures target the brain were generated using an
INTRODUCTION
The blood–brain barrier (BBB) poses the greatest challenge for therapeutic agents meant to act in the brain.1,2 Most drugs cannot enter the brain in pharmacologically relevant amounts, leading investigators to abandon efforts to target them specifically to this organ. 3 One strategy to improve poor drug uptake in the brain is to administer compounds as prodrugs that can be shuttled by transporters specifically expressed at the BBB. 4 Using this approach, researchers have succeeded in designing drug-carrier conjugates that can specifically interact with glucose transporters (GluT1), 5 amino acid transporters (LAT1), 6 and choline transporters, 7 thereby targeting the drugs to the brain. For cationic drugs, however, the transporters that may move them across BBB remain unknown.8,9 Elucidating the mechanism of cationic drugs transport across the barrier may help guide the development of novel brain-targeting prodrugs.
Six transporters that can shuttle cationic drugs across the cell membrane have already been identified in the kidney and liver: OCT1 (SLC22A1), OCT2 (SLC22A2), OCT3 (SLC22A3), OCTN1 (SLC22A4), OCTN2 (SLC22A5), and MATE1 (SLC47A1).10,11 Whether these transporters are expressed and functional at the BBB remains unknown. Several studies suggest that an antiporter system capable of transporting of organic cations may have a useful role in carrying cationic drugs passed the barrier. Yamazaki
Evidence that these putative pyrilamine cationic transporters may participate in an antiporter system came when the transport of secondary or tertiary amine drugs (e.g., nicotine, 15 clonidine, 16 and tramadol 17 ), which shows similar characteristics as pyrilamine transport, was found to be driven by an adverse proton gradient. Pyrilamine inhibited the tranport process, suggesting the existence of a pyrilamine-senstive H+/OC-antiporter. This antiporter is the best current candidate for a transporter capable of moving cationic drugs across the BBB. Little is known about how the transporter moves drugs across the barrier, whether it is the only mechanism that can do so and what are the chemical or steric requirements for effective drug transport.
Previous work from our laboratory has shown that prodrugs modified with ethanolamine-related structures (prodrugs I, II, III, and IV, Figure 1) can target the brain effectively.
18
They showed brain-to-plasma concentration ratios that were 9.31- to 17.0-fold higher than that of the drug dexibuprofen on which they were based. Subsequently, we found that
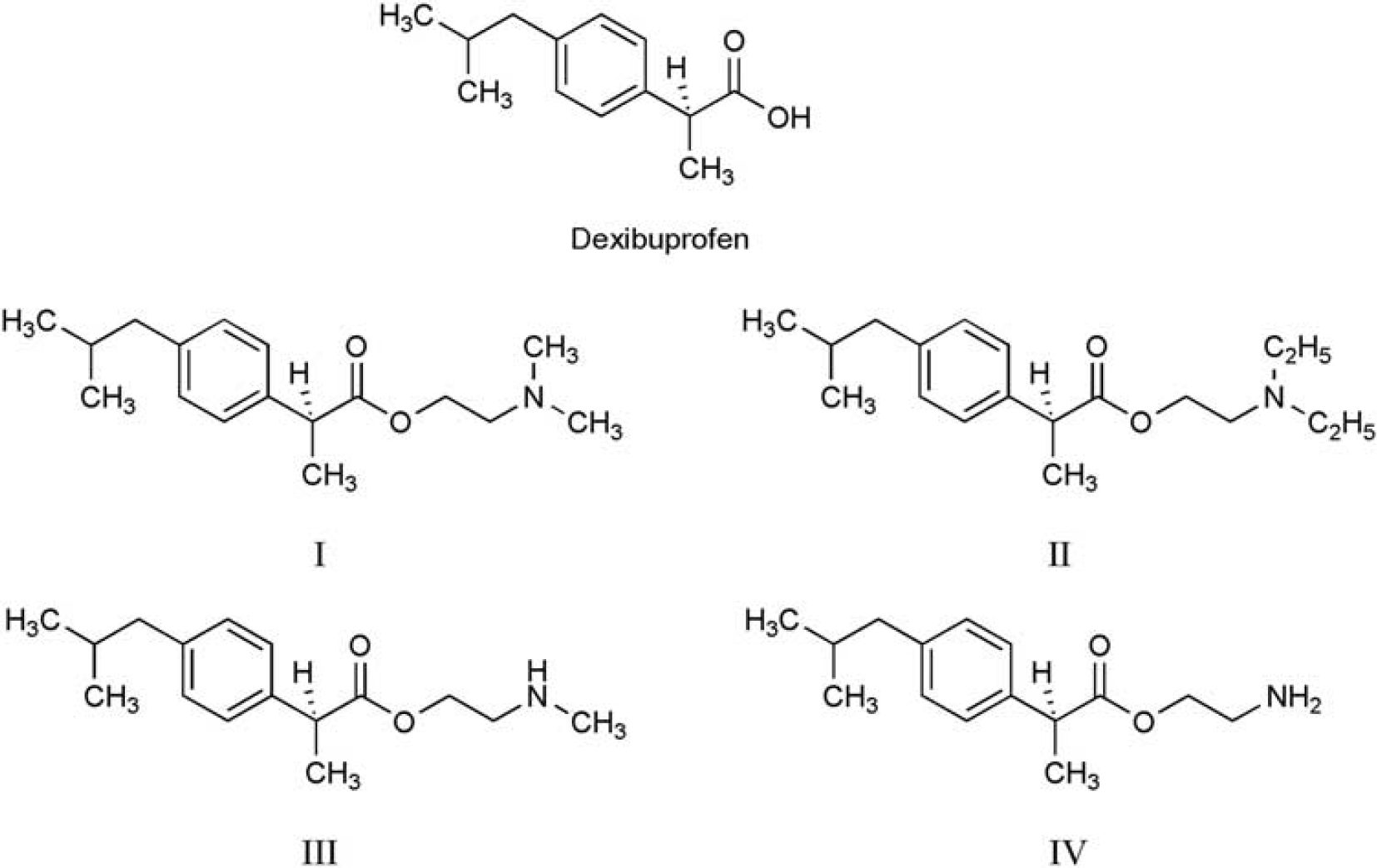
Chemical structures of dexibuprofen and prodrugs I, II, III, and IV.
MATERIALS AND METHODS
Materials
Dexibuprofen (99.8% purity) was purchased from the Fourth Pharmaceutical Company of Suzhou (Suzhou, China). Prodrugs I, II, III, and IV were synthesized and identified according to our previous work.
18
1-(4-(Trimethylamino) phenyl)-6-phenyl-1,3,5-hexatriene (TMA-DPH), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), trypsin,
Animals and Cell Culture
All animal experiments were approved by the Institutional Animal Care and Ethic Committee of Sichuan University (Approved No. SYXK2013-185) and performed in accordance with the NIH Guide for the Care and Use of Laboratory Animals. All animals were fed on a light and dark cycle and allowed free access to standard chow and water. Temperature and relative humidity were kept at 25° and 50%, respectively. After experiment, rats were killed by neck dislocation under anesthesia, and all efforts were made to minimize suffering. The report was written in compliance with ARRIVE guidelines. 22
L929 cells (passages: 3 to 20) that were purchased from the American Type Culture Collection (Rockville, MD, USA) and cultured in DMEM with high glucose (Hyclone, Logan, UT, USA) supplemented with 10% FBS (Hyclone), 100IU/mL penicillin, and 100 μg/mL streptomycin. Cells were maintained at 37° in a humidified atmosphere containing 5% CO2, and the culture medium was changed every other day.
In Vitro Co-Culture Model of the Blood–Brain Barrier
Blood to brain by capillary endothelial cells and astrocytes (ACs) were isolated from rat brain cortex and used to establish primary cocultures as described in ‘Supplementary Information’. Co-cultures were set up using the ‘contact through feet’ approach, in which endothelial cells are cultured on the upper face of transwell inserts and ACs are seeded on the lower face. In this way, ACs can spread their processes through the membrane pores to contact endothelial cells. Cell culture inserts with a microporous polyethylene terephtalate membrane (1 μm diameter, Millipore, Darmstadt, Germany) were turned upside down, and ACs were seeded on top at a density of 10 4 cells per insert. After 4 hours, the insert was turned over (right side-up) and placed in 24-well plates. Astrocyte medium was added and replaced every other day. After 5 days, BCECs were seeded onto the upper side of the insert at a concentration of 5 × 10 4 cells per insert, BCEC culture medium was added and replaced every other day. After an additional 5 days, transendothelial electrical resisitance (TEER) of the modeled BBB was measured using a TEER instrument (Millipore). When TEER reached 250 Ω cm2, the BBB model was considered ready for experiments. The morphologic characteristics and barrier integrity were analyzed by transmission electron microscopy (JEM 100CX, JEOL, Tokyo, Japan) and scanning electron microscopy (S-450, Hitachi, Japan).
Prodrug Effects on Blood–Brain Barrier Integrity
Prodrug Effects on Surface Membrane Potential of Blood to Brain by Capillary Endothelial Cells
Confluent BCEC monolayers were washed three times with PBS, trypsinized, and diluted with HEPES buffer to a final concentration of 4×10 6 cells/mL. Then, TMA-DPH, IBU, or prodrugs were added to cell suspensions to final concentrations of 54 μmol/L (TMA-DPH) or 80 μmol/L (DIBU and prodrugs) and incubated for 30 minutes. Surface membrane potential (zeta-potential, Z) was analyzed as described 23 using a Zetasizer Nano ZS (Malvern, UK) equipped with a He-Ne laser (λ = 632.8 nm). Measurements were taken at 25° using disposable zeta cells with platinum gold-coated electrodes. 24
Prodrug Uptake by Blood to Brain by Capillary Endothelial Cells Blood to brain by capillary endothelial cells were seeded in 6-well culture plates (5×10
5
cells/well), incubated for 48 hours and checked under the microscope for confluence and morphology. Then BCECs were washed twice with 1 mL of PBS and preincubated with HBSS buffer for 20 minutes at 37°. Thereafter, buffer containing different concentrations of DIBU or prodrug was added to initiate uptake. Cells were incubated at 37° for a predetermined time period, after which uptake was terminated by washing three times with 1 mL of ice-cold PBS. Cultures were trypsinized and centrifuged at 1,000
Sample containing prodrugs was subjected to hydrolysis. Lysates containing prodrugs (100 μL) were mixed with 20 μL 1N NaOH and allowed to stand for 10 minutes at 25°. Acetonitrile (200 μL) was added and mixtures were vortexed for 5 minutes, then centrifuged at 14,000
This uptake procedure was performed at both 4° and 37° to examine whether passive diffusion is a driving force for prodrug uptake. The procedure was also performed after 30-minute pretreatment with metabolic energy inhibitors (0.1% NaN3 or 25 μmol/L rotenone) to examine the energy requirements of prodrug transport.
To begin to examine whether the mechanisms of prodrug uptake depend on cell line, we incubated cultures of BCECs and L929 cells for 1 hour at 37° with DIBU or prodrug (80 μmol/L). Cells were washed three times with ice-cold PBS and assayed for internalized prodrug as described above.
Prodrug Uptake Mechanisms in Blood to Brain by Capillary Endothelial Cells
To examine the mechanisms of prodrug uptake in the assay described above, cells were preincubated in the presence of different additives. To test the sodium ion dependency of uptake, cells were preincubated in medium containing
The ability of cationic inhibitors to affect prodrug uptake was assessed by pretreating cultures with the inhibitors (including 200 μmol/L of pyrilamine, propranolol, and imipramine, 500 μmol/L astrocytes (ACs) of hemicholinium-3, choline, L-carnitine, TEA, L-lysine, L-arginine, and spermine) for 30 minutes at 37°, then adding prodrug I or II (80 μmol/L) and incubating another 1 hour. Cultures were rinsed three times with ice-cold PBS and processed as described above. Lineweaver-Burk plot describing uptake of prodrugs I and II was generated by performing the uptake assay at concentrations of 20 to 120 μmol/L in the absence or presence of 200 μmol/L pyrilamine.
Prodrug Transendothelial Transport in an In Vitro Blood–Brain Barrier Model
Transendothelial transport was studied as described.
25
Briefly, the medium in the lower chamber of the BBB model was replaced with 1 mL HBSS, and 0.2 mL HBSS containing DIBU or prodrug (80 μM) was added to the upper chamber. At 15, 30, 60, 90, and 120 minutes, 100 μL of HBSS was collected from the lower chamber, and the same volume of fresh HBSS was added to maintain constant volume. The 100-μL sample was diluted with acetonitrile for HPLC analysis. Transendothelial drug permeability was measured in terms of the accumulated cleared volume (μL): accumulated cleared volume (μL) = drug accumulated in the lower chamber/drug initially added to the upper chamber. An apparent permeability coefficient (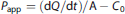
This assay was repeated under different conditions to probe the mechanisms involved. It was performed at 4° to allow comparison with 37° results and thereby assess the role of lipid-mediated simple diffusion. It was performed in the presence of 0.1% NaN3 to assess the energy dependence of the process. To assess the possible role of pyrilamine-sensitive H+/OC antiporter, medium in the upper chamber was supplemented with cationic inhibitors at 37° for 15 minutes before addition of prodrugs I and II.
In situ Rat Brain Perfusion with Prodrugs
Brain perfusion was performed as described. 26 Male Sprague-Dawley rats weighing 220 ±20 g (Dashuo Biotechnology, Chengdu, China) were anesthetized using intraperitoneal chloral hydrate (400 mg/kg), and the right carotid artery was perfused with Krebs-Henseleit buffer (118 mmol/L NaCl, 4.7 mmol/L KCI, 25 mmol/L NaHCO3, 1.2 mmol/L KH2PO4, 2.5 mmol/L CaCl2, 1.2 mmol/L MgSO4, 10mmol/L D-glucose, 10mmol/L HEPES, and 20IU/mL heparin, pH 7.4) containing DIBU or prodrug (80 μmol/L). Perfusion was performed for 30 seconds at a flow rate of 4.9mL/min. After perfusing the brain with drug-free Krebs-Henseleit buffer for another 30 seconds, the right brain hemisphere was collected, rinsed with ice-cold saline solution, wiped dry, weighed, and stored at − 20° until analysis.
To examine the role of passive diffusion in prodrug uptake by the brain, brain capillaries were first perfused for 30 seconds with drug-free perfusion medium cooled to 4°. In additional experiments, medium containing prodrug was precooled to 4° before perfusion, or it was supplemented with 0.1% NaN3. To examine the role of putative OC transporters at the BBB, medium containing prodrug I or II (80 μmol/L) was supplemented with 1 mmol/L pyrilamine before perfusion.
Statistical Analysis
Statistical comparisons were performed by one-way analysis of variance (ANOVA) and Student's t-test,
RESULTS
In the first set of experiments, BCEC monolayers were used to mimic the structure and function of the BBB. These monolayers were incubated with DIBU or each of the four prodrugs to examine cytotoxicity, effects on membrane potential, and mechanisms of prodrug uptake. In the second set of experiments, transendothelial transport of prodrugs in BCEC-AC cocultures was analyzed as a model of the BBB. This two-step experimental approach mirrors the overall process of drug uptake by the brain, which is thought to comprise uptake by endotheliocytes, followed by passage across the BBB into the brain parenchyma.
Characterization of the
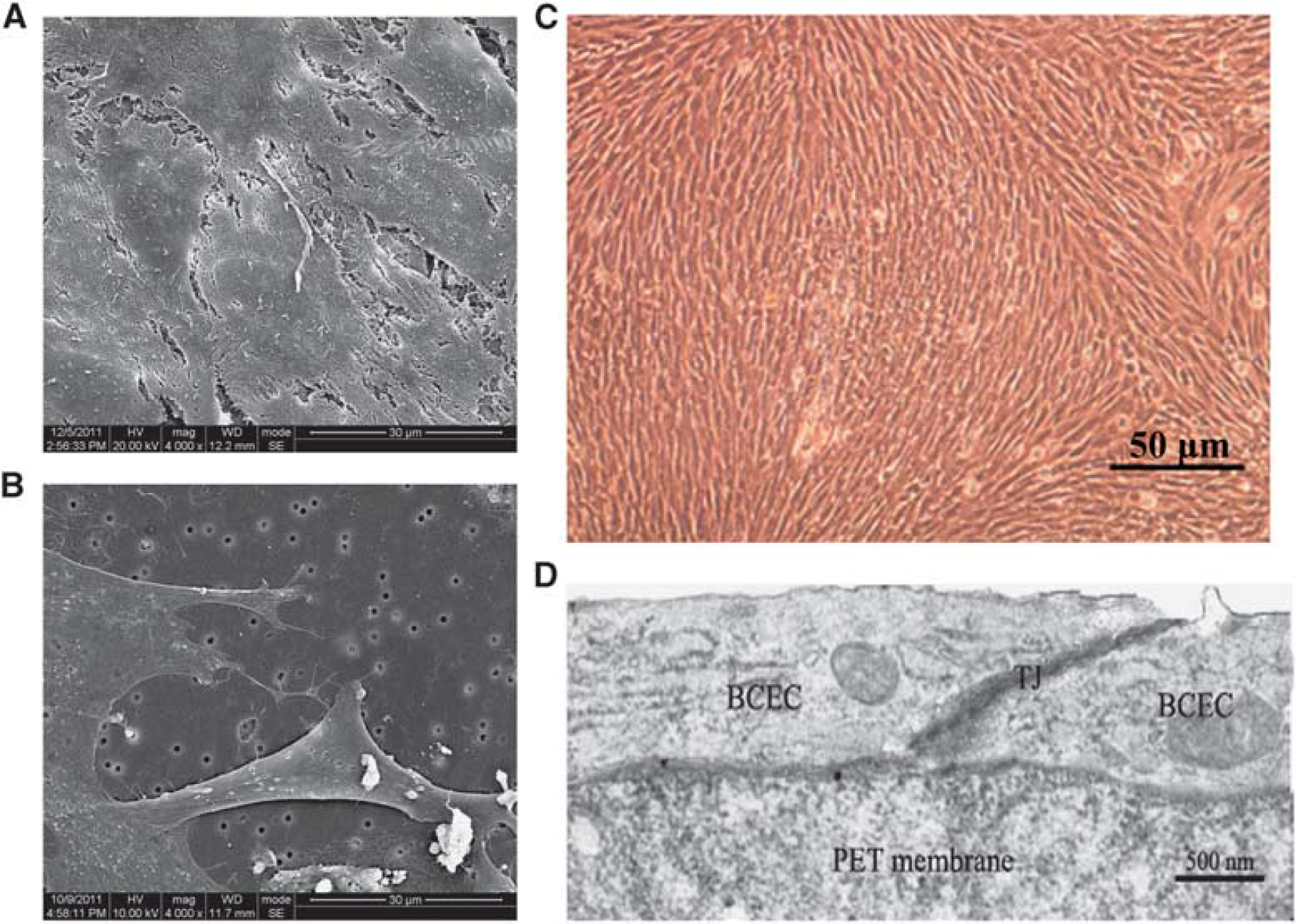
Morphologic analysis of
Prodrug Cytotoxicity in Blood to Brain by Capillary Endothelial Cells
Before beginning any other culture experiments, the MTT assay was used to measure the cytotoxicity of DIBU or prodrugs with ethanolamine-related structures. Even at the highest prodrug concentrations tested, more than 80% of cells remained viable after 4-hour incubation with prodrugs (Figure 3A). This suggests that modifying DIBU with ethanolamine-related structures does not increase its cytotoxicity.
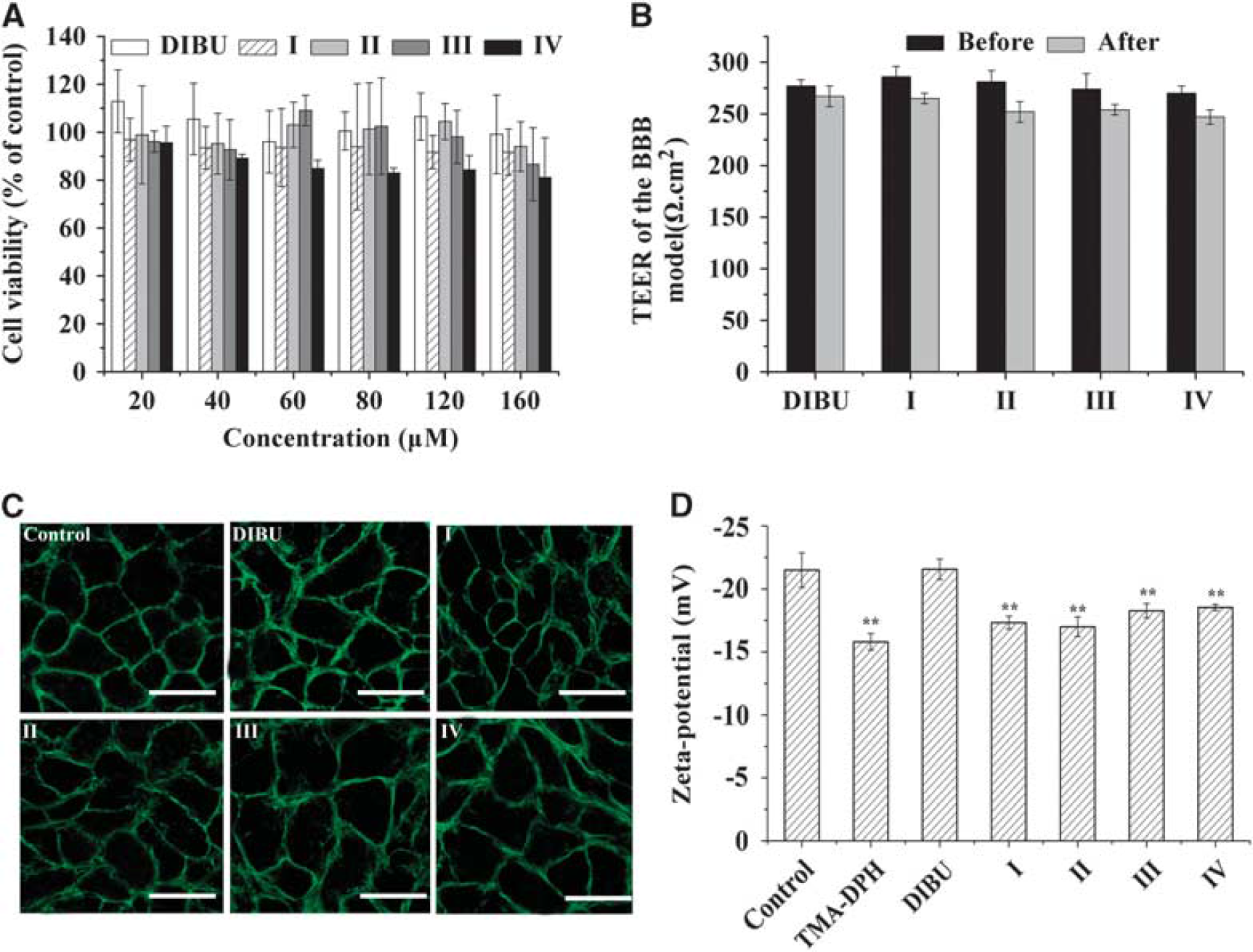
Effects of prodrugs on the tight junctions of the
Transendothelial electrical resisitance of the
Prodrug Effects on the Blood to Brain by Capillary Endothelial Cell Membrane Potential
As reported previously,
22
membrane surface charge (zeta potential) of BCEC monolayers was highly negative, ζ = − 21.50± 1.37 mV (Figure 3D). TMA-DPH, a cationic probe of membrane dynamics in living cells, served as a positive control. This probe intercalates among the hydrophilic head groups of membrane phospholipids and neutralizes the anionic charges of lipids in the outer layer of the cytoplasmic membrane. As expected, TMA-DPH, increased the zeta value (ζ = − 15.80± 0.66 mV), bringing it closer to the uncharged state. While incubating the monolayers with DIBU did not significantly change the zeta potential from the value in the control group (no treatment,
Prodrug Uptake by Blood to Brain by Capillary Endothelial Cells All prodrugs showed significantly greater uptake than DIBU with increasing incubation time and concentrations (Figure 4). Relative uptake under these experimental conditions followed the sequence: I > II > III > IV> DIBU. This is consistent with our previous study of brain targeting by these prodrugs. 18
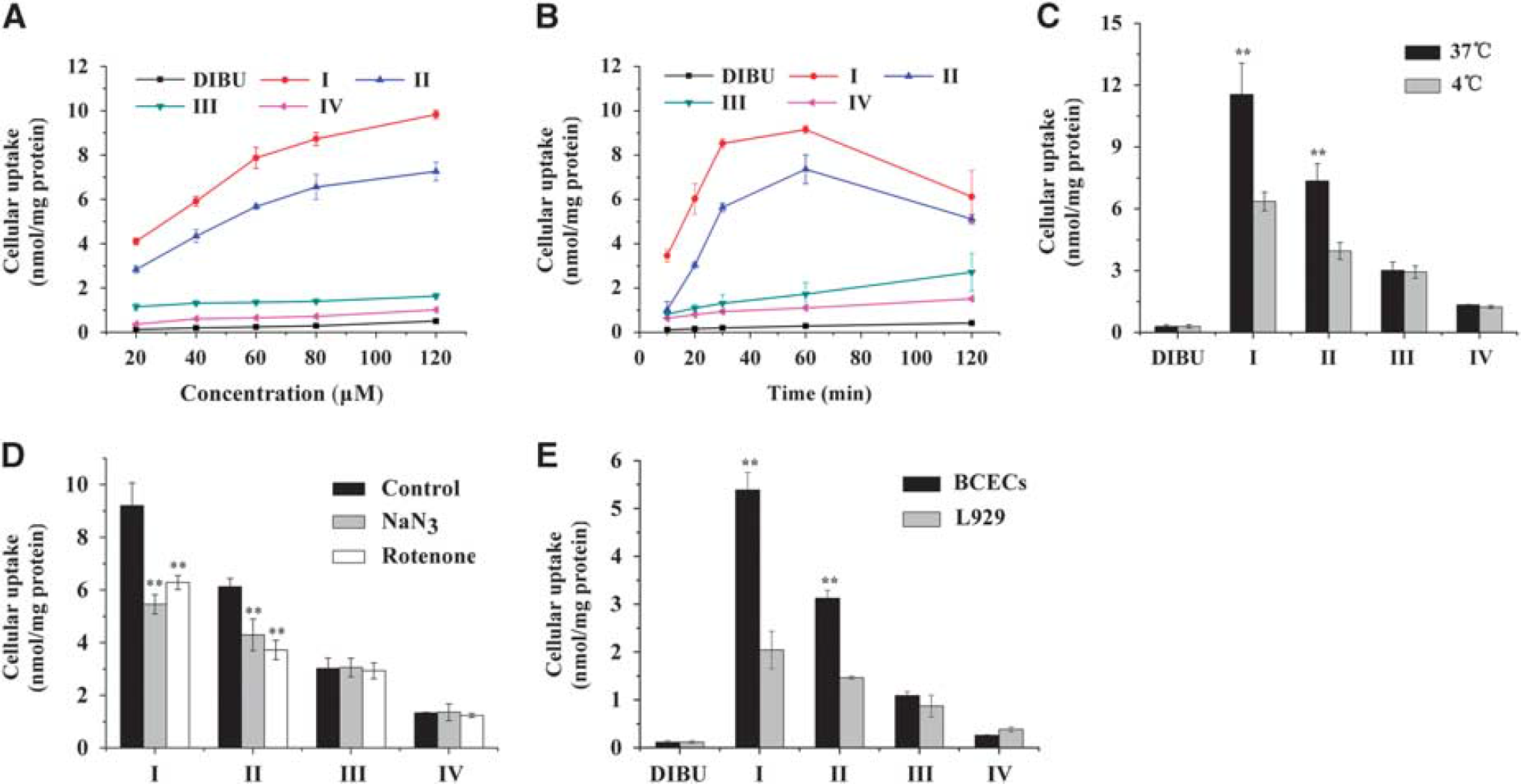
Uptake of dexibuprofen (DIBU) and prodrugs I, II, III, and IV by blood to brain by capillary endothelial cell (BCEC) monolayers under various conditions. (
Uptake characteristics were different for the prodrugs. Uptake of DIBU and of prodrugs III and IV increased linearly with incubation time and concentrations, it did not differ significantly between 37° and 4°, and it was not inhibited by sodium azide or rotenone. These results suggest that DIBU and prodrugs III and IV enter the brain primarily via passive membrane diffusion. Similar results were obtained in BCEC and L929 cultures, suggesting that the uptake mechanism is cell line independent.
Unlike the uptake of these two prodrugs, the uptake of prodrugs I and II by BCECs was time, concentration, temperature, and energy dependent (Figure 4). Uptake was greater at 37° than at 4° for both prodrug I (11.55 ± 1.51 versus 6.36±0.45 nmol/mg) and prodrug II (7.35 ±0.83 versus 3.95 ±0.41 nmol/mg). Uptake of I and II was significantly inhibited by pretreatment with sodium azide and rotenone in BCECs (Figure 4D). These results suggest an active transport process. Consistent with this interpretation, higher doses of prodrug I and II showed saturation behavior (Figure 4B). Prodrug II showed a much lower affinity for the putative transporter system, reflected in a lower maximum uptake (6.36 ±0.42 versus 9.83 ±0.19 nmol/mg). Uptake of prodrugs I and II was much greater in BCECs than in L929 cells (Figure 4E), suggesting that the functional characteristics of the transport system that brings these prodrugs into the brain may differ across cell lines.
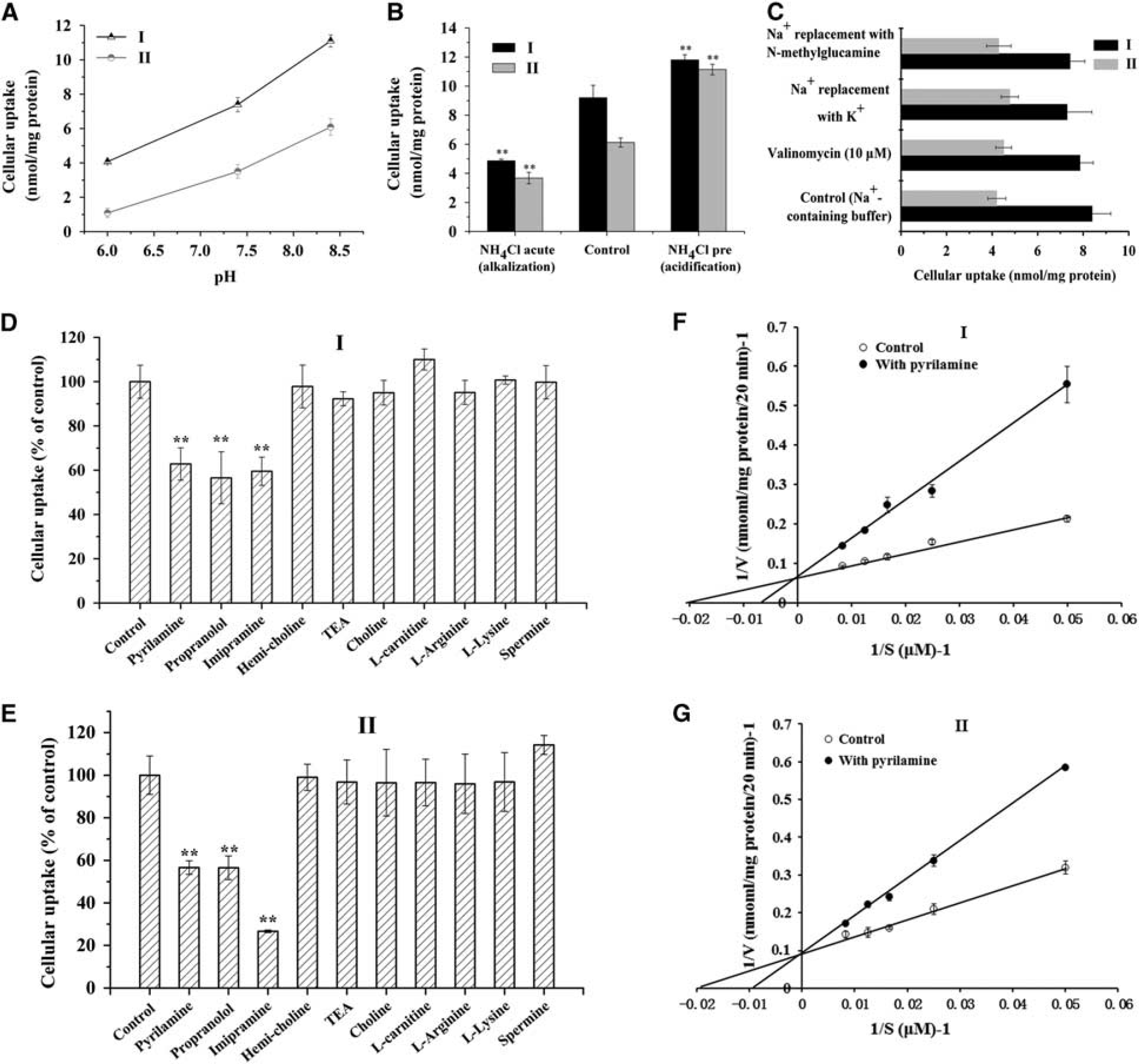
Mechanistic studies of the uptake of prodrugs I and II by blood to brain by capillary endothelial cell (BCEC) monolayers. (
Structures of the reported substrates of the H+/OC antiporter
To examine the effect of intracellular pH on cell uptake, cells were treated with NH4Cl. Acute treatment with NH4Cl, which causes intracellular alkalinization, significantly reduced the uptake of prodrugs I and II to ~ 50% of the control. Pretreatment with NH4Cl, which causes intracellular acidification, significantly stimulated uptake of prodrugs I and II (Figure 5B).
Replacing extracellular Na+ with
To begin to define the molecular characteristics of the putative transporters involved in prodrug uptake in BCECs, the uptake assay was repeated in the presence of potential inhibitors. The cationic drugs such as pyrilamine, propranolol, and imipramine significantly inhibited uptake of I and II (Figures 5D and 6E), but the choline transport inhibitor hemicholinium-3 and the OCTN2-specific substrate L-carnitine did not. Choline, the substrate of the choline transport system, and tetraethylammonium, a classical inhibitor of OCTs, only slightly inhibited uptake. Similarly, alkaline amino acids such as L-lysine, L-arginine, and spermine did not significantly affect uptake.
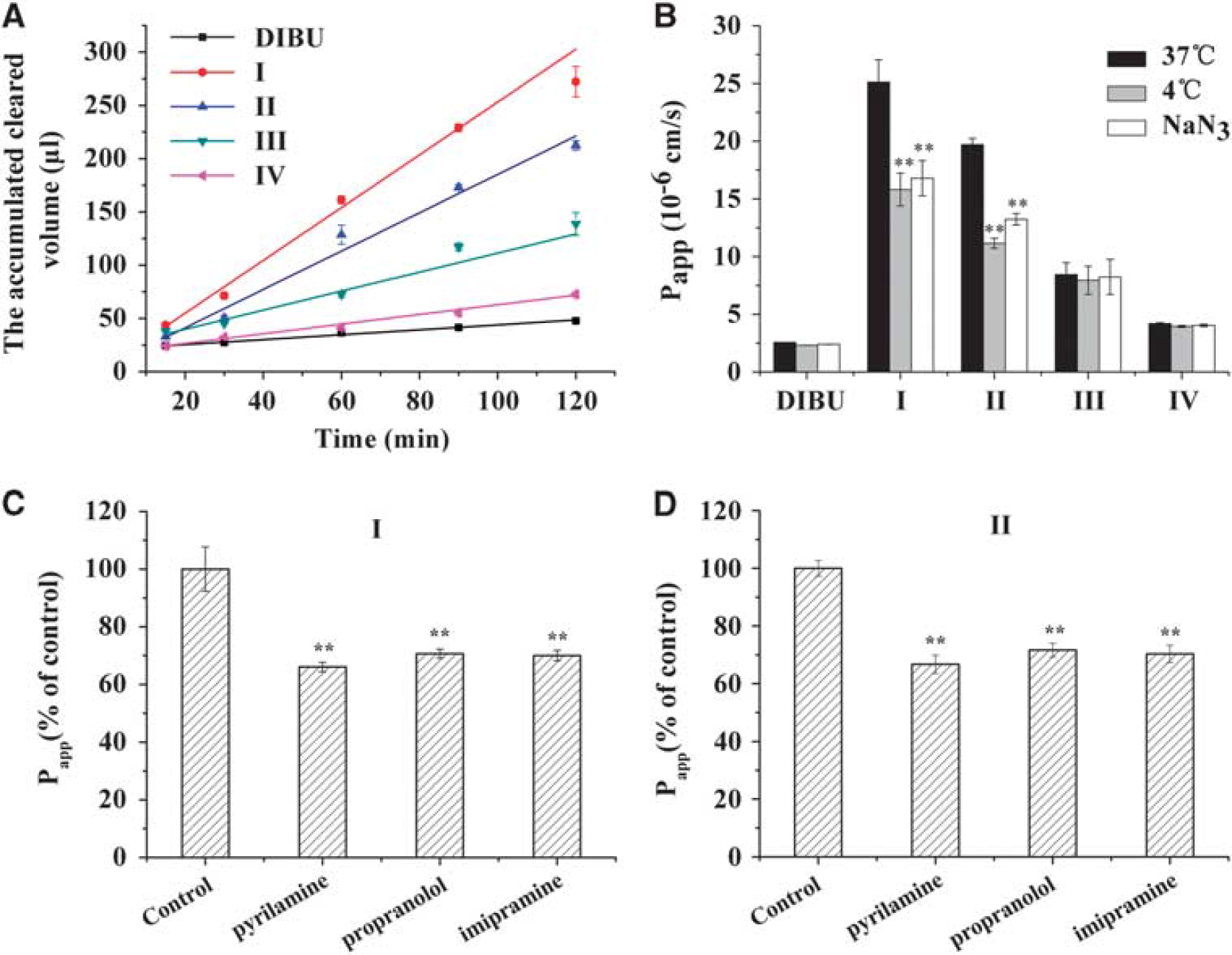
Mechanism of transendothelial transport of DIBU and prodrugs in an
Lineweaver-Burk analysis of prodrug I and II uptake in the presence or absence of pyrilamine showed that this cationic H1 antagonist competitively inhibited uptake of both prodrugs (Figures 5F and 5G); the observed Ki values were 102.61 umol/L for I and 123.35 μmol/L for II.
Transendothelial Transport of Prodrugs in an In Vitro Blood–Brain Barrier Model
In an effort to model how our prodrugs modified with ethanolamine-related groups may move across the blood–brain barrier into the brain parenchyma, we supplemented the BCEC monolayer studies with studies in an
Prodrug Transport from Blood to Brain in Rats
After
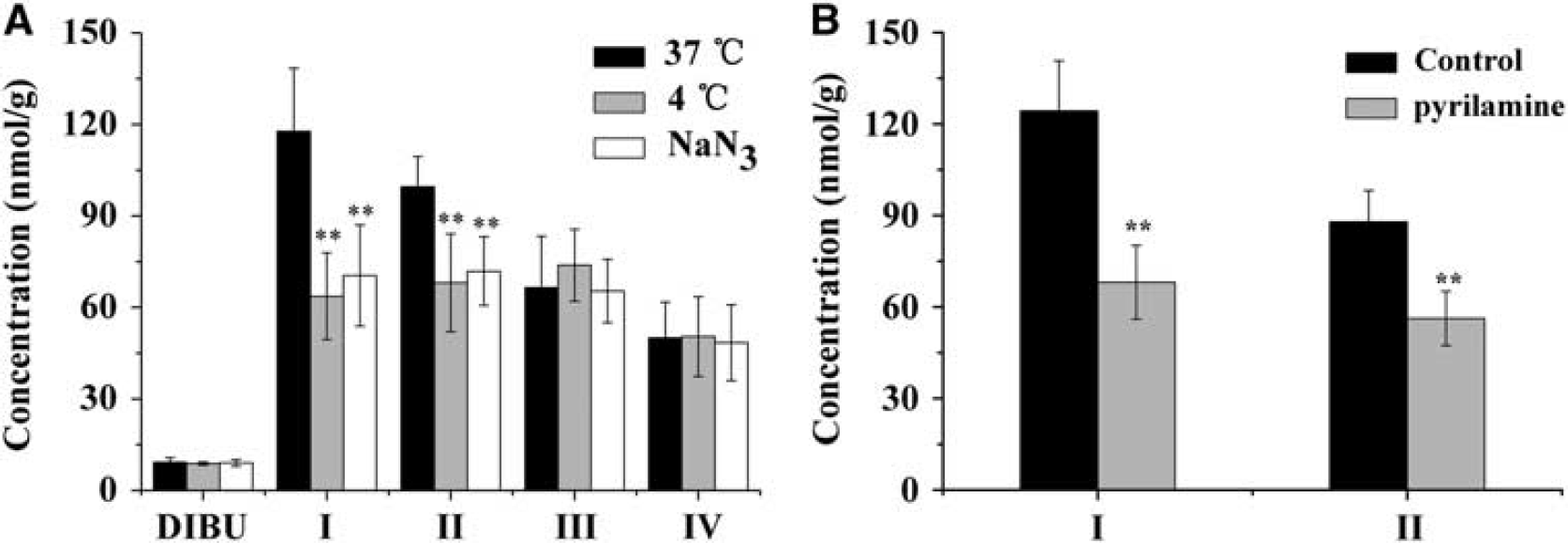
To verify the functional characteristics of the putative transport system observed
DISCUSSION
In previous work as part of our ongoing efforts to synthesize a new generation of drugs capable of reaching the brain in pharmacologically effective doses, we designed and synthesized a series of prodrugs modified with ethanolamine-related structures and showed that they delivered DIBU safely and efficiently to the brain. Nevertheless, despite sharing a similar structure, the four prodrugs exhibited dramatically different targeting efficiencies. Here, we extend that work by providing the first molecular insights into brain targeting of cationic drugs. Prodrugs III and IV appear to be taken up by an energy-independent, passive process. In contrast, prodrugs I and II appeared to be taken up by an active transport process depend on time, concentration, temperature, and energy. These results not only provide the basis for future, more detailed mechanistic studies; they also indicate that even prodrugs with globally similar structure can cross the BBB by quite different mechanisms.
Cytotoxicity of cationic materials or drugs has become a major concern in drug development. Modification with ethanolamine-related structures did not increase the cytotoxicity of prodrugs compared with DIBU (Figure 3A). Nicotine is reported to increase the BBB permeability via the modulation of BBB tight junctions.
28
Ethanolamine-related structures share the same structure unit with nicotine, and thus, we studied the effect of prodrugs on the tight junction of
Endothelial cell membranes are negatively charged, consisting mainly of phosphatidylcholine with smaller amounts of anionic phosphatidylserine. 29 The brain appears to absorb cationic peptides primarily as a result of their interaction with the anionic membrane, 30 and the membranes of brain cells are quite anionic, with their lipids relatively exposed. This may explain why lipophilic cationic compounds are more likely than other molecules to cross the BBB. 23 To analyze interactions between our prodrugs and the membrane of BCECs, we measured the surface charge (zeta potential) in the presence of prodrugs. The results suggest that modifying DIBU with ethanolamine-related structures increased affinity for the anionic surface of endothelial cells (Figure 3D), which may help enhance brain uptake. At the same time, the different ethanolamine modifications lead to the same extent of partial membrane neutralization, in contrast to their substantially different uptake efficiencies. We speculate that while the modifications globally increased interaction with BCECs, chemical and structural differences also strongly contribute to the overall transport efficiency and perhaps even to the underlying transport mechanism.
Primary BCECs express several transporters, including GluT1, LAT1, and OCTN2. Thus, BCECs appear to be a suitable cell line for investigating the BBB transport of prodrugs. In the cellular uptake study, conjugation of ethanolamine-related structures significantly improved the cellular uptake of dexibuprofen in BCECs (Figure 4). The uptake of III and IV was proven to be energy independent, indicating a passive uptake process. On the contrary, the uptake of I and II by BCECs was time, concentration, temperature, and energy dependent, indicating an active transport process is involved in their uptake. Competition experiments showed that cationic compounds (hemicholinium-3, TEA, and L-carnitine), choline, and its analogs did not inhibit uptake of I or II, nor did alkaline amino acids (Figures 5C and 5D). These results suggest that uptake of I and II does not involve OC transporters (OCTs and OCTNs), choline transporters, or the electrostatic interactions observed in the transport of cationic peptides. Instead, the lipophilic cationic drugs pyrilamine, imipramine, and propranolol significantly inhibited the uptake of I and II; in fact, pyrilamine competitively inhibited uptake of I with Ki value of 102.61 μmol/L and II with Ki value of 123.35 μmol/L. These results, together with the observation that transport was dependent on pH but independent of sodium and membrane potential (Figures 5A and 5B), suggest that I and II share a common H+/OC transporter with pyrilamine. 31
To further explore the fate of prodrugs after they are taken up into BCECs, we examined their transendothelial transport in a BCEC-AC coculture model of the BBB. This model relies on transwell culture inserts to create ‘contact through feet’ interface for interactions between the two cell types. In this coculture model, specific functional proteins such as enzymes, transporters, and tight junctional proteins are expressed, which likely help the system achieve a higher TEER than other BBB models.
32
Similar to the uptake process, the transendothelial transport of DIBU and of prodrugs III and IV was energy independent, suggesting a passive transport process. In contrast,
The substrate specificity of the H+/OC antiporter is controversial. While some speculate that it preferentially interacts with OCs with secondary or tertiary amine moieties,
16
others suggested that it recognizes only tertiary amines with
The lipophilicity of the amine modification may also be important. Kubo
34
conducted an analysis of the relationship between the inhibitory effect and the lipophilicity of inhibitors, revealed a lipophilicity-dependent inhibitory effect of amines on the H+/OC antiporter. Work from our laboratory supports the idea that lipophilicity of the
The
The present work provides the first mechanistic details about brain targeting by dexibuprofen prodrugs modified with ethanolamine-related structures.
Footnotes
YL, TG, and ZZ conceived and designed the study; YL performed the experiments and wrote the paper; YZ and JJ assisted with ‘Cellular uptake assay’; XW assisted with the ‘In situ rat brain perfusion study’; YF edited the paper.
The authors declare no conflict of interest.
ACKNOWLEDGMENTS
The authors would like to thank Prof. Linying Feng (Shanghai Institute of Materia Medica, Chinese Academy of Sciences) for the help with the construction of
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.