
Abstract
The phosphocreatine/creatine kinase (PCr/CK) system in the brain is defined by the expression of two CK isozymes: the cytosolic brain-type CK (BCK) and the ubiquitous mitochondrial CK (uMtCK). The system plays an important role in supporting cellular energy metabolism by buffering adenosine triphosphate (ATP) consumption and improving the flux of high-energy phosphoryls around the cell. This system is well defined in muscle tissue, but there have been few detailed studies of this system in the brain, especially in humans. Creatine is known to be important for neurologic function, and its loss from the brain during development can lead to mental retardation. This study provides the first detailed immunohistochemical study of the expression pattern of BCK and uMtCK in the human brain. A strikingly dissociated pattern of expression was found: uMtCK was found to be ubiquitously and exclusively expressed in neuronal populations, whereas BCK was dominantly expressed in astrocytes, with a low and selective expression in neurons. This pattern indicates that the two CK isozymes are not widely coexpressed in the human brain, but rather are selectively expressed depending on the cell type. These results suggest that the brain cells may use only certain properties of the PCr/CK system depending on their energetic requirements.
INTRODUCTION
Creatine is a guanidino organic acid whose primary purpose is to receive the terminal phosphate group from adenosine triphosphate (ATP). 1 This occurs via a reversible phosphotransfer reaction catalyzed by creatine kinase (CK): creatine + ATP → phosphocreatine (PCr) + adenosine diphosphate (ADP). Two isozymes of CK can be expressed within a cell: a cytosolic form and a mitochondrial form. 1 In the brain, the isoforms of each type expressed are brain-type CK (BCK) as the cytosolic isozyme, and ubiquitous mitochondrial CK (uMtCK), located on the outer face of the inner mitochondrial membrane. 1 Altogether, the function derived from the CK reaction is referred to as the PCr/CK system.
The CK isozymes are expressed widely in tissues throughout the body; however, they tend to be enriched in tissues that have high energy requirements, such as muscle fibers and the brain. 2 Derived from research in skeletal muscle fibers, there are three main functions ascribed to the PCr/CK system: (1) temporal buffering of ATP where pools of PCr are depleted to maintain ATP levels during periods of energy consumption; 1 (2) spatial buffering where the diffusion of creatine and PCr between energy sources and sinks acts as a form of facilitated diffusion of ATP by proxy; 3 and (3) through the action of the mitochondrial CK, improving the efficiency of oxidative phosphorylation. 1 Incorporating these properties is the phosphocreatine circuit model, where CK isozymes are specifically associated with ATP sources and sinks, allowing the cell to direct the flux of energy toward designated sites. 1
These functions, though traditionally an ascribed part of muscle physiology, are equally applicable to the brain. Indeed, the brain appears to show an enhanced sensitivity to disturbances in the PCr/CK system compared with muscle fibers. The loss of creatine from the brain during development and early childhood, as a result of mutations in the genes responsible for creatine synthesis or transport, leads to a severe childhood mental retardation syndrome and a surprisingly minimal effect on muscle function. 4 Moreover, in neurodegenerative diseases such as Huntington's disease (HD), where energetic dysfunction is a common observation, creatine shows neuroprotective effects. 5 In HD, BCK has also been observed to be reduced, though it is not known whether this is an underlying cause of cell death or a result of disease processes. 6
It is still commonly assumed that the PCr/CK system operates in all cells of the brain. However, there is some evidence that BCK and uMtCK may have quite different cellular distributions. 7 Such dissociated expression patterns would require a reconsideration of how the PCr/CK system actually operates in different brain cells. In the human brain, the distribution of BCK and uMtCK has not been well described. To address these issues, we have conducted a detailed immunohistochemical analysis of BCK and uMtCK expression in the human brain. We show that BCK and uMtCK are indeed selectively expressed, particularly between astrocytes and neurons, and that this expression pattern can be rationalized by considering the specific phenotype of energy metabolism that each cell expresses. The results suggest that the PCr/CK system is used in ways by each cell depending on the demands placed on their energy system that their specific function requires.
MATERIALS AND METHODS
Acquisition and Processing of Human Brain Tissue
Fixed-frozen, post-mortem human brain tissue was acquired from the Neurological Foundation of New Zealand Human Brain Bank, located in the Department of Anatomy with Radiology and Centre for Brain Research at The University of Auckland, New Zealand. The brain tissue is collected by donor program and is used in accordance with The University of Auckland Human Participants Ethics Committee (2008/279). The right hemisphere is fixed by perfusion with 15% formalin solution before being cryoprotected in sucrose and then frozen (fixed-frozen); the left hemisphere is frozen without fixation (fresh-frozen). Details of these procedures has been published previously. 8
A total of 12 neurologically normal human brains were used for this study. The average age at death was 63 years and the average postmortem time was 16 hours. A full list of cases and regions they were used for can be found in Table 1. For 3,3'-diaminobenzidine tetrahydrochloride (DAB)-peroxidase and immunofluorescent labeling, each case as detailed in Table 1 was stained at least twice in separate experiments and compared for consistency.
List of cases
F, female; M, male.
3,3'-Diaminobenzidine Tetrahydrochloride–Peroxidase Immunohistochemistry
Coronal sections (50 μm) through the spinal cord (at the C1-2 level), brainstem, cerebellum, thalamus, basal ganglia, hippocampus, and motor cortex were stained for either BCK or uMtCK by the DAB-peroxidase method as previously described. 8 Briefly, sections were incubated in a blocking solution of 50% methanol and 1% H2O2 for 20 minutes, either the goat anti-BCK or goat anti-uMtCK (see ‘Antibodies’) for 72 hours, a biotinylated rabbit anti-goat IgG secondary antibody (B7024; 1:2,000; Sigma-Aldrich, St Louis, MO, USA) for 24 hours, and then ExtrAvidin (E2886; 1:1,000; Sigma-Aldrich) for 4 hours. All antibodies or ExtrAvidin were diluted in phosphate-buffered saline (PBS) with 1% rabbit serum (01-6101; Invitrogen, Camarillo, CA, USA), 0.2% Triton X-100, and 0.04% merthiolate (‘immunobuffer’). Incubation steps were preceded by 3 × 10-minute washes in PBS with 0.2% Triton X-100 (PBST). Labeling was developed in a solution of 0.1 mol/L phosphate buffer, 0.05% DAB, 0.01% H2O2, and 0.04% nickel ammonium sulfate. Sections were mounted onto glass slides, dehydrated through a graded series of alcohols, and then cleared in xylene. Photomicrographs were taken with a Leica DMRB microscope or a Leica MZ6 dissecting microscope (Wetzlar, Germany) with a digital camera attached. Images were contrast-adjusted in Photoshop CS4 (Adobe Systems Software, San Jose, CA, USA) to improve clarity.
Immunofluorescent Histochemistry
Double or triple labeling of sections by immunofluorescent histochemistry follows a similar protocol to DAB-peroxidase staining. 8 In brief, sections were incubated in a solution of 90% acetone and 10% methanol for 20 minutes, a solution of the primary antibodies (see list below) diluted in immunobuffer (with 1% normal donkey serum (D9663; Sigma-Aldrich)) for 72 hours, and then a solution of the Alexa Fluor-conjugated secondary antibodies for 24 hours diluted in immunobuffer (Molecular Probes, Eugene, OR, USA). All steps were preceded by washing in PBST. Sections are then incubated in a solution of 70% ethanol with 0.1% Sudan black for 15 minutes, rinsed 2 × 2 minutes in PBST, and then mounted onto glass slides. Sections were then coverslipped with SlowFade Gold (S36936; Invitrogen) and then sealed with nail varnish. Slides were imaged using a Zeiss 710 confocal laser scanning microscope (Carl-Zeiss, Jena, Germany).
Antibodies
For DAB-peroxidase labeling, either a goat anti-BCK (sc-15157; 1:1,000; Santa Cruz Biotechnology, CA, USA) or goat anti-uMtCK (sc-15165; 1:200; Santa-Cruz) was used. For fluorescent double labeling of uMtCK and BCK together, a mouse anti-BCK (sc-373686; 1:200; Santa-Cruz) was used with the goat anti-uMtCK (1:200). Several other antibodies were used to label proteins that serve as anatomic markers for double or triple labeling. Those used were: rabbit anti-glial fibrillary acidic protein (GFAP; Z0334; 1:1,000; Dako Corporation, Carpinteria, CA, USA); rabbit anti-neuronal nuclei (NeuN; ABN78; 1:200; Merck-Millipore, Billerica, MA, USA), or mouse anti-NeuN (MAB377; 1:200; Millipore); mouse anti-parvalbumin (235; 1:5,000; SWANT, Bellinzona, Switzerland) or guinea pig anti-parvalbumin (195 004; 1:1,000; Synaptic Systems, Goettingen, Germany); rabbit anti-synaptophysin (M0776; Dako); guinea pig anti-vesicular glutamate transporter 2 (VGluT2; VGluT2-GP-Af670-1; 1:100; Frontier Institute, Hokkaido, Japan); rabbit anti-vesicular γ-butyric acid transporter (VGAT; 131-003; 1:1,000; Synaptic Systems); mouse anti-calbindin (300; 1:500; SWANT); mouse anti-SMI32 (SMI32R; 1:500, Sternberger Monoclonals Inc., Convance, Princeton, NJ, USA); mouse anti-choline acetyltransferase (ChAT; MAB5270; 1:200; Millipore), rabbit anti-Olig2 (AB9610; 1:250; Millipore), and mouse antimicrotubule-associated protein 2 (MAP2; M9442; 1:200; Sigma-Aldrich).
For fluorescent double labeling, Alexa Fluor 488- and 594-conjugated anti-IgG antibodies were used. For fluorescent triple labeling, an additional Alexa Fluor 647-conjugated anti-IgG antibodies was used. A full list of these antibodies is provided here (all are donkey host antibodies used at 1:400 from Invitrogen, unless stated otherwise): anti-goat IgG Alexa 488 (A11055) or Alexa 594 (A11058); anti-mouse IgG Alexa 488 (A21202), Alexa 594 (A21203), or Alexa 647 (A31571); anti rabbit IgG Alexa 488 (A212106), Alexa 594 (A21207), or Alexa 647 (A31573); and anti-guinea pig IgG Alexa 488 (Jackson ImmunoResearch, West Grove, PA, USA, 706-545-148; 1:200).
Controls for Immunohistochemistry
No-primary controls, where the primary antibody is omitted from the immunobuffer, were conducted on each case, for each region, and for each secondary antibody (biotinylated and fluorescent) to assess the degree of nonspecific labeling because of the secondary antibodies. The specificity of the antibodies against the CK isozymes were assessed by western blotting (see below) and preabsorption of the antibodies with the antigenic peptide used to produce the antibodies. In this case, 25 μg of blocking peptide was incubated with 0.5 μg of antibody for 1 hour at 4°C before being diluted and used in either western blotting or immunohistochemical applications as described.
Western Blotting
Antibodies against the CK isozymes were blotted against human brain protein extracted from the motor cortex of case H120 (Table 1). Western blotting was performed using the XCell Surelock Minicell and XCell II blot module systems (Invitrogen) according to the manufacturer's instructions. This procedure has been detailed previously. 9 All primary antibodies were used at 1:200.
RESULTS
The expression pattern of BCK and uMtCK was assessed in major regions of the human brain. A qualitative assessment of their staining intensity was conducted on DAB-peroxidase-stained sections. The results are summarized in Table 2 and further detailed below.
Regional distribution and immunoreactivity of brain-type creatine kinase (BCK) and ubiquitous mitochondrial creatine kinase (uMtCK) in the human brain
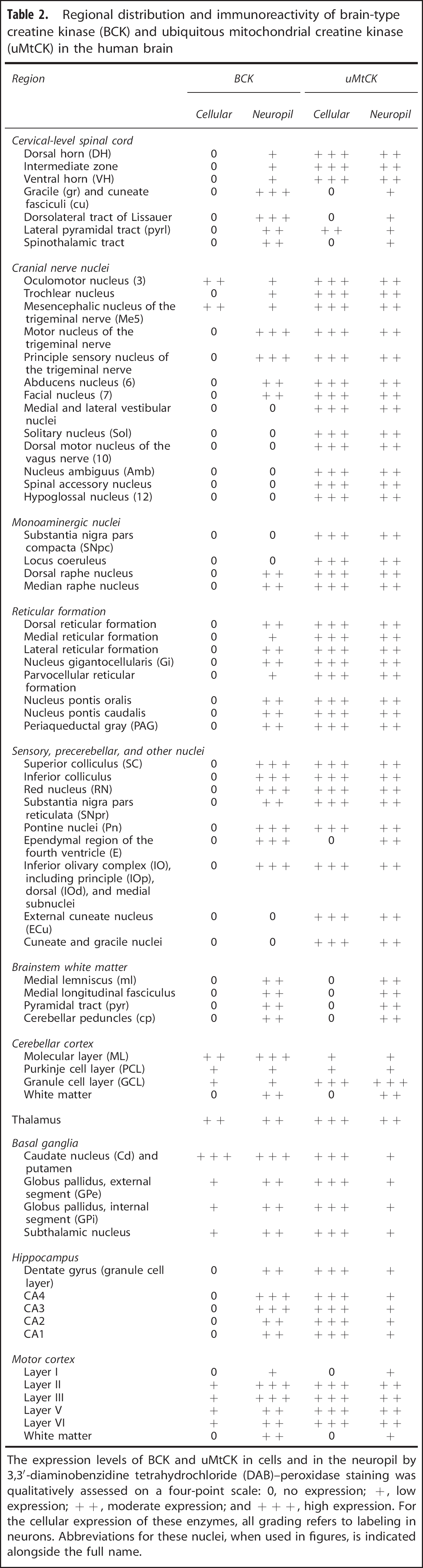
The expression levels of BCK and uMtCK in cells and in the neuropil by 3,3'-diaminobenzidine tetrahydrochloride (DAB)-peroxidase staining was qualitatively assessed on a four-point scale: 0, no expression; +, low expression; + +, moderate expression; and + + +, high expression. For the cellular expression of these enzymes, all grading refers to labeling in neurons. Abbreviations for these nuclei, when used in figures, is indicated alongside the full name.
Western Blotting
Western blotting of the antibodies against the CK isozymes (goat anti-BCK, goat-uMtCK, and mouse anti-BCK) using human brain homogenate from the motor cortex labels a single band at 43 kDa (Figure 1B). This corresponds to the molecular mass of the CK monomer for both isozymes. 10 Preabsorption of the goat anti-BCK antibody with its blocking peptide, or goat anti-uMtCK with its blocking peptide, eliminates this 43 kDa band on a western blot (Figure 1B), and any specific staining by immunohistochemistry (Figure 1C).
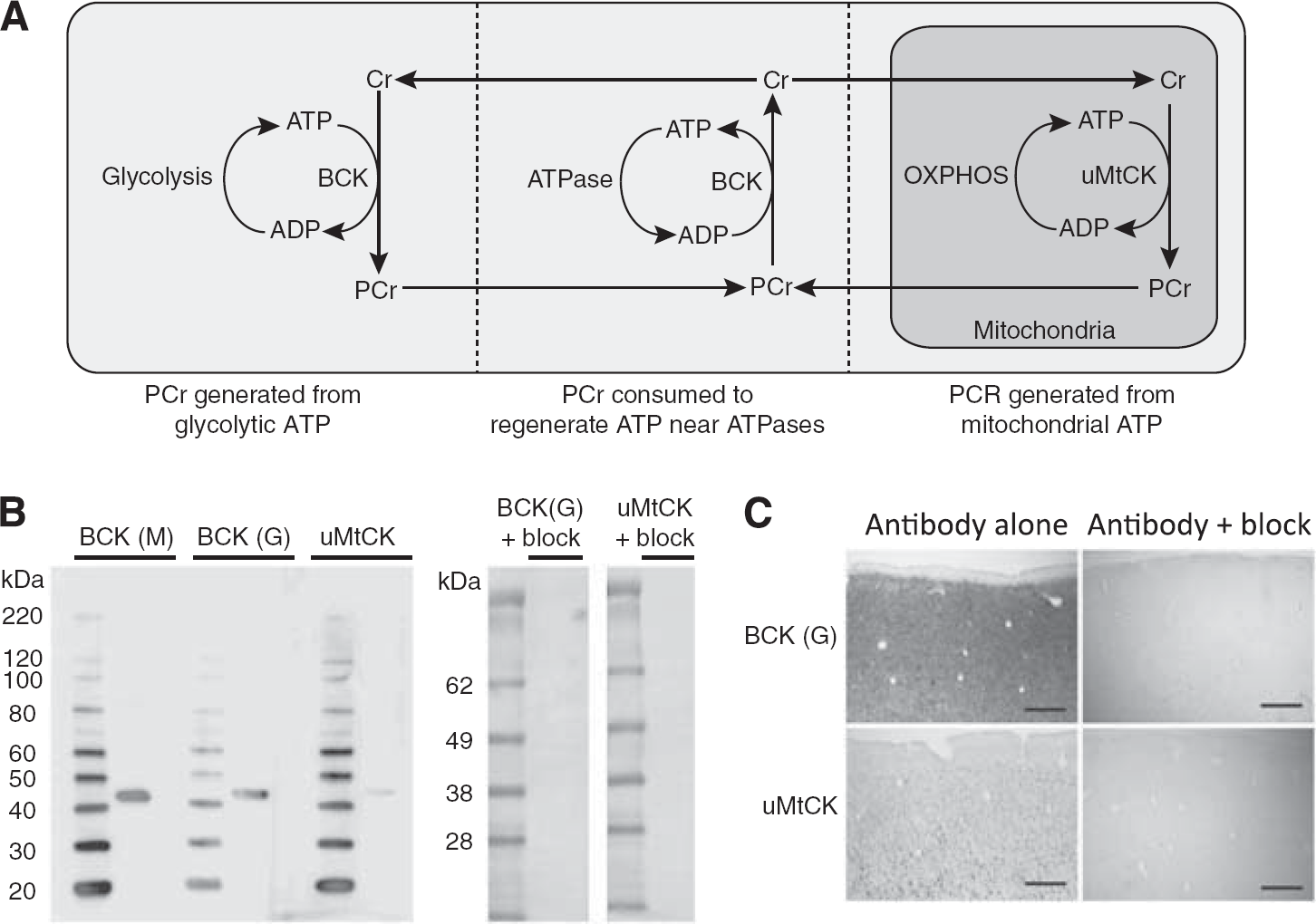
Model of creatine function and western blotting controls. (
Consistency of Staining Between Cases
In total, 12 individual cases were examined for BCK and uMtCK expression (Table 1). These cases showed only minor variability in their global level of reactivity, but showed a consistent pattern of staining, with similar relative degrees of reactivity, between cases.
Spinal Cord and Brainstem
Throughout the C1-2 levels of the spinal cord and in the brainstem, in all cases examined, the pattern of BCK or uMtCK expression shows a consistent distribution. Figures 2A to 2H show macroscopic images of BCK and uMtCK reactivity in coronal sections through the spinal cord, medulla, pons, and midbrain. Ubiquitous MtCK shows strong and consistent staining in all of the nuclear regions where the cell bodies of neurons reside. White matter tracts also show some reactivity, although at a lower level. In contrast, BCK is strongly reactive in selective regions corresponding to white matter tracts. This is particularly apparent in the spinal cord section where BCK reactivity is confined to the superficial aspects, notably in the gracile and cuneate fasciculi, dorsolateral fasciculus, and part of the lateral pyramidal tract (Figure 2A). The gray horns, including the dorsal horn and ventral horn, are unstained for BCK but strongly reactive for uMtCK.
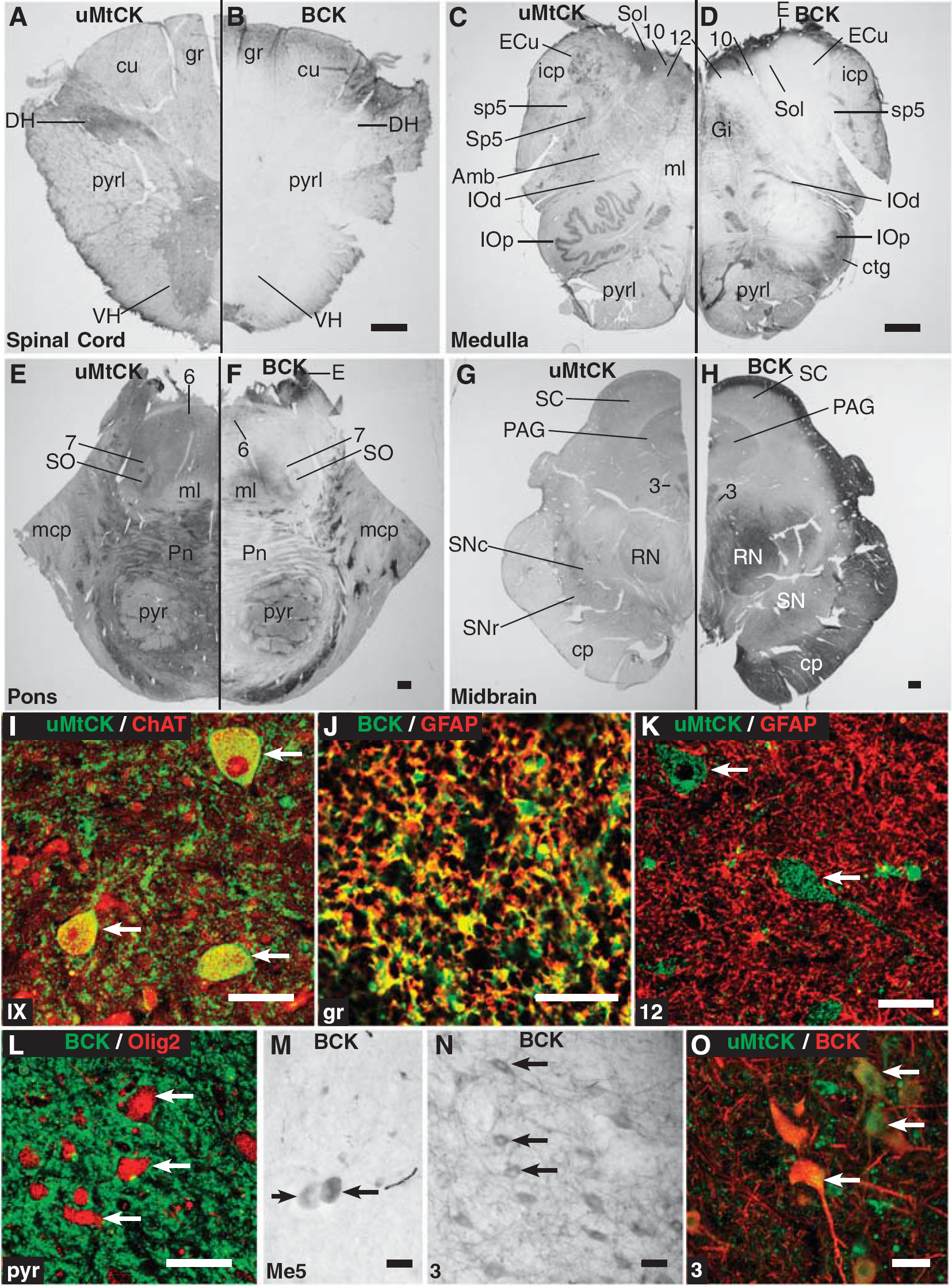
Immunohistochemical expression of brain-type creatine kinase (BCK) and ubiquitous mitochondrial creatine kinase (uMtCK) in the spinal cord and brainstem. (
The macroscopic pattern of BCK expression in the spinal cord (see Figure 2B) does appear restricted in a manner that may suggest a methodological issue, such as poor antibody penetration. However, it is noted that uMtCK (Figures 2A and 2I) showed strong expression throughout the spinal cord. In addition, the three spinal cord cases (Table 1) all gave consistent labeling of BCK as in Figure 2B with both the goat anti-BCK and mouse anti-BCK antibodies (see Materials and Methods for details) with DAB-peroxidase staining.
In the ventral horn of the spinal cord, lamina IX, or the motor column, uMtCK is strongly expressed in (motor) neurons in the nucleus, particularly the cell body and proximal dendrites, where there is a punctate pattern of staining. This reactivity colocalizes with the motor neuron marker ChAT (Figure 2I). In the neuropil, uMtCK is also strongly expressed as punctate bodies. In contrast, BCK shows scant expression in the entirety of the gray horns (see Figure 2B).
In the white matter regions where BCK is strongly expressed, BCK is almost completely expressed in astrocytes based on its colocalization with GFAP (Figure 2J). Unlike BCK, uMtCK does not colocalize with GFAP, as shown in Figure 2K in the hypoglossal nucleus where uMtCK-reactive puncta can be observed in the cell bodies and fibers of motor neurons similar to that in the ventral horn of the spinal cord (Figure 2I). Furthermore, BCK reactivity does not colocalize with the oligodendrocyte marker Olig2 (Figure 2L).
Only two nuclear regions of the brainstem showed evidence of BCK reactivity in neurons. The first is the mesencephalic nucleus of the trigeminal nerve where BCK expression appears in the pseudounipolar neuronal soma in the characteristic ovoid shape (Figure 2M). The second is the nucleus of the oculomotor nerve (Figure 2N; can also be seen in Figure 2H). In this nucleus, BCK is expressed in the soma of the motor neurons as well as dendrites within the nucleus. Ubiquitous MtCK is also expressed in these same neurons (Figure 2O).
Hence, in the spinal cord and brainstem, BCK is expressed primarily in astrocytes (and not oligodendrocytes), whereas uMtCK is expressed primarily in neurons and not in astrocytes. Only two regions showed coexpression of uMtCK and BCK—the mesencephalic nucleus of the trigeminal nerve and the nucleus of the oculomotor nerve.
Cerebellar Cortex
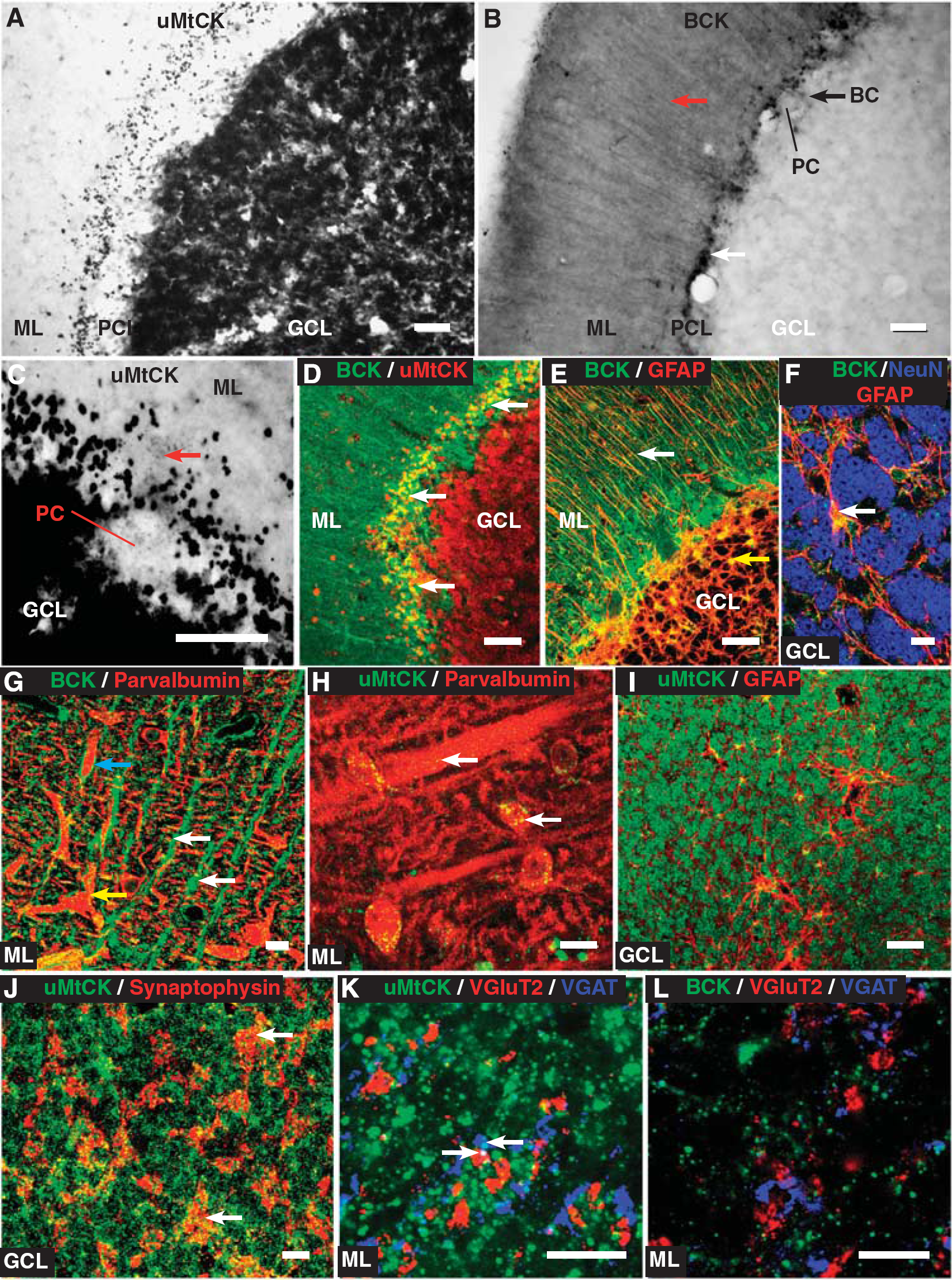
The DAB-peroxidase staining of the CK isozymes in the cerebellar cortex shows a starkly contrasting pattern of expression (Figures 3A and 3B). Ubiquitous MtCK is intensely expressed in the granule cell layer and in the densely packed granule cells, whereas it shows much lower expression in the Purkinje cell layer and molecular layer. In contrast, BCK is expressed strongly in the molecular layer, particularly in thin vertically oriented fibers, with much lower expression in the Purkinje cell layer and granule cell layer. However, both BCK and uMtCK show dark punctate staining in the interface between the Purkinje cell layer and molecular layer (see Figures 3A and 3B) that likely corresponds to the cell bodies of basket cells.
Expression of brain-type creatine kinase (BCK) and ubiquitous mitochondrial creatine kinase (uMtCK) in the cerebellum. (
The BCK is expressed weakly in the cell bodies of Purkinje cells, the main output neuron of the cerebellar cortex (Figure 3B). Ubiquitous MtCK is also found weakly expressed and, at higher magnification, it shows small puncta within the cell body of Purkinje cells (Figure 3C). Ubiquitous MtCK is highly expressed within granule cells in the granule cell layer, whereas in this layer, BCK has a reticular-like pattern of expression (Figure 3D). Colabeling with GFAP shows that this corresponds to BCK expression in astrocytes that surround granule cells (Figures 3E and 3F). The vertically oriented fibers expressing BCK in the molecular layer also colocalize with GFAP (Figure 3E) and are likely to be the processes of Bergmann glia. Colabeling between BCK and parvalbumin, a marker of Purkinje cell dendrites, in the molecular layer reveals how the Bergmann glial fibers are organized. In Figure 3G, the BCK-reactive vertically oriented fibers are clearly evident. However, many of these fibers also surround parvalbumin-reactive dendrites, which is a characteristic of Bergman glia. Moreover, in Figure 3G, weaker BCK reactivity in parvalbumin-reactive dendrites is also apparent. Colabeling between uMtCK and parvalbumin shows punctate labeling through parvalbumin-reactive dendrites and in the cell bodies of stellate cells (Figure 3H).
Although uMtCK is expressed in granule cells in the granule cell layer (see Figure 3A), it is not in the GFAP-expressing astrocytes that were shown to express BCK (Figure 3I; see also Figures 3E and 3F). In addition, uMtCK is also expressed in other parts of the tripartite synapse representing the inputs onto granule cells, known as the cerebellar glomerulus. Ubiquitous MtCK colocalizes with synaptophysin, a presynaptic marker that labels the inputs of the glomerulus (Figure 3J). When labeled individually, with VGluT2 for mossy fibers and VGAT for Golgi terminals, uMtCK colocalizes with both (Figure 3K), although weaker than in granule cells. In contrast, BCK does not colocalize with either VGluT2 or VGAT (Figure 3L).
Overall, uMtCK is expressed widely in neuronal elements in the cerebellar cortex, although most strongly in granule cells. It is expressed at or near synaptic structures in parvalbumin-expressing dendrites of Purkinje cells and in the neuronal components of the glomerulus. In contrast, BCK is enriched in astrocytes throughout the cerebellar cortex, but does show weaker expression in some neuronal components such as Purkinje cells and basket cells.
Thalamus, Globus Pallidus, and Subthalamic Nucleus
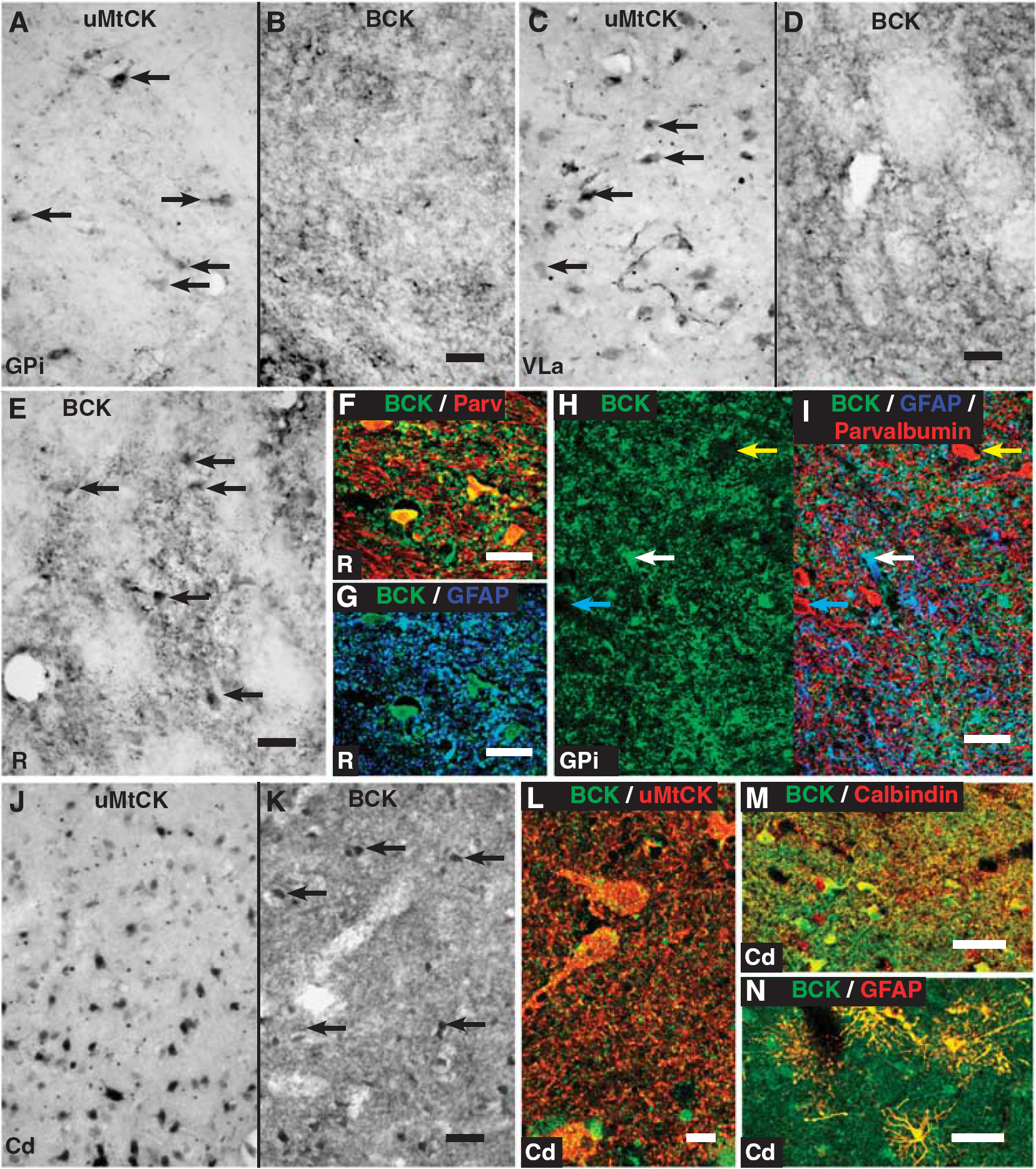
Both BCK and uMtCK individually show similar patterns of staining in the thalamus, globus pallidus, and subthalamic nucleus (Figure 4). Ubiquitous MtCK widely labels neuronal soma and a limited extent of proximal processes (Figures 4A and 4C). In contrast, BCK shows a reticular pattern of staining of fibers with small punctate cell bodies through all regions (Figures 4B, 4D, and 4E). Unlike in most of the thalamic nuclei, the globus pallidus, or the subthalamic nucleus, there is overt staining of neuronal cell bodies in the thalamic reticular nucleus, part of the ventral thalamus (Figure 4E). In this nucleus, BCK reactivity colocalizes with both parvalbumin-reactive neurons (Figure 4F) and GFAP-reactive astrocytes (Figure 4G). Moreover, fluorescent labeling of pallidal neurons with parvalbumin also reveals that in some neurons, there is a very low level of BCK reactivity, although the reticular pattern of BCK staining colocalizes extensively with GFAP (Figures 4H and 4I). Therefore, in these nuclei, uMtCK is widely expressed in neurons, whereas BCK has a limited expression in select neurons and a widespread expression in astrocytes.
Expression of brain-type creatine kinase (BCK) and ubiquitous mitochondrial creatine kinase (uMtCK) in the basal ganglia and thalamus. (
Striatum
The striatum shows a strong expression of both uMtCK and BCK. Like in other regions, uMtCK is widely expressed in neurons, especially their cell bodies and proximal processes (Figures 4J and 4K). Interestingly, BCK also shows expression in neurons as well as a dense fiber labeling in the neuropil (Figure 4K). At high magnification, uMtCK puncta colocalize within BCK-reactive fibers within the striatum (Figure 4L). These BCK-reactive neurons, and the fibrous staining in the neuropil, extensively colocalize with calbindin, a marker of medium spiny neurons (Figure 4M). These observations point to a strong coexpression of the CK isozymes within medium spiny neurons in the striatum. In addition, like in other regions of the brain, BCK also colocalizes with GFAP-reactive astrocytes (Figure 4N).
Hippocampus
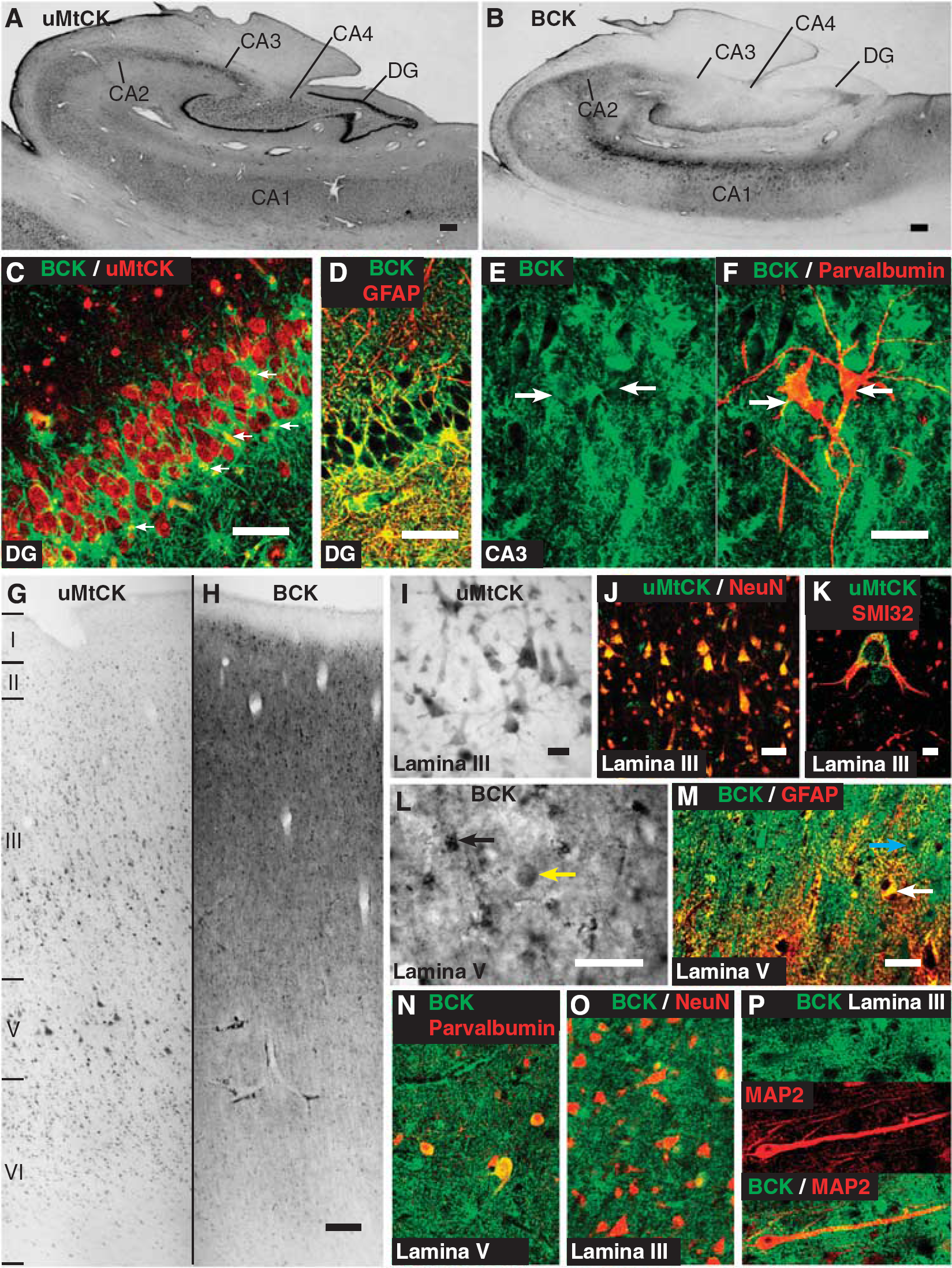
The DAB-peroxidase labeling of the CK isozymes in the hippocampus shows uMtCK expression in the granule cells of the dentate gyrus, and the neurons of all four fields of Ammon's horn (Figure 5A). In these regions, BCK also shows reactivity but without any overt labeling in neurons; rather, it is expressed in fibers in the neuropil and glial cells that have astrocytic morphologies (Figure 5B). The pattern of coexpression between the CK isozymes is highlighted with double labeling in the dentate gyrus (Figure 5C). Here, uMtCK is strongly expressed in the granule cell soma, tightly packed into the granule cell layer. In contrast, BCK is expressed in processes that surround the cell bodies (Figure 5C). There is no colocalization between the markers. The BCK reactivity in the dentate gyrus, and in most of the other regions of the hippocampus, largely colocalizes with GFAP (Figure 5D). Interestingly, BCK does colocalize with parvalbumin-reactive interneurons throughout the hippocampus (Figures 5E and 5F). Their morphology and parvalbumin-expression suggests that these neurons are either basket cells or chandelier cells.
Expression of brain-type creatine kinase (BCK) and ubiquitous mitochondrial creatine kinase (uMtCK) in the hippocampus and motor cortex. (
Motor Cortex
At the macroscopic level, BCK and uMtCK show remarkably different patterns of expression in the motor cortex (Figures 5G and 5H). Ubiquitous MtCK is widely expressed in neurons, with the large population of pyramidal neurons particularly evident. At the cellular level, uMtCK is expressed in the cell body and proximal processes (Figure 5I); it colocalizes extensively with the pan-neuronal marker NeuN (Figure 5J), and at a higher magnification, uMtCK forms small puncta throughout the cell body and proximal processes (Figure 5K).
In contrast, at the macroscopic level, BCK shows dense neuropil reactivity with small dark cell bodies scattered through layers II to VI (Figure 5L; also see Figure 5H). These cells, and much of the neuropil reactivity, colocalize with GFAP (Figure 5M). Interestingly, there appears to be little of the cortical gray matter that lacks any BCK reactivity, which may be expected if it were not expressed in some cells, such as neurons. Large ovoid cell bodies, with weaker staining than in astrocytes, are prevalent in the deep part of layer III and layer V (see Figure 5L). These cells colocalize with parvalbumin (Figure 5N). This, along with their morphology and localization, suggests that they are basket cells. More generally, a number of other neurons, often with pyramidal morphologies, show BCK reactivity at lower intensities than the surrounding neuropil staining (see Figure 5M). The BCK colocalizes weakly in NeuN-positive soma (Figure 5O). This weak BCK neuronal expression also extends into the larger primary dendrites of neurons as can be seen between BCK and MAP2 colocalization (Figure 5P).
Overall, uMtCK is expressed in neurons throughout the motor cortex, whereas BCK is expressed mainly in astrocytes, moderately in basket cells, and weakly in other neurons including pyramidal neurons.
DISCUSSION
The Anatomy of the PCr/CK System in the Brain
This detailed study has characterized the expression patterns of the two CK isozymes in the human brain. It complements previous animal studies,7,11 which are in good agreement, and extends our understanding of the nature of the PCr/CK system in the human brain. Principally, it was found that the two isozymes have opposing expression patterns: uMtCK is expressed exclusively in neurons, and BCK in astrocytes with weaker selective expression in neurons. With anatomic data comparing the two CK isozymes, it is possible to make predictions regarding the operation of the PCr/CK circuit in the human brain. Most interestingly, the data suggest that the separate expression of BCK and uMtCK in astrocytes and many neurons would lead to a separate functionality in cells that have high BCK (and low uMtCK) and those with high uMtCK (and low BCK). It is important to first clarify the degree of coexpression of these two enzymes.
Whereas the expression of uMtCK in neurons was widespread, the neuronal expression of BCK was restricted to specific populations. Neurons that either did not express BCK at all, or expressed at very low levels, tended to be excitatory projection neurons. Most of the neurons of the cranial nerve nuclei were devoid of BCK reactivity, as were the pyramidal neurons in Ammon's horn of the hippocampus. Pyramidal neurons expressing SMI32 in the motor cortex had very low levels of BCK, similar to its reported distribution in the rat brain. 7 Those neurons that had higher BCK expression were, conversely, inhibitory neurons. Notable examples include Purkinje cells in the cerebellum, pallidal neurons, thalamic reticular neurons, and basket cells in the motor cortex. In fact, all these neurons are linked by their coexpression of the calcium-binding protein parvalbumin. The highest level of neuronal BCK expression was in medium spiny neurons of the striatum. Tachikawa et al 7 noted that there was no expression of BCK (or uMtCK) in oligodendrocytes. Indeed, in this study, BCK appeared to be expressed solely in astrocytes in white matter regions and did not colocalize with the oligodendrocyte marker Olig2.
Hence, it is better to consider the level of BCK expression in neurons to be on a continuous scale from no expression (excitatory projection neurons), low expression (Purkinje cells), moderate expression (pallidal neurons), to high expression (medium spiny neurons and basket cells). In contrast to this variable expression of BCK is the expression of uMtCK that is often assumed to be one part of a creatine circuit with BCK. 1 The expression levels of uMtCK were observed to be more consistent than BCK in neurons; however, marginally lower levels of uMtCK are visible in those cells that express high levels of BCK. Altogether, these results point to an opposing pattern of expression for the two CK isozymes.
The Differential Expression of the CK Isozymes is also Apparent in Muscle Tissue
The expression pattern of the CK isozymes is surprising because these enzymes are typically considered to be coexpressed as a requirement for the full use of the PCr/CK system in cells with high energy demands. Muscle fibers have high energy demands, although for different functions, and express high levels of CKs. 2 Skeletal muscle fibers express muscle-type CK (MCK) as the cytosolic CK isoform (comparable to BCK), and the sarcomeric mitochondrial CK (sMtCK) as the mitochondrial CK isoform (comparable to uMtCK). 1 These two enzymes are not expressed equally between different types of muscle fibers. In fact, like in the brain, MCK and sMtCK appear to have dissociated expression patterns: high sMtCK/low MCK in oxidative or slow-twitch muscle fibers, and low sMtCK/high MCK in glycolytic muscle fibers.12–14 As noted above, the expression patterns relate to the differential use of ATP synthesis pathways: sMtCK is higher in muscle fibers that mainly use oxidative phosphorylation to generate ATP (and also have higher numbers of mitochondria), 15 whereas MCK is higher in muscle fibers that mainly use glycolysis to generate ATP (and similarly have less mitochondria). 15
The correlation between MCK/sMtCK expression and energetic phenotype also translates, in general terms, to the brain; and helps explain the dissociated expression pattern of BCK and uMtCK. The oxidative/glycolytic division between slow- and fast-twitch muscle fibers is not so stark in brain cells. Oxidative phosphorylation is considered to account for upwards of 90% of ATP produced in the brain, 16 most of it used for maintaining ion fluxes across the plasma membrane.16,17 Like slow-twitch muscle fibers, neurons have high mitochondrial densities, 18 cristal packing density, 18 and typically show high cytochrome oxidase activity by histochemical staining. 19 The opposite tends to be true for astrocytes, which are more similar to fast-twitch muscle fibers.18,20 Depolarization of the astrocytic membrane by glucose deprivation cannot be reversed with the administration of pyruvate, whose energy is released by oxidative phosphorylation, but can be by the restoration of glucose.21–23 However, astrocytes are likely to make use of oxidative phosphorylation and glycolysis. Hertz et al 20 describe astrocytes having a ‘spatiotemporal dependence’ on glycolysis, meaning that astrocytes use glycolysis in specialized sites within the cell (where mitochondria are restricted) and for tasks where the high ATP synthesizing rate of glycolysis is advantageous over the slow but more sustained oxidative phosphorylation.
The Organization and Function of the PCr/CK System in the Human Brain
The oxidative-glycolytic division between neurons and astrocytes, and its similarity with the oxidative-glycolytic division in muscle fibers, indicates a different requirement for energy systems in either cell type. In the oxidative neuronal cells, uMtCK is widely and dominantly expressed. Coupling between this enzyme and adenine nucleotide translocator (ANT) in the inner mitochondrial membrane and porin in the outer mitochondrial membrane rapidly and specifically links ATP synthesized by oxidative phosphorylation with the PCr/CK system. 24 This improves the efficiency of oxidative phosphorylation by allowing the quick recycling of ADP, the principle driver of oxidative phosphorylation, back into the matrix. 24 With creatine, only micromolar concentrations of ADP are required to fully stimulate oxidative phosphorylation in permeabilized muscle cells, whereas without creatine, millimolar concentrations of ADP are required. 25 Indeed, in vitro studies on hippocampal neurons have shown that creatine can stimulate mitochondrial activity that promotes synaptogenesis. 26 Furthermore, the activity of uMtCK in the complex with ANT, porin, and cyclophilin D in the matrix reduces the formation of the apoptosis-inducing mitochondrial permeability transition. 24
Conversely, in the glycolytic astrocytic cells, BCK is expressed without uMtCK, which was also noted in the rat brain. 7 Functionally, the cytosolic localization of BCK means that there is no specific compartmentation of a CK enzyme to mitochondria, although BCK in the proximity of mitochondria can still integrate ATP produced by oxidative phosphorylation into the PCr/CK circuit once it has diffused out. Theoretical analysis of a system with a cytosolic CK expressed homogeneously within a volume and at near equilibrium points to the ability of such a system to support the buffering of ATP temporally and spatially, the latter via facilitated diffusion. 3 In several 31P nuclear magnetic imaging (NMR) magnetization transfer studies, the forward and reverse rate constants for CK in the brain are statistically indistinguishable, suggesting that they are near equilibrium.27,28 Thus, alone, BCK is likely to be mediating the buffering of ATP in the cytosol as it is consumed by energy-intensive processes, and speeding the flux of high-energy phosphoryls around the cell. Hertz et al 20 suggest that the latter function is important in delivering energy to regions of astrocytes, there called peripheral astrocytic processes, where the available volume is too small to accommodate mitochondria.
The association between the cytosolic CK and glycolytic-type muscle fibers also suggests that BCK in astrocytes may support energetic features akin to that seen in like-muscle fibers: a form of ‘metabolic capacitance’ whereby CK activity smoothes large spikes in energy consumption as discussed by Sweeney. 29 Possible sources of such ‘spiking’-type energy demands in astrocytes is still ambiguous, although the close link between astrocytes and glutamate signaling between neurons is one possible source. 20 Indeed, ion fluxes, which in astrocytes result from stimulation by glutamate or the clearance of extracellular potassium, 20 are thought to be linked to energy derived from glycolysis. 16
In some neurons, BCK and uMtCK are coexpressed. In our study, these tend to be interneurons that express parvalbumin, but notably also include striatal medium spiny neurons. The combination of uMtCK-enhancing oxidative metabolism and the spatiotemporal support of cytosolic energy consumption by BCK are the key features provided to such neurons. A further synergistic feature is the establishment of the ‘phosphocreatine shuttle’ that allows the directed transfer of energy, carried on PCr, between sites that the CK isozymes become targeted to. 1 One is uMtCK in the mitochondrion that links the PCr/CK system to ATP produced by oxidative phosphorylation. The other, less well-defined aspect, is the functional (or sometimes physical) association of the cytosolic CK isozyme with ATP-producing and -consuming sites in the cytosol. This has been well noted in muscle fibers where MCK is known to associate with, and therefore target PCr to, the Na+ /K+ ATPase, glycolytic complexes, the sarco/endoplasmic reticulum calcium ATPase, and the myofibrillar ATPase. 1 In the brain, this has been less well studied, but several papers have described the association between BCK and the neuron-specific potassium cotransporter KCC2.30,31 This transporter is important for the formation and maintenance of dendritic spines. 32 The BCK-KCC2 interaction is important in inhibitory neurons, notably where BCK is expressed, because its loss can depolarize the reversal potential mediating inhibitory signaling, viz, the chloride conductance. 31 These compartmentalized CK isozymes would clearly not operate near equilibrium despite the results of the 31P-NMR magnetization transfer studies (see above).27,28 However, as Wallimann 33 discusses in detail, it would appear that such compartmentalized CK isozymes may be ‘invisible’ to the current 31P-NMR magnetization transfer study protocols.
The specific expression of BCK in inhibitory interneurons may also relate to the energetic properties of these neurons derived from their signaling behaviors. Excitatory neurons, which tend to have very low or undetectable levels of BCK expression, overall have consistent but low firing rates, which in part relates to the high energetic cost of excitatory signaling.17,34 These energetic properties appear more amenable to a system of energy delivery that favors consistency and longevity—that is, afforded by mitochondrial oxidative phosphorylation and is akin to sMtCK-enriched oxidative slow-twitch muscle fibers.14,34 In contrast, inhibitory neurons often show very high but very short firing rates that may require a system that can deliver large amounts of energy in a short period of time—that is, afforded by glycolysis in the short term—and also probably oxidative phosphorylation in the long term, which is akin to MCK-enriched fast-twitch glycolytic muscle fibers.14,34
The strong expression of BCK in medium spiny neurons is enigmatic. These neurons have low firing rates (1 Hz), 34 but show high oxidative capacity by cytochrome oxidative histochemistry 35 despite being inhibitory. However, they are not interneurons, being one of the few inhibitory projection neurons in the brain. They also show a selective sensitivity to neurotoxins that target oxidative energy metabolism, such as 3-nitropropionic acid and malonate. 36 These toxins are often used to produce striatal lesions that mimic some of the symptoms and changes in HD. 36 Medium spiny neurons receive large numbers of excitatory inputs from the cortex and thalamus, which impinge on a dense network of spiny dendrites. This would create a major energetic burden on the neurons17,37 and is thought to be the reason why the striatum shows such high energy requirements but low firing rates. 34 Moreover, it is especially notable that toxicity associated with overexcitation of the striatum is a key feature of HD and that the effects of the energy-impairing neurotoxins can be attenuated with N-methyl-D-aspartate receptor antagonists. 36 Several lines of evidence give an indication of the importance of glycolysis in medium spiny neurons. During neuronal stimulation in the striatum, Lowry and Fillenz 38 showed a period where there is an increase in glucose consumption without an increase in oxygen consumption. During HD, an increased ratio of the cerebral metabolic rate of oxygen consumption to glucose consumption is an indication of an impairment in glycolysis. 39 Furthermore, the selective inhibition of glycolysis with iodoacetate increases the susceptibility of medium spiny neurons to excitotoxic cell death in the R6/2 transgenic mouse model of HD. 40 All of these properties seen in medium spiny neurons point to a neuron type that is energetically unique in the brain: a strong dependence on oxidative metabolism and glycolysis to cope with the high energy demands as a result of its connectivity. Separately, BCK and uMtCK expressed in these neurons can support oxidative and glycolytic metabolism of glucose as discussed. They probably also synergistically support energy metabolism in the medium spiny neurons by the formation of the phosphocreatine shuttle, directing the flux of energy toward high energy-consuming sites, possibly including KCC2.30,31 It is further notable that during HD, there is a loss of BCK in the striatum 6 that further reinforces the energy-related deficits seen in this disease.
Summary and Conclusion
This study has characterized the distribution of the two key CK isozymes expressed in the brain. Their distribution was found to be contrasting and sometimes mutually exclusive. By considering the energetic properties of the classes of cells that express each combination of CK isozymes, and relating them to the functions that this implies, it becomes clear that the brain expresses different combinations of these enzymes, matching the energetic requirements of each cell type. This study hypothesizes that uMtCK supports oxidative energy metabolism that is widely used by neurons that require consistent and reliable energy supplies. In contrast, it hypothesizes that BCK supports glycolytic processes in astrocytes and some neurons that show uniquely high and fluctuating energy requirements. These hypotheses parallel the well-studied distributions and functions of these enzymes in muscle fibers.1,14 All aspects of the PCr/CK system are probably used to different degrees in the brain: from spatiotemporal buffering in astrocytes3,14,20 to directed shuttling of high energy phosphoryls by the phosphocreatine circuit model in neurons.1,14
Further complexity is expected with a more in-depth study of this and other energetic systems in the brain. ‘Microcompartmen-tation,’ the term for the specific compartmentation of select enzymes within cells, is now considered to be a key mechanism by which cells control metabolic processes in a dense and complex three-dimensional environment. 14 Like the ‘tip of the iceberg’ metaphor, this study of the PCr/CK system clarifies some key organization features of creatine usage in the brain, but indicates much more still unseen, ready to be revealed.
DISCLOSURE/CONFLICT OF INTEREST
The authors declare no conflict of interest.