
Abstract
Thrombin and iron are two major players in intracerebral hemorrhage-induced brain injury and our recent study found that thrombin contributes to hydrocephalus development in a rat model of intraventricular hemorrhage (IVH). This study investigated the role of red blood cell (RBC) lysis and iron in hydrocephalus after IVH. There were three parts to this study. First, male Sprague-Dawley rats received an injection of saline, packed, or lysed RBCs into the right lateral ventricle. Second, rats had an intraventricular injection of iron or saline. Third, the rats received intraventricular injection of lysed RBCs mixed with deferoxamine (0.5 mg in 5 μL saline) or saline. All rats underwent magnetic resonance imaging at 24 hours and were then euthanized for brain edema measurement, western blot analysis, or brain histology. We found that intraventricular injection of lysed RBCs, but not packed RBCs, resulted in ventricular enlargement and marked increases in brain heme oxygenase-1 and ferritin at 24 hours. Intraventricular injection of iron also resulted in ventricular enlargement and ventricular wall damage 24 hours later. Coinjection of deferoxamine reduced lysed RBC-induced ventricular enlargement (
INTRODUCTION
Intraventricular hemorrhage (IVH) is often associated with intracerebral hemorrhage (ICH) and subarachnoid hemorrhage. 13 Hydrocephalus frequently occurs after IVH, developing in up to 55% of IVH patients. 1
Although it is well known that IVH is associated with hydrocephalus, the underlying mechanisms are still not clear. Evidence suggests a relationship between the incidence of posthemorrhagic hydrocephalus and the amount of intraventricular blood. In ICH, thrombin formation, red blood cell (RBC) lysis, and iron toxicity have a major role in brain injury after hemorrhage. Our recent study showed that thrombin contributes to IVH-induced hydrocephalus. 4
There have been few studies evaluating the relationship between iron released from blood into cerebrospinal fluid (CSF) and hydrocephalus; however, there is some clinical evidence of a link between iron and posthemorrhagic hydrocephalus. Nonprotein-bound iron was found in CSF from preterm infants with posthemorrhagic ventricular dilatation compared with infants without IVH. 5 In subarachnoid hemorrhage patients, a high level of the endogenous iron chelator ferritin in CSF is an independent predictive factor for chronic hydrocephalus. 6 In addition, iron and ferritin deposition are found in ependymal or subependymal location after neonatal IVH.7–9 Our recent studies indicate that iron may have a role in hydrocephalus development.10,11
In the current study, we investigated whether RBC lysis and iron cause hydrocephalus in adult rats.
MATERIALS AND METHODS
Animal Preparation and Intraventricular Injection
Animal use protocols were approved by the University of Michigan Committee on the Use and Care of Animals. The University of Michigan has an Animal Welfare Assurance on file with the Office for Protection from Research Risks and is fully accredited by the American Association for the Accreditation of Laboratory Animal Care. The studies follow the Guide for The Care and Use of Laboratory Animals (National Research Council) and comply with the ARRIVE guidelines for reporting
Experimental Groups
There were three parts to this study. First, 30 μL of saline, packed, or lysed RBCs was injected into the right lateral ventricle. All rats were euthanized at 24 hours for western blot analysis (
Magnetic Resonance Imaging and Ventricular Volume Measurement
Imaging was performed in a 7.0-T Varian MR scanner (183-mm horizontal bore; Varian, Palo Alto, CA, USA). Rats were anesthetized with 2% isoflurane/air mixture throughout the MRI examination. A T2 fast spin-echo sequence (repetition time/echo time = 4,000/60 ms) was performed. The field of view was 35 × 35 mm, and the matrix was 256 × 128 mm. A total of 25 coronal slices (0.5 mm thick) were acquired to cover the entire axis of the lateral ventricles. All image analysis was performed using NIH Image J (NIH, Bethesda, MD, USA). Lateral ventricular volumes were calculated from T2 images as described previously. 13 Both lateral ventricles were outlined and the areas were measured. Ventricular volume was obtained by combining the ventricle area over all slices incorporating the lateral ventricles and multiplying by section thickness (0.5 mm). A blinded observer performed all the measurements. It is important to differentiate the edematous white matter from the lateral ventricles on T2 MRI. Lateral ventricle volumes were measured on T2 images based on the brain anatomy and were confirmed by brain histology.
Western Blot Analysis
Western blot analysis was performed as previously described. 10 The brains were perfused with saline before decapitation. The periventricular zone (PVZ) (~1 mm thick brain tissue around the ventricle) was sampled. Protein concentration was determined by the Bio-Rad protein assay kit (Hercules, CA, USA) and 50 μg protein from each sample was separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto a Hybond-C pure nitrocellulose membrane (Amersham, Pittsburgh, PA, USA). The primary antibodies were polyclonal rabbit anti-rat heme oxygenase-1(HO-1) IgG (1:2,000 dilution; StressGen, Farmingdale, NY, USA), rabbit anti-ferritin heavy chain polyclonal antibody (1:2,000 dilution; Cell Signaling Technology, Danvers, MA, USA) and goat anti-ferritin light chain polyclonal antibody (1:2,000 dilution; Abnova, Taipei, Taiwan). The antigen-antibody complexes were visualized by chemiluminescence system (Denville Scientific Inc., Metuchen, NJ, USA) and exposed to photosensitive film. The relative densities of bands were analyzed with NIH Image J.
Immunohistochemistry
Rats were anesthetized with pentobarbital (100 mg/kg, intraperitoneally) and perfused with 4% paraformaldehyde in 0.1 mol/L pH 7.4 phosphate-buffered saline. The brains were removed and kept in 4% paraformaldehyde for 4 to 6 hours, then immersed in 30% sucrose for 3 to 4 days at 4°C. The brains were embedded in optimal cutting temperature compound (Sakura Finetek USA, Inc., Torrance, CA, USA) and 18-m-thick slices were cut using a cryostat. Immunohistochemistry was performed using the avidin–biotin complex technique as previously described. 10 The primary antibodies were polyclonal rabbit anti-human ferritin IgG (DAKO, Glostrup, Denmark; 1:400 dilution) and polyclonal rabbit anti-rat HO-1 IgG (StressGen, 1:400 dilution).
Electron Microscopy
Electron microscopy was performed as previously described. 14 Rats were anesthetized and subjected to intracardiac perfusion with 4% paraformaldehyde and 2.5% glutaraldehyde in 0.1 mol/L Sorensen's buffer (pH 7.4). The brains were removed and a 1-mm-thick coronal brain slice was cut with a blade ~4 mm from the frontal pole. The ipsilateral ventricular wall tissue was sampled and immersed in the same fixative overnight at 4°C. Samples were then postfixed with 1.0% OsO4 and dehydrated in graded ethyl alcohol. After complete dehydration, samples were infiltrated with propylene oxide, embedded in Epon, and sectioned.
Ultra-thin sections were then stained with uranyl acetate and Reynold's lead citrate, and evaluated using Philips CM 100 TEM (Hillsboro, OR, USA) and digitally imaged using a Hamamatsu (Hamamatsu City, Shizuoka, Japan) ORCA-HR camera.
Brain Water Content
The determination of brain water content was performed as described previously. 15 Briefly, the rat brain was removed after euthanasia and divided into left hemisphere, right hemisphere, and cerebellum. After weighing, the samples were dried in a gravity oven for 48 hours. The water content was determined as (wet weight — dry weight)/wet weight.
Statistical Analysis
Values are given as mean ± S.D. Student's
RESULTS
Physiologic parameters of rat, including mean arterial blood pressure, blood gas, hematocrit, blood pH, and glucose were monitored and controlled in normal ranges. Mortality in the lysed erythrocyte group was 19% (5 of 27) and zero in all other groups.
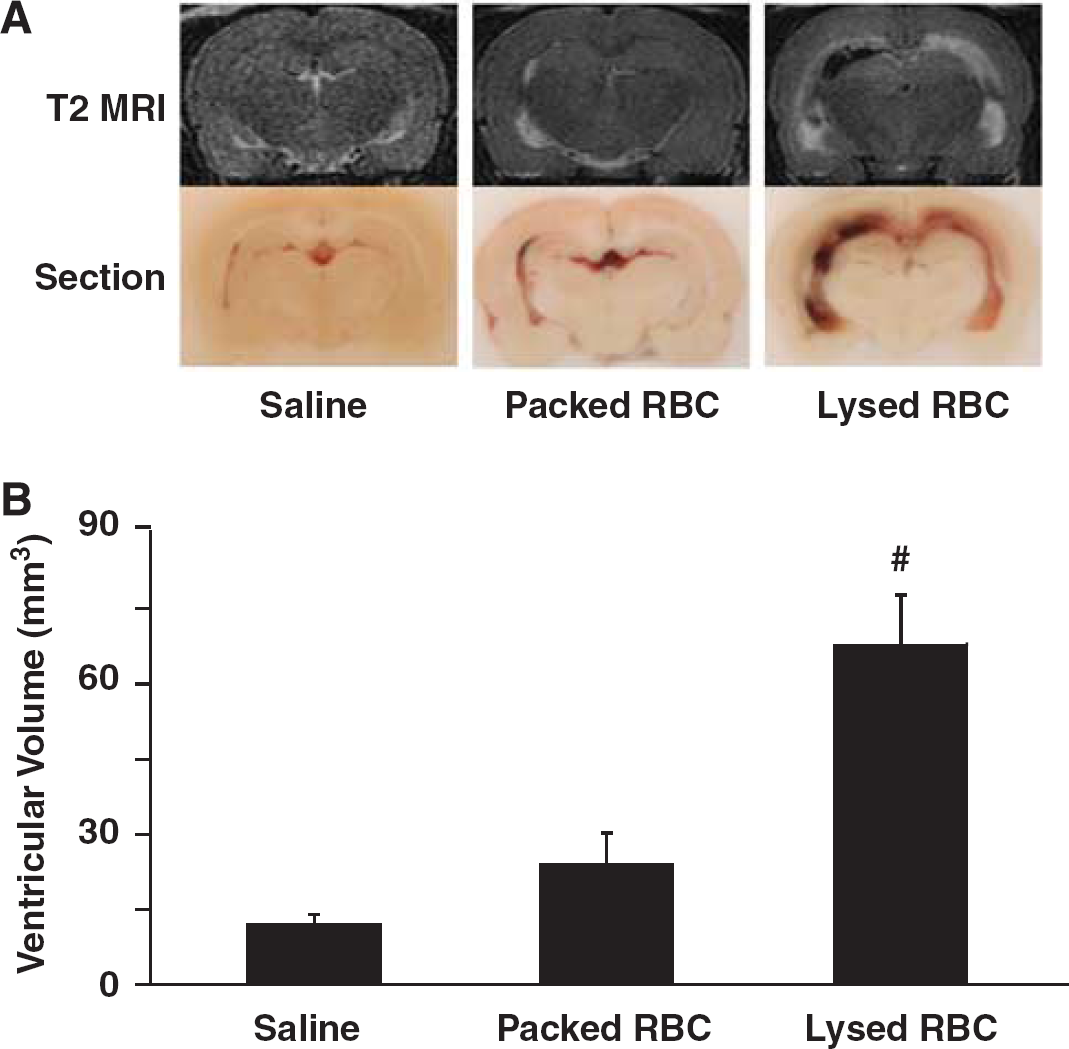
Lysed Red Blood Cells But Not Packed Red Blood Cells Result in Hydrocephalus
To examine the effect of erythrocyte lysis on hydrocephalus development, saline, packed RBCs, or lysed RBCs were injected into the right lateral ventricle. Injection of lysed RBCs resulted in significantly larger ventricular volumes (lateral ventricle volume 67.9 ± 10.0 mm3) compared with injection of packed RBCs (24.1 ± 6.2 mm3,
Intraventricular Injection of Lysed Red Blood Cells Results in Upregulation of Periventricular Heme Oxygenase-1 and Ferritin
The expression of HO-1, a key enzyme for heme degradation, was evaluated in periventricular tissue. There were many more periventricular HO-1-positive cells after injection of lysed RBCs than after saline or packed RBC injection. Heme oxygenase-1-1-positive cells were mostly gliallike (Figure 2). HO-1 protein levels in the periventricular area were significantly increased in rats receiving lysed RBCs (3,566 ± 2,481 pixels vs. 115 ± 141 pixels in saline group and 295 ± 311 pixels in the packed RBC group,
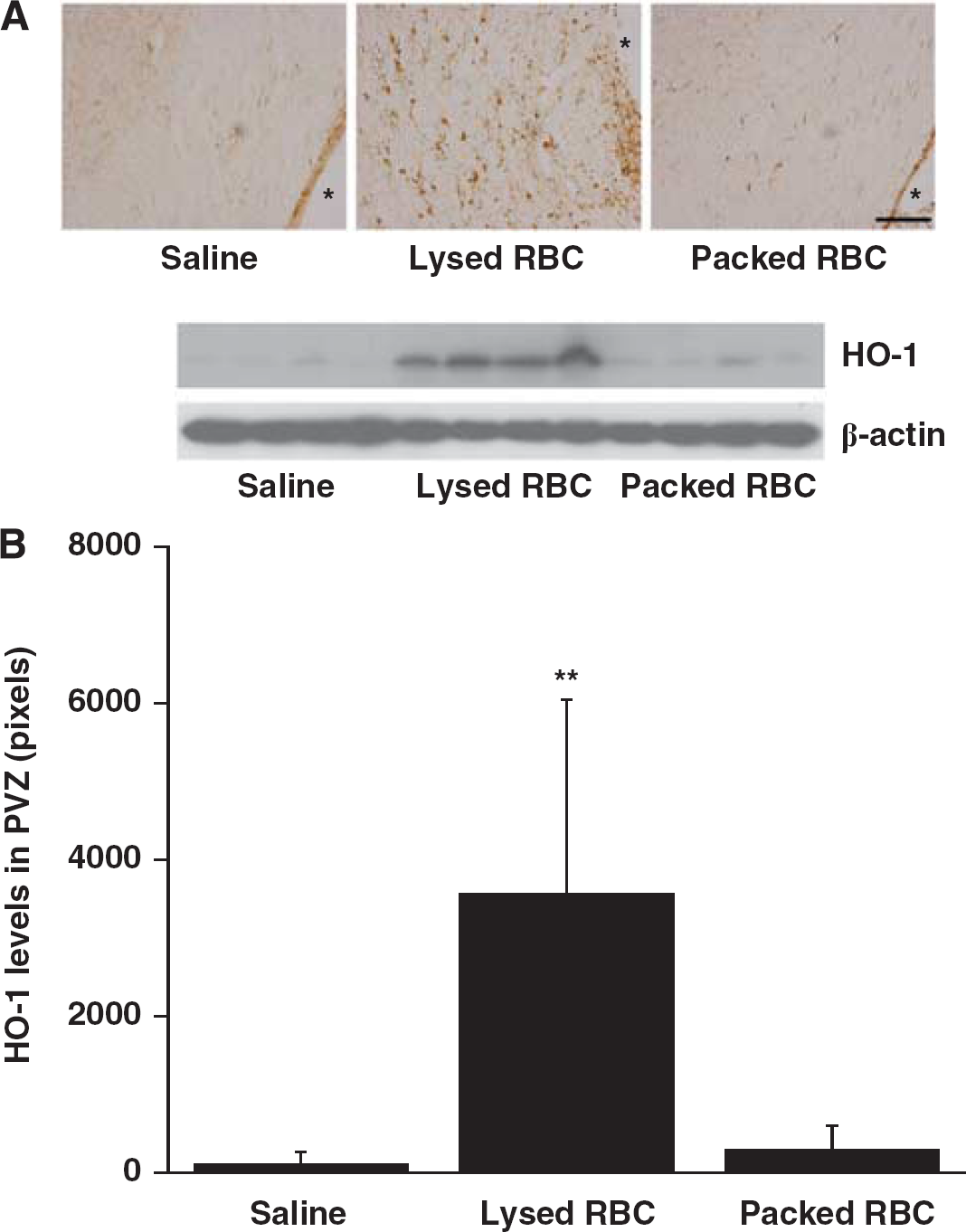
Heme oxygenase-1(HO-1) immunoreactivity
Ferritin was also investigated in the periventricular area 24 hours after injection as it is an iron storage protein. There was a marked increase in periventricular ferritin immunoreactivity after injection of lysed RBCs (Figure 3A) compared with saline or packed RBC injection. Most ferritin-positive cells were also gliallike although there was also evidence of expression in the ependyma. Ferritin upregulation in the PVZ was confirmed by western blotting. Injection of lysed RBCs resulted in higher protein levels of ferritin heavy chain (2,350 ± 429 pixels vs. 800 ± 326 pixels in packed RBC group,
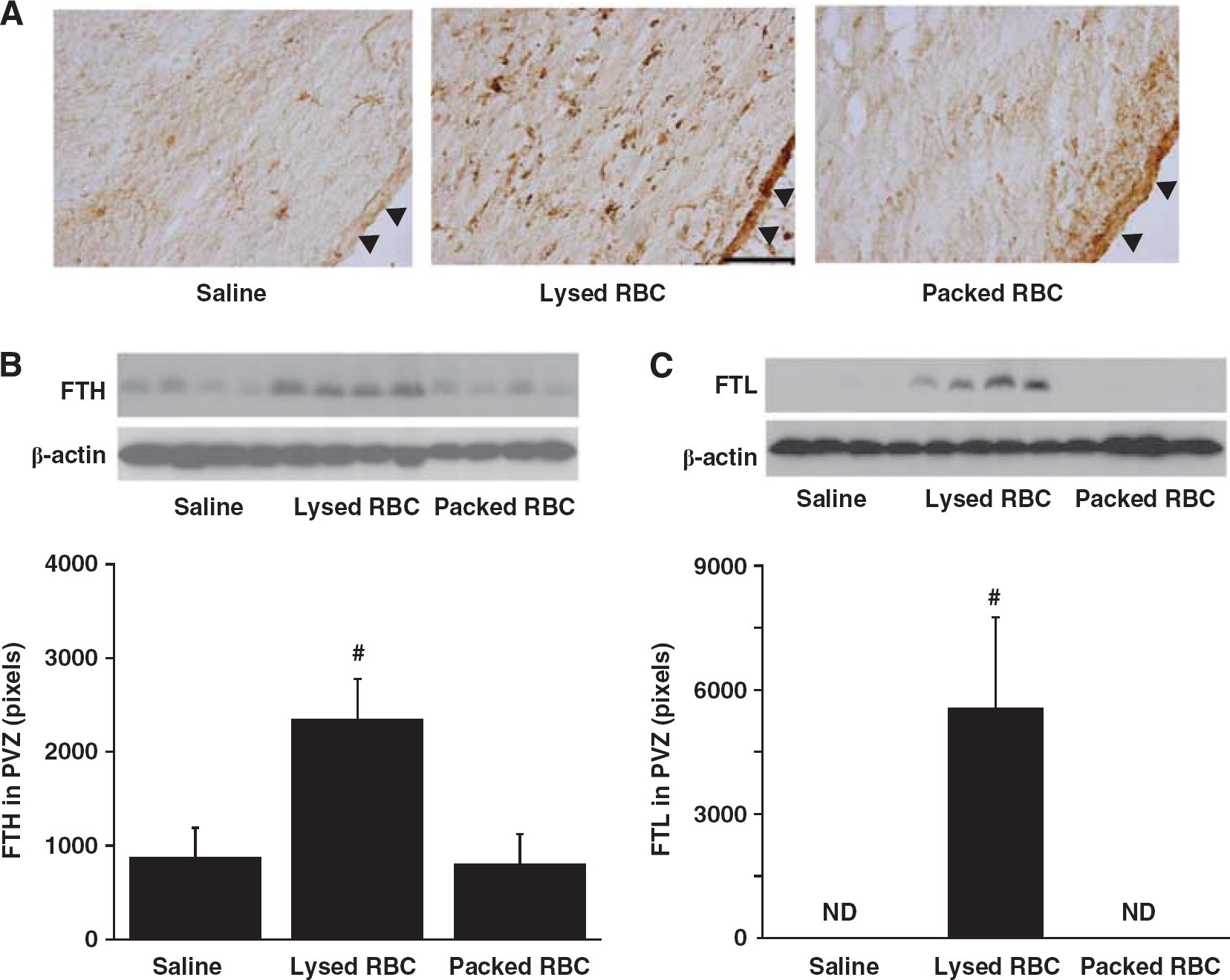
Ferritin immunoreactivity
Intraventricular Injection of Iron Results in Ventricular Enlargement and Ependymal Cell Injury
To evaluate the effect of iron on hydrocephalus development, rats received an injection of either 50 μL iron (Fe3+) or saline into the right lateral ventricle. Twenty-four hours after injection, T2 MRI showed a significant bilateral ventricular enlargement in the iron-injected group (17.9 ± 5.7 mm3) compared with saline-injected rats (9.2 ± 5.7 mm3,
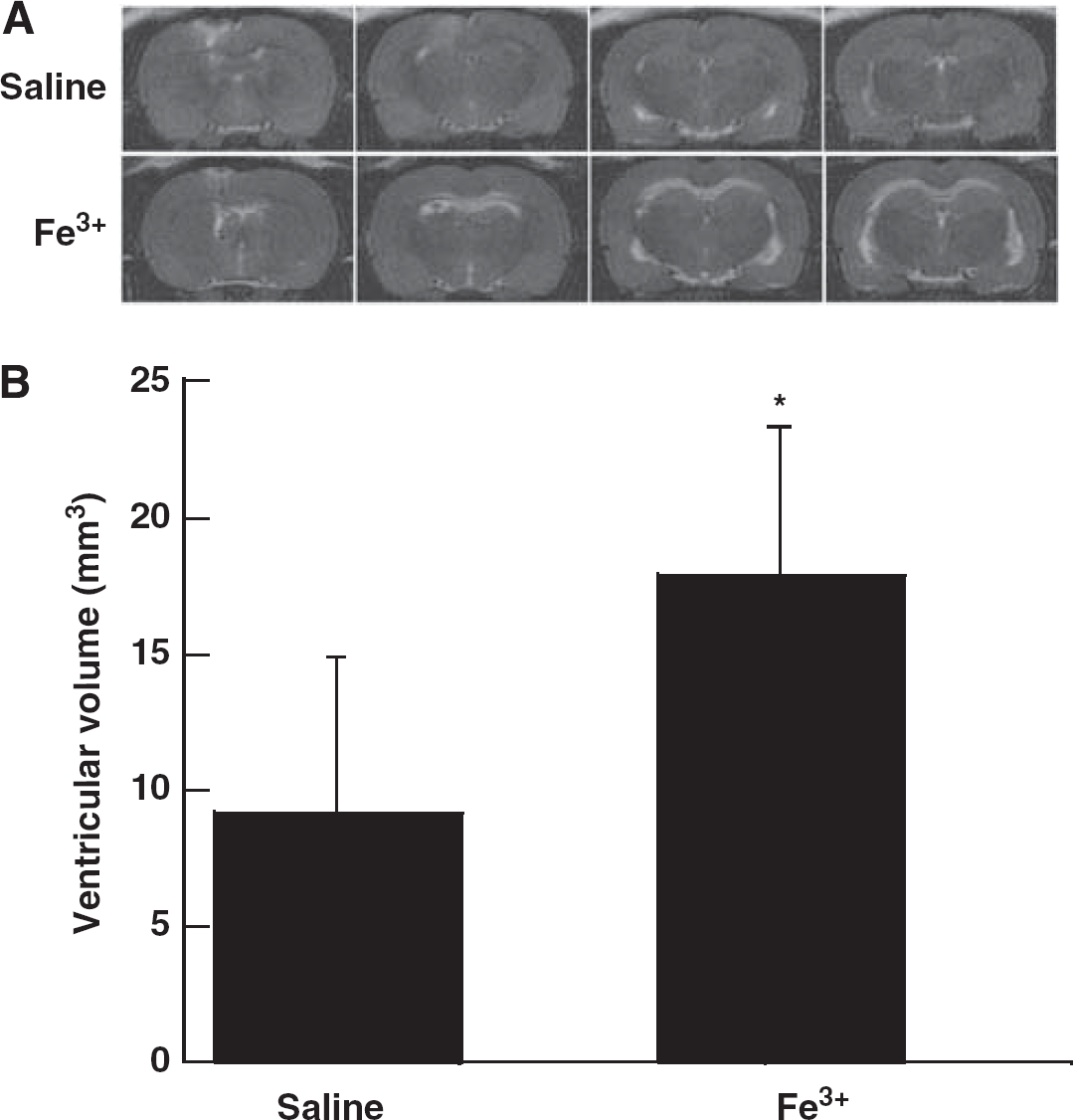
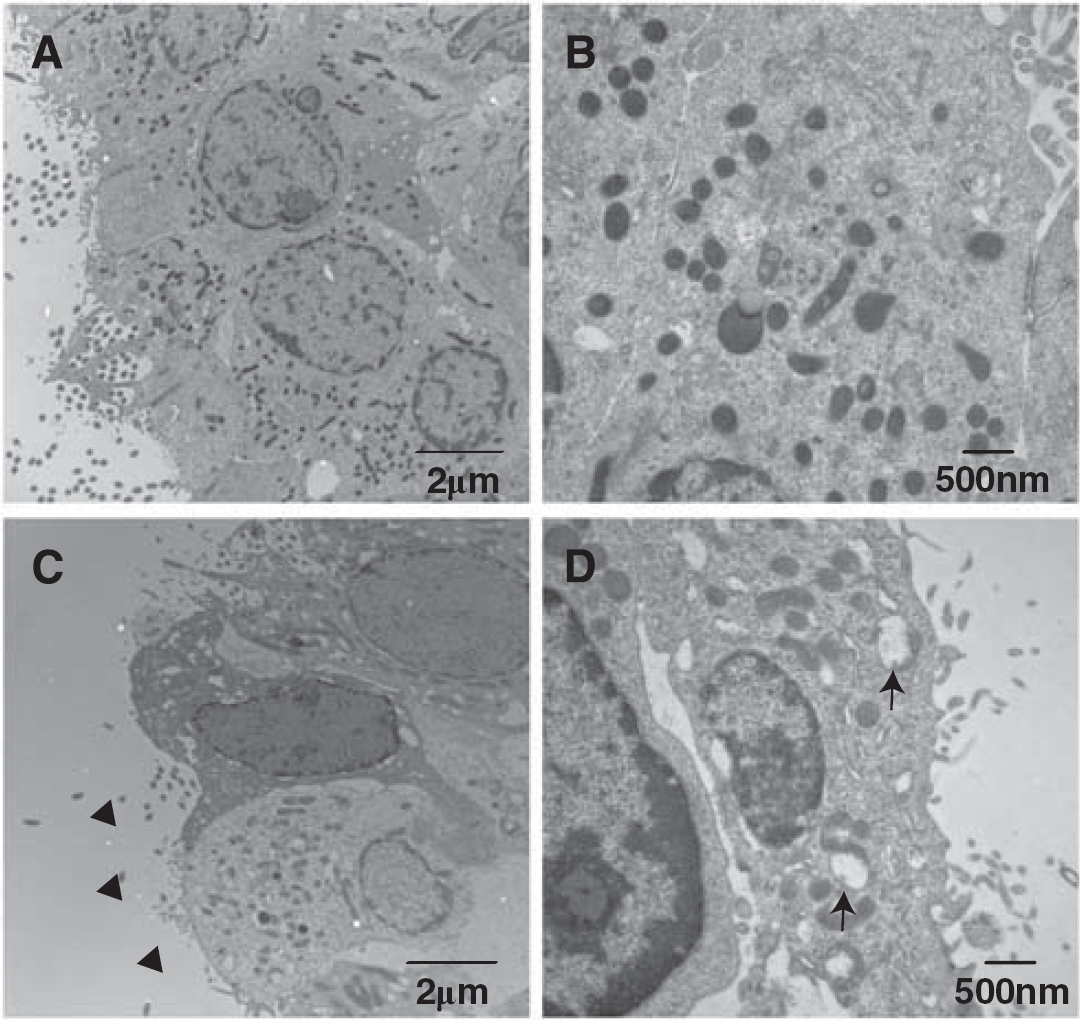
Electron micrographs showing the ultrastructure of ependymal cells of the ipsilateral right ventricle at 24 hours after 50 μL FeCl3 injection (
Deferoxamine Attenuates Lysed Red Blood Cell-Induced Ventricular Enlargement
Deferoxamine, an iron chelator was used to evaluate the role of iron in ventricular enlargement after intraventricular injection of lysed RBCs. Coinjection of deferoxamine with lysed RBC reduced ventricular enlargement (47.8 ± 17.8 vs. 59.9 ± 15.7 mm3 in saline coinjection group,
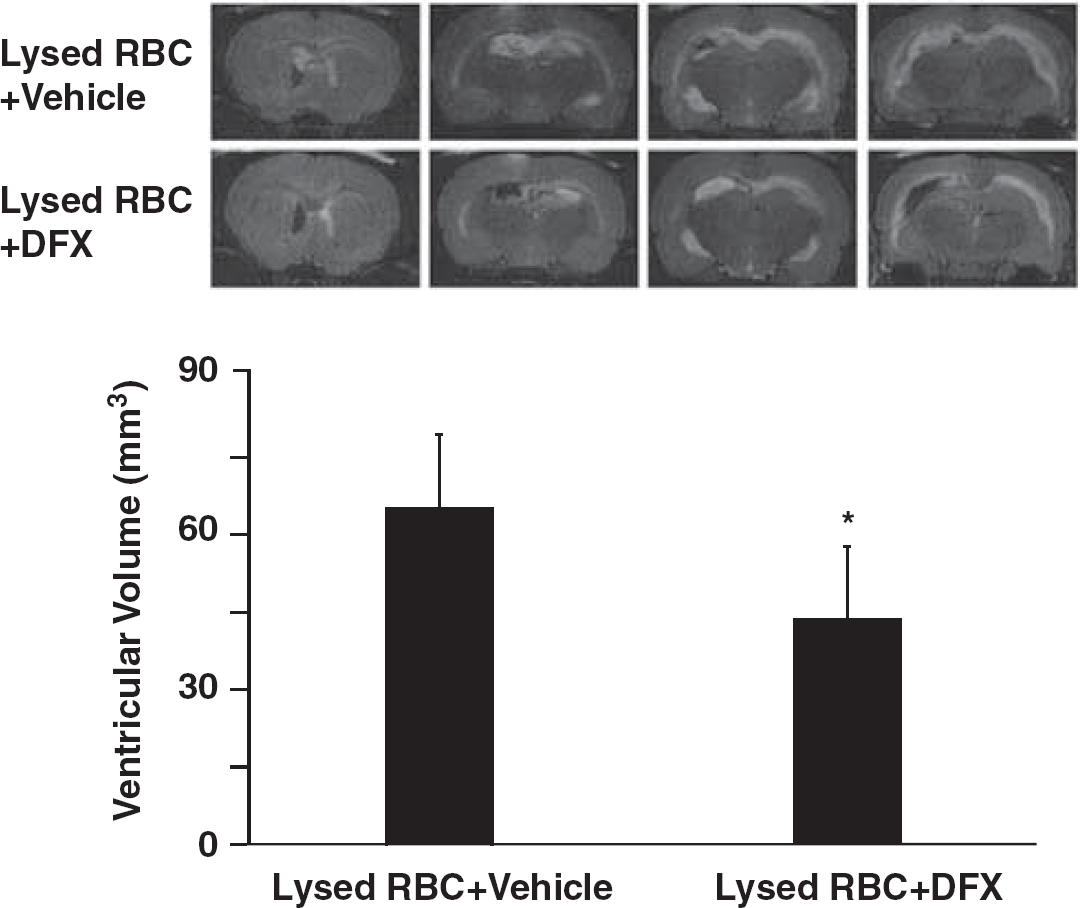
T2-weighted magnetic resonance imaging (MRI) showing lateral ventricle volumes 24 hours after intraventricular injected of lysed red blood cells (RBCs) mixed with DFX or saline. Values are means ± s.d.,
DISCUSSION
The major findings of this study are (1) intraventricular injection of lysed RBCs but not packed RBCs resulted in hydrocephalus; (2) lysed RBCs upregulated brain HO-1 and ferritin levels; (3) intraventricular injection of iron also caused hydrocephalus; and (4) iron chelation with deferoxamine reduced lysed RBC-induced hydrocephalus. Our previous studies have shown that IVH resulted in prolonged hydrocephalus.10,16 In this proof-of-concept study, hydrocephalus was determined at 24 hours to avoid brain tissue loss-related ventricle enlargement.
Blood components, including thrombin and iron, have a major role in brain injury after ICH.17,18 Our most recent study has shown that thrombin and thrombin receptor are involved in IVH-induced hydrocephalus. 4 The current study found that RBC lysis and iron toxicity can cause hydrocephalus. These results suggest a major role of blood components in hydrocephalus development after IVH. Indeed, it is possible that the effects of iron and thrombin may combine to exacerbate IVH-induced injury, as found in ICH. 19 In this proof-of-concept study, deferoxamine was given by intraventricular injection. Our previous study has shown that systemic use of deferoxamine can reduce IVH-induced hydrocephalus. 10 Future experiments should determine therapeutic time window of systemic deferoxamine treatment and whether combination therapy is needed for IVH-induced hydrocephalus.
Iron levels in plasma are low, however, iron concentrations are very high in RBCs. 20 When RBCs start to lyse, hemoglobin degradation can result in iron release. The perihematomal iron concentration could reach as high as 10 mmol/L, which causes marked brain damage. 21 The form of the iron within heme depends on whether there is oxyhemoglobin (Fe2+) or methemoglobin (Fe3+). In initial experiments, we examined the effect of intraventricular Fe2+ and Fe3+, both caused brain injury. The current study focused on Fe3+. High levels of methemoglobin have been reported in CSF after IVH.22,23 We will continue to pursue the potential role of Fe2+ in IVH-induced brain injury in other studies, but this is hampered by a lack of specific chelators available for ferrous iron.
There is increasing evidence that iron is involved in brain injury after cerebral hemorrhage.17,18 It is hypothesized that lysis of RBCs within the hemorrhage cause release of hemoglobin, which is metabolized by heme oxygenases resulting in the release of iron. In a rat IVH model, we have found that iron deposition after IVH is present in the first day and upregulation of brain ferritin lasts for several weeks. 10 It is well known that brain iron overload can stimulate free radical production and cause oxidative injury, including lipid peroxidation and DNA damage.18,24,25 The current study shows that injection of lysed RBCs, but not packed RBCs, can mimic the acute ventricular enlargement induced by IVH. 10
Red blood cell lysate contains hemoglobin and its components iron and heme, and although multiple factors could have a role in ventricular enlargement, we show that coinjection with an iron chelator, deferoxamine, reduces ventricular enlargement induced by lysed RBCs. In addition injection of FeCl3 also results in ventricular enlargement further supporting the role of iron in posthemorrhagic hydrocephalus. This pinpoints the importance of iron in hydrocephalus after RBC lysis. It should be noted that two proteins that have crucial roles in iron regulation, HO-1 and ferritin, were markedly increased in the PVZ after injection of lysed RBCs. Heme oxygenase-1 is involved in heme metabolism and the release of iron and ferritin is the major endogenous cellular iron chelator. 25 An upregulation of ferritin after injection of lysed RBCs or IVH10,26 may limit iron-induced brain injury. Indeed, upregulation in the ependyma may limit the penetration of iron into the rest of the brain. Whether HO-1 may be a therapeutic target in IVH, by limiting the rate of iron release, has not been studied. There is, though, evidence in ICH that HO inhibitors reduce injury. 27
Iron chelation may have multiple effects on IVH-induced brain injury. Our finding that iron can mimic the effect of IVH on ventricular dilation indicates a direct effect of iron (and iron chelation) on the brain in the absence of RBCs. However, we have found that iron chelation alters the rate of hematoma resolution 28 and there are other components of RBCs that may contribute to brain injury (e.g., carbonic anhydrase, 29 ).
Acute hydrocephalus is mostly considered as being the result of reduced CSF absorption. For example, in a prior study from our group on a rat IVH model, hydrocephalus was associated with a threefold increase in CSF absorption resistance suggesting a physical block in the CSF drainage pathways. 16 However, in the current study, injection of both lysed RBCs and FeCl3 induced acute hydrocephalus. Neither is likely to cause an acute physical blockage of the CSF drainage pathways. Another possibility is the iron-related hydrocephalus may result from ependymal cell injury. Ependymal cells overlie the cerebral ventricles and their apical surface is covered by cilia that beat in a coordinated manner to facilitate CSF microcirculation. There is an increasing body of evidence that genetic mutants that affect ependymal cilia structure or function also cause hydrocephalus.30–32 Our results showed that iron can cause ependymal cell death and loss of cilia suggesting the possibility that ependymal cell injury may result in defective CSF dynamics and aggravate hydrocephalus. It is difficult to translate from studies on genetic mutants to acute injury and more studies are needed on the relationship between acute ependymal changes and hydrocephalus.
In conclusion, this study provides evidence that products produced by erythrocyte lysis and, particularly, iron can induce hydrocephalus and may be a therapeutic target for IVH. The effects of erythrocyte lysate and iron in inducing hydrocephalus may be related to ependymal damage.
Footnotes
The authors declare no conflict of interest.