
Abstract
Excitotoxicity is a major contributor to cell death during the acute phase of ischemic stroke but aggressive pharmacological targeting of excitotoxicity has failed clinically. Here we investigated whether pretreatment with low doses of memantine, within the range currently used and well tolerated for the treatment of Alzheimer's disease, produce a protective effect in stroke. A coculture preparation exposed to modeled ischemia showed cell death associated with rapid glutamate rises and cytotoxic Ca2+ influx. Cell death was significantly enhanced in the presence of high memantine concentrations. However, low memantine concentrations significantly protected neurons and glia via excitotoxic cascade interruption. Mice were systemically administered a range of memantine doses (0.02, 0.2, 2, 10, and 20 mg/kg/day) starting 24 hours before 60 minutes reversible focal cerebral ischemia and continuing for a 48-hour recovery period. Low dose (0.2 mg/kg/day) memantine treatment significantly reduced lesion volume (by 30% to 50%) and improved behavioral outcomes in stroke lesions that had been separated into either small/striatal or large/striatocortical infarcts. However, higher doses of memantine (20 mg/kg/day) significantly increased injury. These results show that clinically established low doses of memantine should be considered for patients ‘at risk’ of stroke, while higher doses are contraindicated.
Introduction
Cerebral stroke is the third leading cause of death in developed countries and the leading cause of adult disability. The only licensed pharmacological treatment available for acute cerebral stroke is thrombolysis with recombinant tissue plasminogen activator but because of its narrow therapeutic window (<4.5 h) and safety concerns, approximately only 15% of stroke patients receive recombinant tissue plasminogen activator.
1
Focal cerebral ischemia initiates a cascade of complex pathophysiologic events including excitotoxicity, acidotoxicity, ionic imbalance, oxidative stress, inflammation, and apoptosis.2, 3, 4 In an attempt to limit the acute cell death after the onset of ischemia a variety of neuroprotective strategies have been developed, which aim to antagonize injurious biochemical and molecular events that culminate in neuronal death. However, such strategies have failed to translate effectively into clinically available treatments for stroke patients (see Davis
The most rapid, and possibly the most severe, pathophysiologic mechanism initiated after ischemic stroke is that of excitotoxicity, which triggers widespread necrosis and subsequent functional impairment.6, 7 Historically, the focus of neuroprotection research has been to use strategies that supress the excitotoxic response after induction of ischemia and primarily target the glutamate system.
8
Various NMDA (
To exert a beneficial effect after ischemic stroke the majority of NMDA receptor antagonists have to be administered at high doses, which increases the possibility of producing unwanted psychotomimetic and cardiovascular effects. 18 However, not all NMDA receptor antagonists produce unwanted side effects at clinically effective doses and some are in current clinical use. One such NMDA receptor antagonist, memantine (1-amino-3,5-dimethyladamantane), has been approved since 2002 in Europe (and since 2003 in the USA) for the treatment of moderate to severe Alzheimer's disease, 19 and has been shown in clinical trials to be a safe and effective treatment for vascular dementia. 20 Memantine is distinct from other NMDA receptor antagonists as it possesses fast on/off kinetics, low-moderate receptor affinity and is able to block the effects of excessive glutamate without interfering with the physiologic activation of NMDA receptors. 21 Although acute postischemic administration of other NMDA receptor antagonists has been shown to be protective in experimental models (for review see Lipton, 2004) there is a wealth of evidence showing that this therapeutic approach of trying to limit cell death during the acute phase of ischemia is difficult to achieve. Thus, a more realistic approach may be prophylactic administration of memantine to those patients identified at being at risk of ischemic stroke, at a dose already tolerated in clinical practice, e.g., for the treatment of Alzheimer's disease. The aim of this study was to determine if systemic administration of the antiexcitotoxic drug, memantine hydrochloride, at doses currently administered for moderate to severe Alzheimer's disease, could produce protection from ischemic stroke.
Material and Methods
Cell Culture
High-density cultures (HDCs): Cortices were obtained from E16 balb-c mice after humane cervical dislocation under UK Home Office regulations. UK home office regulations were followed for all experimental work which was conducted in accordance with the relevant guidelines and regulations. The animal welfare and ethics committee of University of Leicester approved all the experimental protocols. The tissue was placed in Hank's balanced salt solution, trypsinized (1% trypsin/DNase), triturated, centrifuged (250
Cell Culture Characterization
Cover slips were washed (0.1 mol/l phosphate-buffered saline (PBS)), fixed (4% paraformaldehyde/PBS or methanol/acetone 1:1), permeabilized (PBS/10% goat serum (Dako, Cambridge, UK)/0.5% Triton-X (Sigma-Aldrich, Gillingham, UK), PBSGT), incubated (PBSGT overnight at 4°C with primary antibody), exposed to appropriate secondary antibody (60 minutes PBSGT), mounted (SuperFrost Plus slides, Menzel-Glaser, Braunschweig, Germany) in PermaFluor (Thermo Fisher Scientific, Loughborough, UK), and imaged using a Leica (Milton Keynes, UK) TCS SP2 confocal microscope. Primary antibodies: glial fibrillary acidic protein and neuron specific enolase (1:400, Sigma-Aldrich); NSE (prediluted, Sigma-Aldrich); CNPase (1:100, Chemicon, Nottingham, UK); IB-4 (1:100, Molecular probes, Fisher Thermo Scientific). Projections (eight slices) were viewed using Leica software, Fluoview (Olympus, Southend-on-Sea, UK), or Metamorph (Molecular Devices, Sunnyvale, CA). Multiple slide areas were imaged and total cell number established either using the Hoechst 33,342 nuclear stain or light images of the cells; both methods produced similar cell counts (Supplementary Figure S1). At least three separate cell cultures and 3 to 8 slides were analyzed from each culture.
Cell Imaging
Oxygen—glucose deprivation (OGD)-induced intracellular Ca2+ ([Ca2+]i) changes were assessed using FURA-2FF (Invitrogen, Fisher Thermo Scientific), a low affinity dye that does not affect cell viability during ischemia. 22 The more sensitive FURA-2 was used for agonist responses. In both cases, cells were acetoxymethyl-loaded (see Fern 23 for more details of imaging methods). FURA dyes tended to leak from astrocytes over longer recording periods so, cell viability during OGD was assessed using 5-chloromethylfluorescein diacetate (Invitrogen) acetoxymethyl-loaded at 2.5 μmol/l. Dye-loaded cultures were mounted into a perfusion chamber (atmosphere chamber, Warner Instruments, Hamden, CT, USA), perfused at 2 ml/min (artificial cerebrospinal fluid in mmol/l: NaCl, 126; KCl, 3; NaH2PO4, 2; MgSO4, 2; CaCl2, 2; NaHCO3, 26; and glucose, 10; pH, 7.45, bubbled with 5% CO2/95% O2) and maintained at 37°C on the stage of an epifluorescence microscope (Nikon, Kingston Upon Thames, UK). Oxygen—glucose deprivation involved switching to artificial cerebrospinal fluid containing no glucose prebubbled with 95% N2/5% CO2, while chamber atmosphere was switched from 95% O2/5% CO2 to 95% N2/5% CO2. Cell temperature was monitored and maintained via flow-through, objective, and room heaters. A rapid exchange perfusion system was used for short application agonist experiments (ValveBank8.2, AutoMate Scientific, Berkeley, CA, USA). Oil immersion x20 images were collected at 520 nm or 508 nm using appropriate filter sets (Chroma Technology Corporation, Bellows Falls, VT, USA). For FURA-loaded cultures, cells were illuminated at 340, 360, and 380 nm; 5-chloromethylfluorescein diacetate cells were illuminated at 489 nm (Optoscan, Cairn Research). Images were captured by a coolSNAP HQ camera (Roper Scientific, Sarasota, FL, USA) controlled via MetaFluor (Molecular Devices) with background signal subtracted. For FURA-2 imaging, 340:380 was converted to [Ca2+]i using a calcium calibration kit (Invitrogen). Cell death was characterized by sudden collapse of the fluorescent signal to the background level and this phenomenon was used to calculate cell death rates and precise time points of cell death for all cells within the field of view. Cells were also imaged before and after the experiment in quadrants surrounding the field of view, where initial cell counts and surviving cell counts were measured. Cell death data plotted as a time series represents the real-time recordings from the field of view, while total cell death data includes the cells from the surrounding quadrants.
Cultured astrocytes and neurons are morphologically distinct under phase contrast. The majority of neurons are high contrast which have either pyramidal, fusiform, or multipolar characteristics. 24 Astrocytes show low phase-contrast with a flat morphology forming a continuous layer once confluent. These criteria for identification were tested and confirmed using immunohistochemical staining of fixed cocultures for glial fibrillary acidic protein and neuron specific enolase, (Supplementary Figure S2). The criteria were used to distinguish cells using initial phase-contrast and fluorescence images. Any unidentifiable cells were excluded from subsequent analysis.
Biosensors
Glutamate microelectrode biosensors (Sarissa Biomedical, Coventry, UK),
25
amplified via a Duo-Stat ME-200+ potentiostat (Sycopel International, London, UK) were used to record real-time glutamate or adenosine triphosphate concentration changes
Focal Cerebral Ischemia
This study was conducted in accordance with the UK Animals (Scientific Procedures) Act, 1986 (Project License 60/4315). Male adult C57 BL6 mice (Charles River, Oxford, UK) weighing between 22 and 32 g at the time of surgery were randomly assigned to a treatment group (the surgeon was masked to treatment and subsequent analyses). A total of 69 mice were used, 7 mice were excluded because of severe blood loss during surgery or poor recovery after middle cerebral artery occlusion (MCAO). Treatment groups were vehicle (
Mice were anesthetized with isoflurane (induction 4%; maintenance 1.5% in N2O/O2 70/30%). A small subcutaneous incision was made on the midflank and the osmotic mini-pump inserted. The wound was sutured and animals recovered for 24 hours before focal ischemia. Focal cerebral ischemia was induced for 60 minutes by occlusion of the right middle cerebral artery as previously described. 26 Body temperature was monitored throughout surgery (rectal probe) and maintained at 37.0°C±0.6°C using a heating mat (Harvard Apparatus, Holliston, MA, USA). Laser Doppler flowmetry (Moor Instruments, Axminster, UK) was used to monitor relative cerebral blood flow for 5 minutes before and 5 minutes after MCAO. After 60-minute MCAO, mice were reanesthetized and the occluding filament withdrawn. Mice were weighed at 24 and 48 hours after surgery and neurologic status assessed using a 28-point neurologic score. 27 At 48 hours after surgery, mice were killed via cervical dislocation and brains were removed, sectioned (10 × 1 mm coronal slices) and stained (2% 2,3,5-triphenyltetrazolium chloride in saline) for 30 minutes. 2,3,5-Triphenyltetrazolium chloride is a marker of mitochondrial function and has been shown to be a reliable indicator of ischemic areas for up to 3 days after ischemia. 28 Sections were stored in 10% formalin solution at 4°C and photographed for analysis. Infarct areas were calculated as previously described 29 using an indirect method whereby overestimation of the infarct area because of edema is avoided.
Statistical Analysis
In focal ischemia experiments, lesion volume, behavior, and body weight assessment were performed (by MT who was masked) and are presented as mean±s.e.m. The population distribution for lesion volume in the vehicle infusion group was not uniformly distributed, falling into two groups: small lesions located in the striatum and large lesion encompassing both the striatum and cortical areas. When separated into these two patterns of injury, lesion volume in the vehicle group had a normal distribution and a parametric one-way analysis of variance (Fisher's) test was therefore used to determine significance between vehicle and test groups in these data sets. For
Results
Establishing a Coculture Model of In Vitro Ischemia
To facilitate biosensor recording of neurotransmitter release
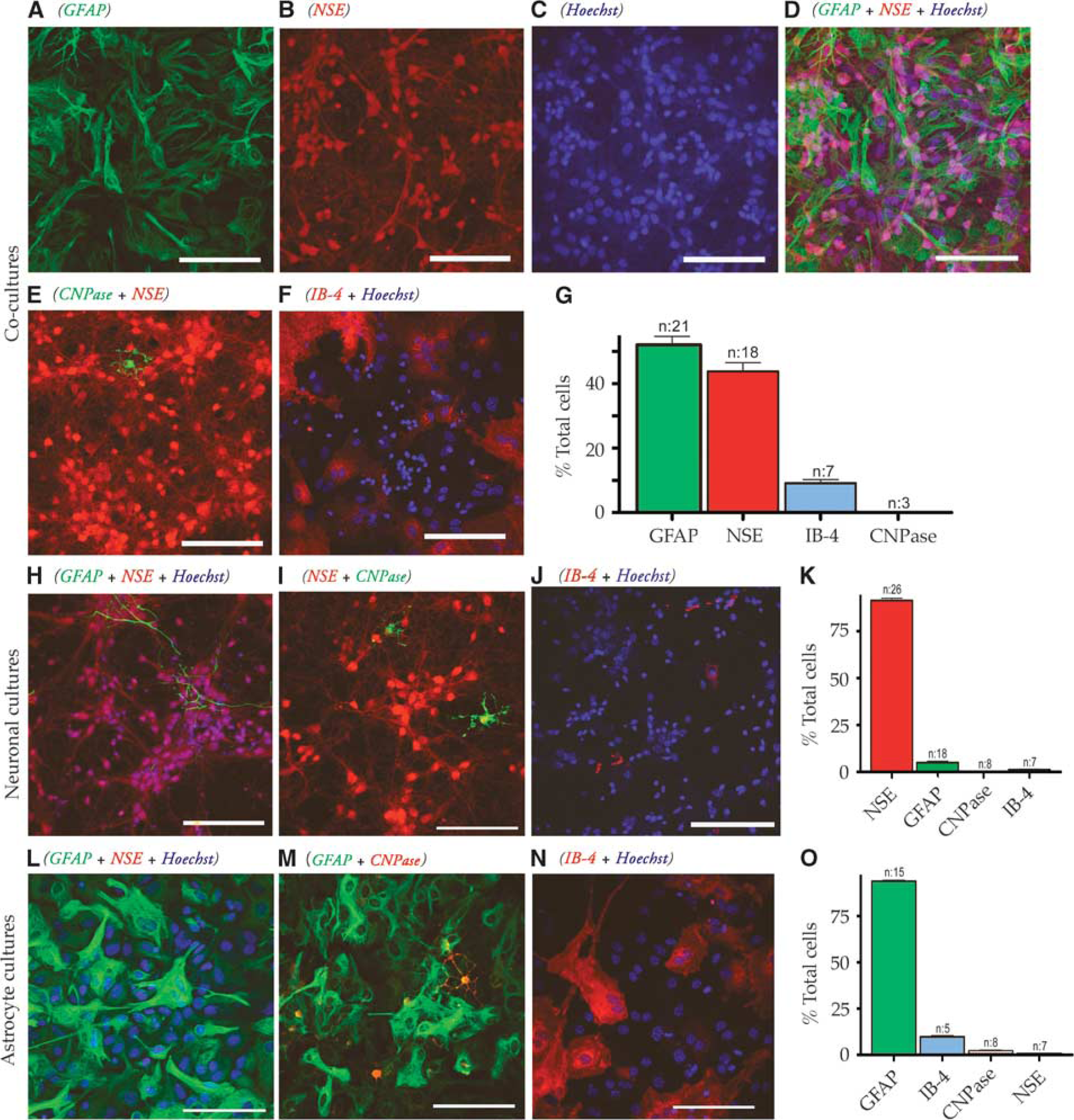
Characterization of high-density cultures (HDCs). (
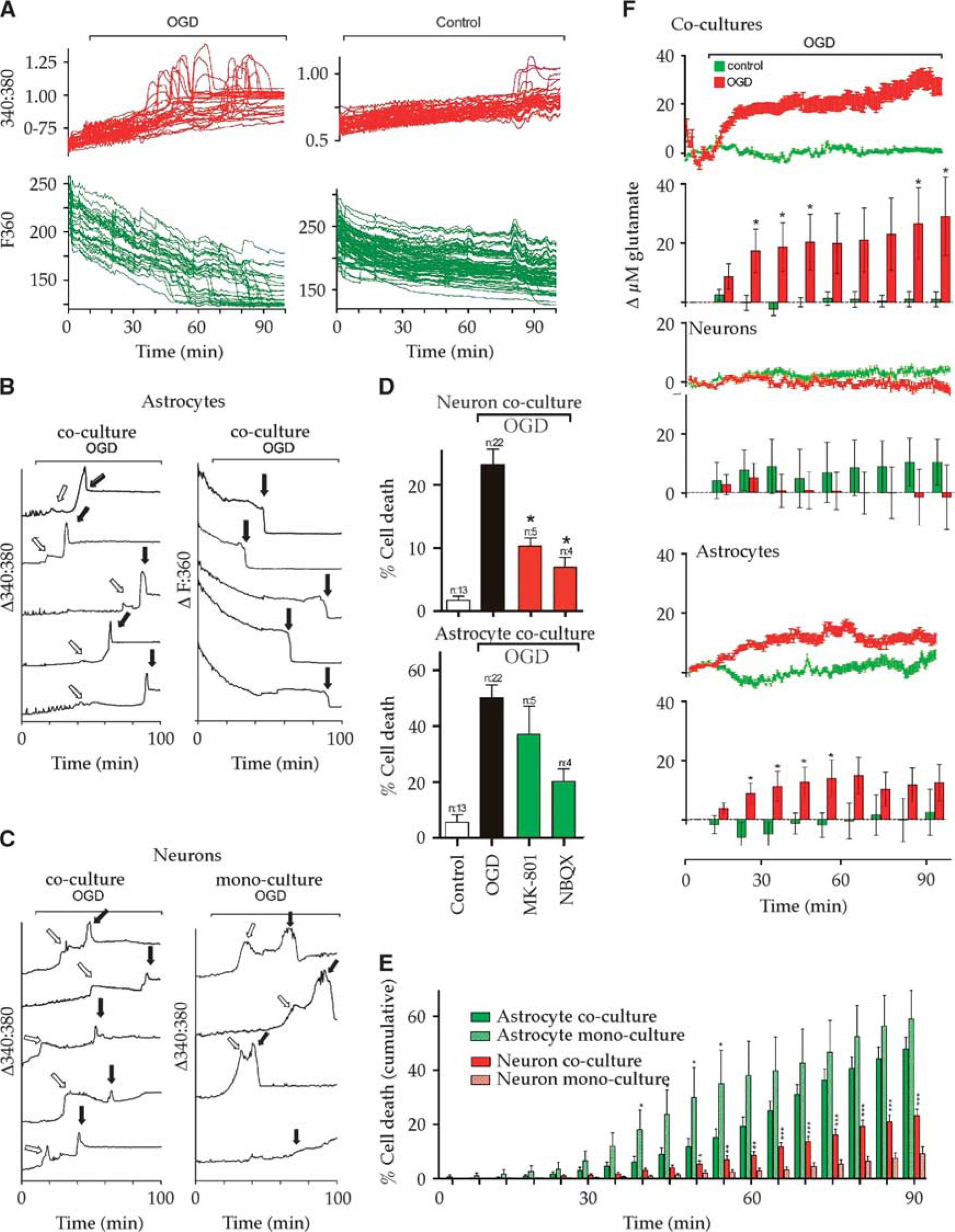
Cellular interactions during ischemia in high-density cultures (HDCs). (
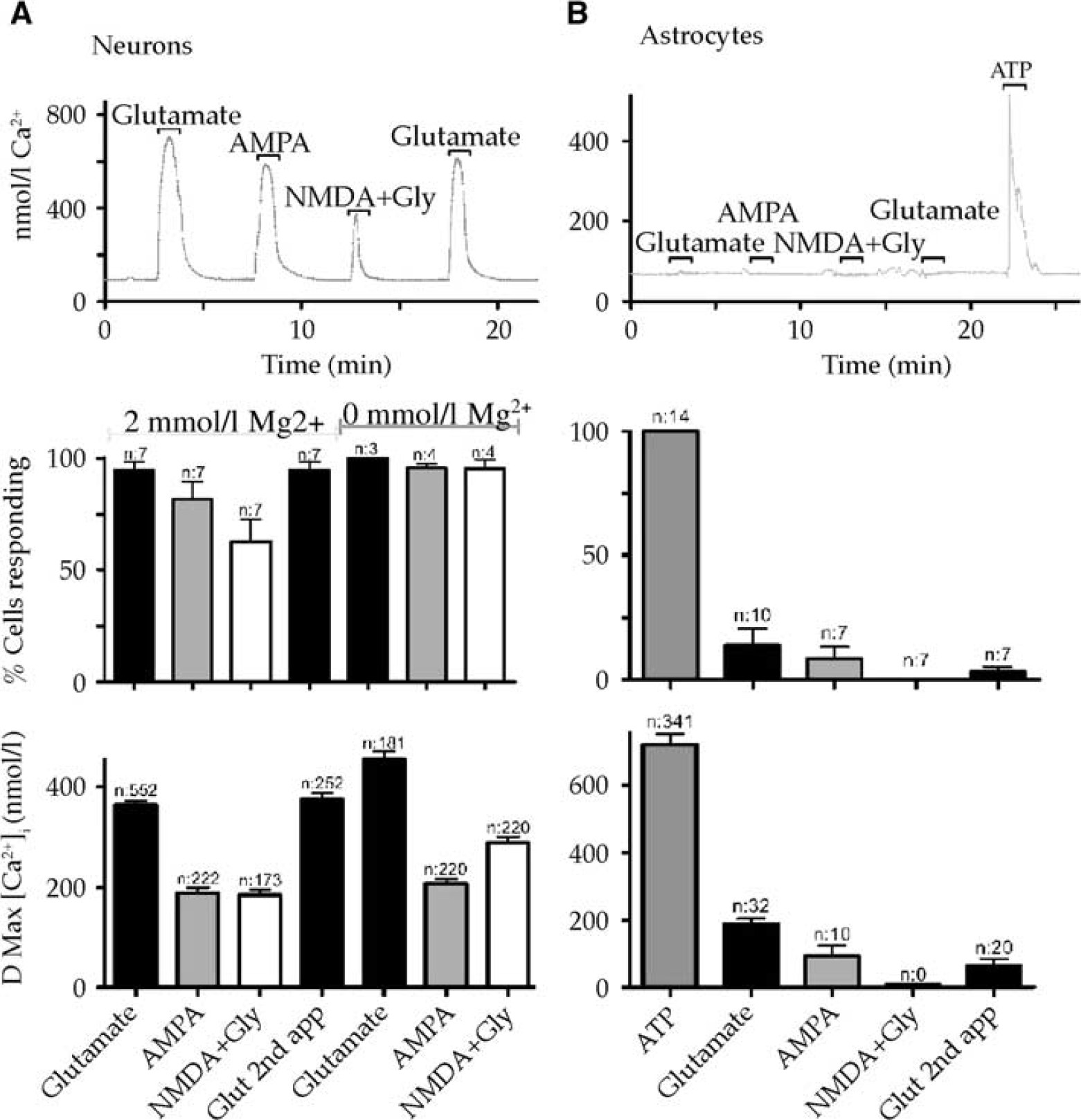
Intracellular Ca2+ ([Ca2+]i) changes evoked in neurons (
Dose-Dependent Effects of Memantine after In Vitro Ischemia
High concentrations of memantine (100 to 300 μmol/l) produced toxicity in normal-density cocultured cells when perfused for 40 to 90 minutes, affecting both astrocytes and neurons (Figures 4A and 4B). Although toxic effects of NMDA receptor antagonists have been noted previously (see Longuemare
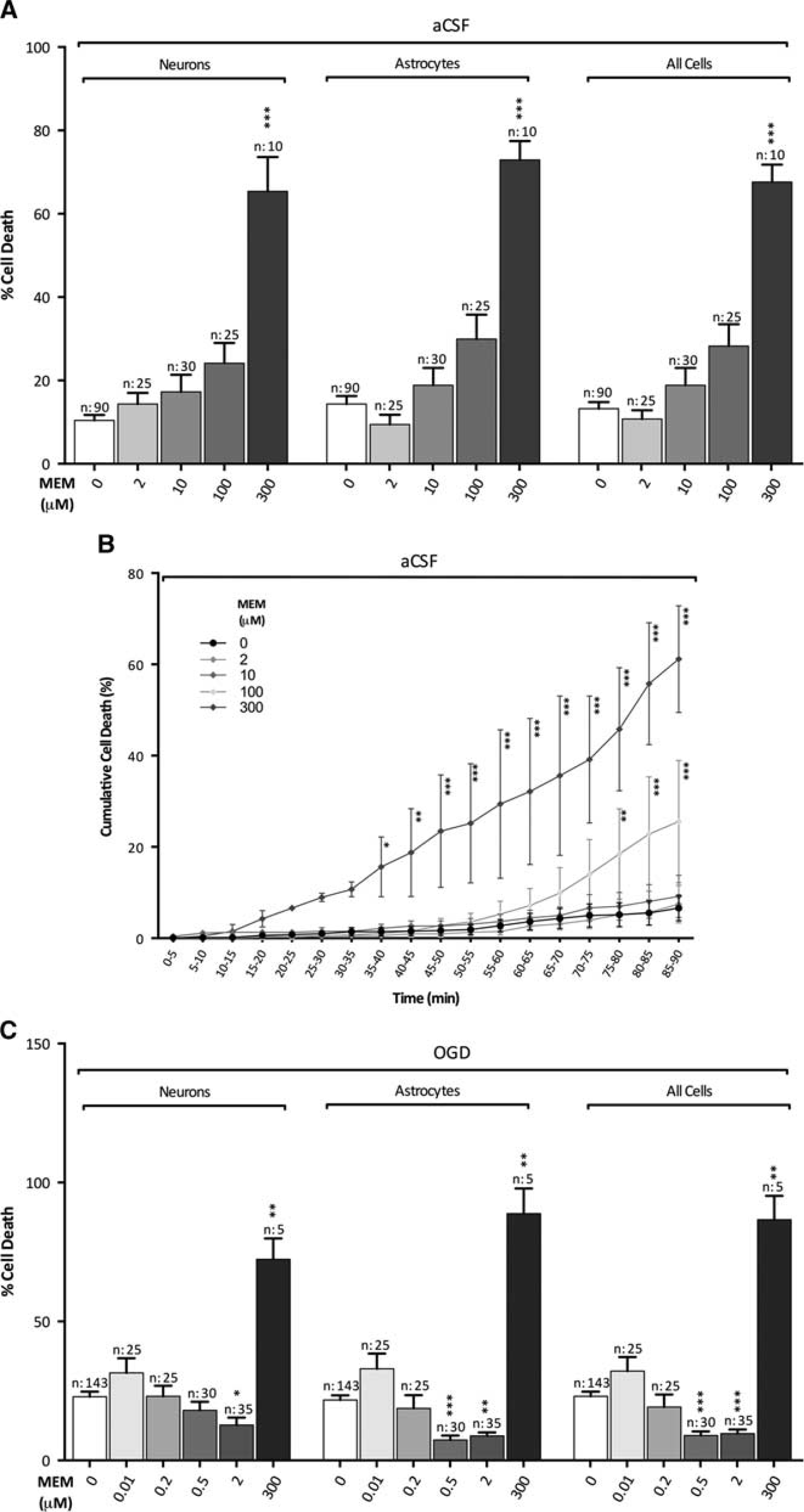
Protection and toxicity of memantine in normal-density cultures. (
Establishing the Dose-Dependent Effects of Memantine after In Vivo Cerebral Ischemia
Preinfusion for 24 hours with low-dose memantine reduced whole lesion volumes but significance levels were not achieved using the appropriate nonparametric comparison (Figure 5A). Significant reduction in both striatal and striatocortical lesions (examples shown in Figures 5B and 5C) were found in the 0.2-mg/kg memantine infusion group when the two lesion types were separated (Figure 5C). Significance was tested using a parametric analysis of variance
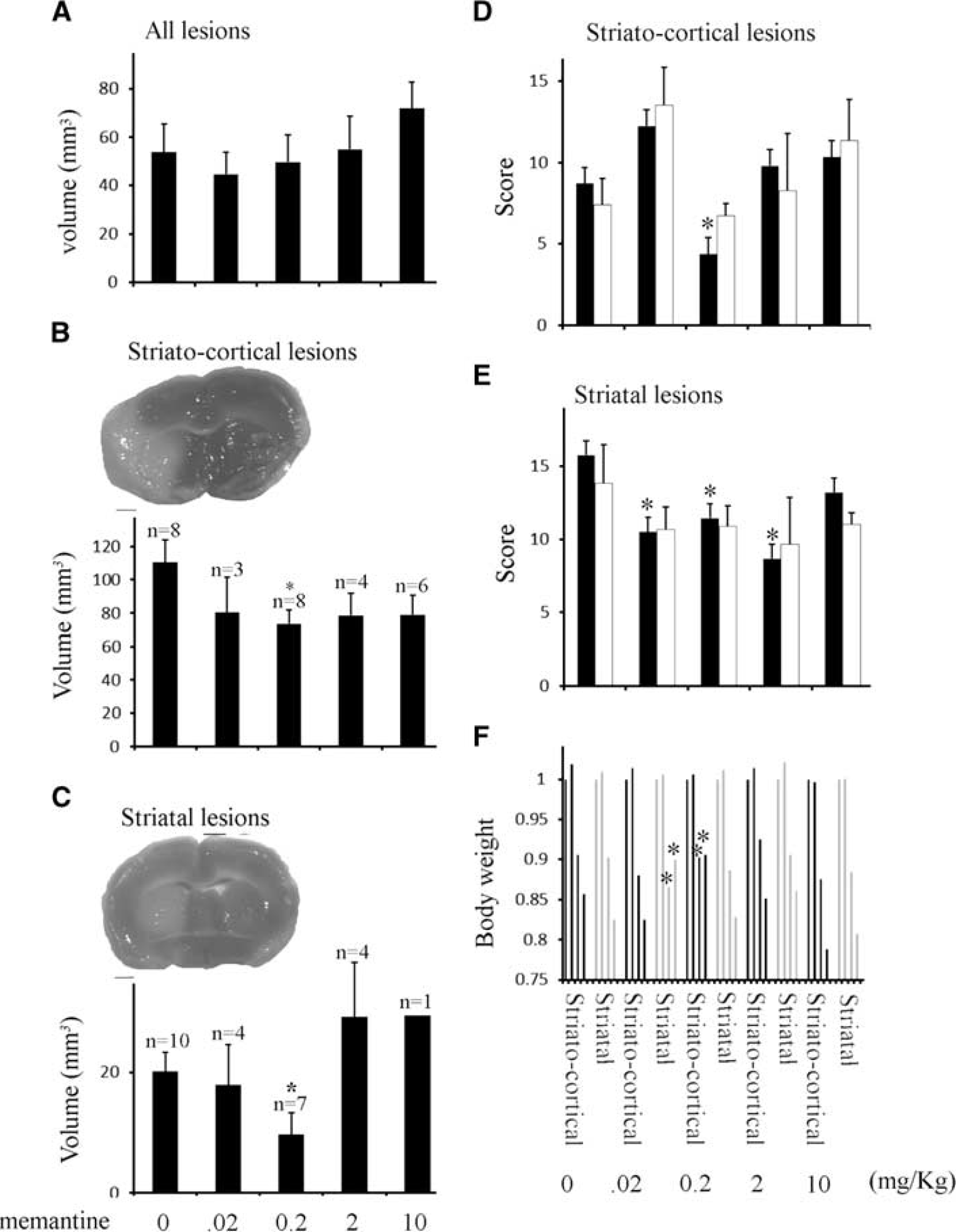
Pretreatment with low doses of memantine are highly protective in both striatal and striatocortical lesions. (
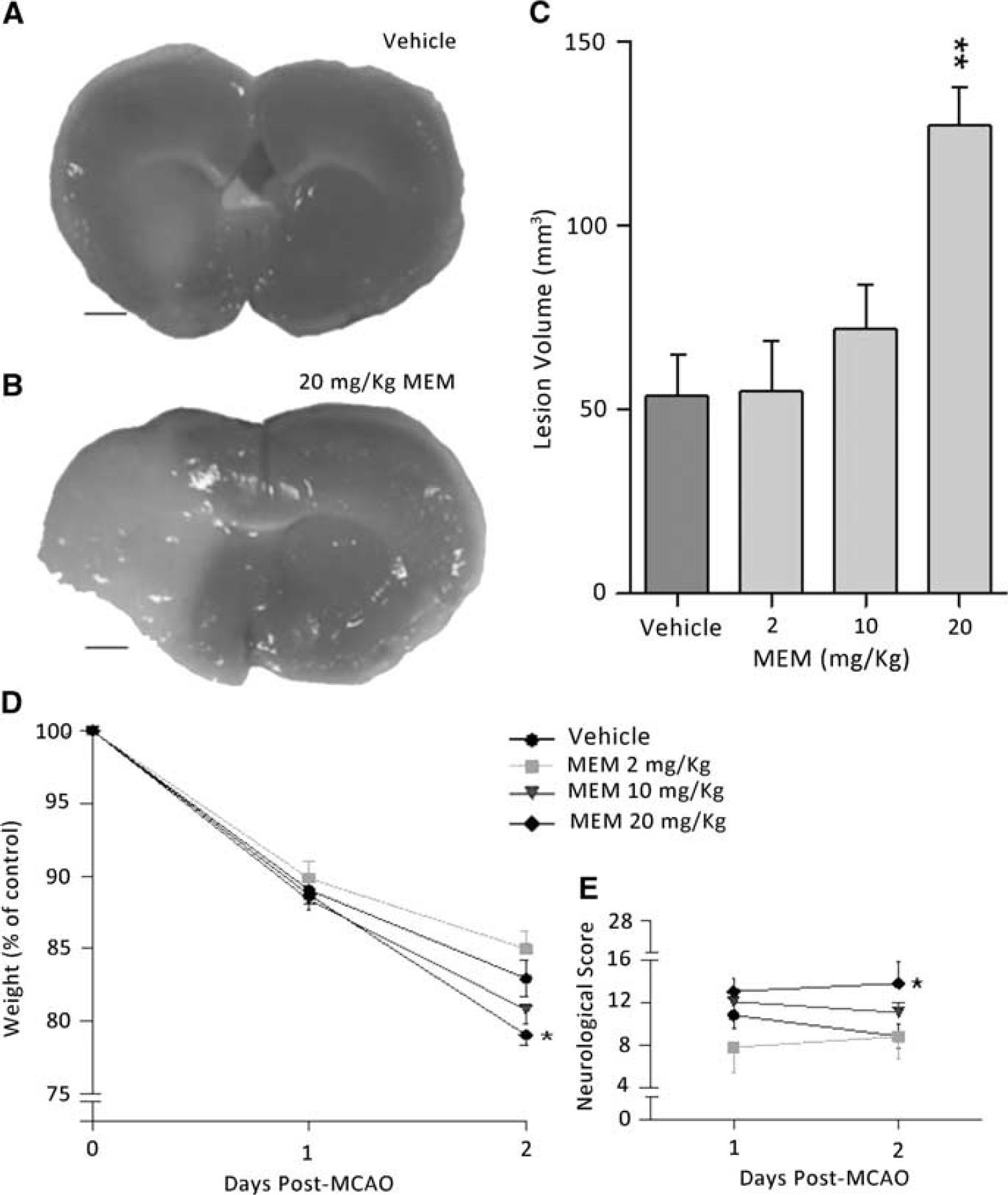
Potentiation of injury at high memantine doses. (
Discussion
In the current study, memantine exerted a dose-dependent effect in both the degree of protection provided against ischemic injury and the degree of toxicity directed against neural cells. A broad range of concentrations/doses were tested, revealing significant protection at levels several orders of magnitude lower than previously tested in
To investigate the effect of memantine
Although higher concentrations of memantine potentiated cell death during 90 minutes of OGD in coculture, significant protection was seen in the presence of lower concentrations (0.5 to 2 μmol/l). The current well-tolerated therapeutic dose for Alzheimer's treatment is 5 to 10 mg/day, with 5 mg/day producing a CSF concentration in patients of ∼0.05 μmol/l,
38
although the effective concentration at the receptor mouth
Antiexcitotoxic drugs were first identified as potential stroke therapies over 30 years ago, but have failed to translate into clinical practice because of dose-dependent complications.
39
Memantine is a moderate affinity noncompetitive NMDA receptor antagonist, binding directly within the pore of the channel in its open configuration.35, 36, 40 Although it displays fast on/off kinetics, it also shows partial trapping on agonist removal.
41
These properties make memantine effective in models of stroke at high (20 mg/kg) concentrations when administered in the acute treatment window;42, 43 20 mg/kg/day infusion produces a CSF concentration in rodents of 0.5 to 1 μmol/l,
18
an order of magnitude higher than that reported in patients receiving a standard 5 mg/day regime.
38
Although there are no data indicating rodent-CSF levels at lower infusion doses, the 0.2 mg/kg/day dose found to be protective in the current experiments is two orders of magnitude lower than the standard 20 mg/kg/day protocol use elsewhere and is likely to correlate to the 0.05 μmol/l levels reported in patients taking 5 to 10 mg/day. That protection was found at doses lower than previously reported and which are likely to correlate to well-tolerated current clinical practice suggests that prophylactic treatment for patients at risk of stroke is feasible. A discrepancy between the low protective concentrations found
In the current study, as others have also reported, we observed two distinct patterns of ischemic injury; small striatal lesions and large corticostriatal lesions. Why a standard protocol of focal ischemia should produce one of two distinct injury patterns may relate to variability in vascular anatomy or genetic makeup, or may arise from a normal distribution in the vascular field size producing lesions that encroach from the striatum into the cortex in a proportion of cases, 46 where a separate injury cascade may spread the lesion in an all or none manner throughout a large cortical area. Initiation of spreading depression, for example, may act to propagate injury from a relatively small cortical border zone through the cortical hemisphere.47, 48
This study has showed both the protective and toxic effects of memantine after
Footnotes
The authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.