
Abstract
Intracellular pH (pHi) in the vascular wall modulates agonist-induced vasocontractile and vasorelaxant responses in mesenteric arteries, whereas effects on myogenic tone have been unsettled. We studied the role of Na+,HCO3− cotransporter NBCn1 in mouse isolated middle cerebral arteries and the influence of pHi disturbances on myogenic tone. Na+,HCO3− cotransport was abolished in arteries from NBCn1 knockout mice and steady-state pHi ∼0.3 units reduced compared with wild-type mice. Myogenic tone development was low under control conditions but increased on treatment with the NO-synthase inhibitor N-nitro-
INTRODUCTION
Cellular acid extrusion in mesenteric arteries is mediated by the Na+,HCO3− cotransporter NBCn1 (slc4a7)1, 2 and the Na+/H+ exchanger NHE1 (slc9a1).3, 4 While NBCn1 is active in the near-physiologic intracellular pH (pHi) range and important for regulation of resting steady-state pHi, NHE1 is active primarily at low pHi values.2, 4 During mesenteric artery constriction, NBCn1 is activated in the vascular smooth muscle cells (VSMCs) through a Ca2+-mediated, calcineurin-dependent mechanism to extrude the excess acid load associated with VSMC contraction. 5 If the net acid extrusion mechanisms are compromised, intracellular acidification ensues and the vasomotor function of the mesenteric arteries is substantially affected; acute intracellular acidification of VSMCs causes vasoconstriction, 6 whereas sustained intracellular acidification of VSMCs inhibits rho-kinase-dependent Ca2+ sensitivity.2, 4 In addition, acidification of endothelial cells in mesenteric arteries inhibits NO-synthase activity and NO-mediated vasorelaxation,2, 4 whereas endothelial cell alkalinization attenuates gap-junction-dependent intercellular coupling and endothelium-dependent hyperpolarizations. 7 These findings have established a fundamental role for pHi in modulating the vasomotor function of mesenteric arteries. In contrast, remarkably little is known about the regulation of pHi and its functional implications in other vascular beds, including cerebral arteries where NBCn1 is known to be expressed,1, 8 but its functional role for pHi regulation and vasomotor function requires further investigation.
Although arteries from different vascular beds share many functional characteristics, prominent differences have also been described concerning, e.g., responses to arachidonic acid metabolites 9 and changes in transmural pressure, 10 expression levels for adrenergic and muscarinic receptors, 11 and mechanisms of agonist-induced endothelium-dependent relaxations. 10 These functional variations likely reflect in part the diverse roles of different arterial beds; mesenteric arteries, for instance, are strongly influenced by transmitter release from perivascular sympathetic nerves and have a major role for the control of peripheral vascular resistance and blood pressure. 12 In contrast, cerebral artery tone is thought to be modulated primarily by local metabolic, paracrine, and mechanical factors, and the major role of the cerebral vasculature is to adjust cerebral blood flow to the local metabolic demand in spite of changes in hemodynamic conditions. The myogenic response has a critical role in the autoregulation of cerebral blood flow and in control of capillary pressure. 13 Even though the myogenic response to an increased transmural pressure was already described in 1902 by Bayliss, 14 many elements of its mechanistic background remain controversial. Nonetheless, it is well-accepted that NO-mediated and rho-kinase-dependent signaling are involved in the regulation of myogenic tone of cerebral arteries.15, 16 On the basis of the influence of pHi on NO-mediated and rho-kinase-dependent signaling in mesenteric arteries,2, 4, 17 we hypothesized that pHi disturbances interfere with myogenic tone development in cerebral arteries.
In the present study, we show that NBCn1 is responsible for the Na+,HCO3− cotransport in VSMCs of middle cerebral arteries and has a major role for the regulation of pHi. Disruption of NBCn1 expression interferes with myogenic tone development in the middle cerebral arteries and, thus, likely impedes the capacity for adjusting vascular resistance and cerebral blood flow in response to changes in cerebral perfusion pressure.
MATERIALS AND METHODS
NBCn1 knockout mice were generated on the 129/SVJ genetic background as previously described, 2 and were backcrossed for at least 10 generations into C57BL/6 J mice. Middle cerebral arteries were isolated from 7 to 12 weeks old, male, NBCn1 knockout and wild-type mice, mounted in pressure myographs (110P or 120CP; DMT, Aarhus, Denmark) for isobaric investigations, and studied on the stage of an Olympus CK40 inverted microscope equipped with an Olympus NDA × 10 objective (Ballerup, Denmark; numerical aperture 0.25). The physiologic saline solution used for these studies contained 114 mM NaCl, 10 mM HEPES, 25 mM NaHCO3, 1.20 mM MgSO4, 4.70 mM KCl, 5.50 mM glucose, 26 μM EDTA, 1.18 mM KH2PO4, and 1.60 mM CaCl2. The physiologic saline solution was heated to 37 °C, continuously aerated with a gas mixture of 5% CO2 balance air, and adjusted to pH 7.40. Images of the pressurized arteries were recorded using a USB CCD Color Camera (DMK 41AU02, The Imaging Source, Bremen, Germany) and the outer diameter of the arteries was traced in real time using DMTVAS 6.2 software (IonOptix LLC, Milton, MA, USA). The level of myogenic tone was determined from the diameter response to stepwise increases in transmural pressure from 20 to 100 mm Hg. Responses to serotonin, the thromboxane analog U46619, or depolarization with 80 mM extracellular K+ were determined at a transmural pressure of 80 mm Hg. The level of tone under any given condition was calculated as: Tone=(Dmax−D)/Dmax; where D is the diameter under the given experimental conditions and Dmax is the diameter at the same transmural pressure but in the combined presence of 10 μM papaverine (phosphodiesterase inhibitor) and 10 μM Y-27632 (rho-kinase inhibitor) or under Ca2+-free conditions in the presence of 5 mM EGTA. All animal procedures were conducted according to the Danish Animal Welfare Legislation and approved by the Danish Animal Care and Use Committee.
Intracellular pH Measurements
Fluorescence microscopy of 2′,7′-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein (BCECF)-loaded middle cerebral arteries was performed to measure pHi of VSMCs. Arteries were mounted in a pressure myograph (DMT), incubated with 5 μM BCECF-AM in 0.02% dimethyl sulfoxide for 20 minutes at 37 °C, and investigated on the stage of a Leica DM IRB inverted microscope (Ballerup, Denmark) connected to a Photon Technology International Deltascan system (Birmingham, NJ, USA). A Leica HCX APO × 20 objective (Ballerup, Denmark; numerical aperture 0.5) was employed. Arteries were alternately excited at 440 and 495 nm, while the emission light was collected at 530 nm. Background fluorescence was measured before the loading procedure and subtracted from the measured emissions before calculation of fluorescence ratios. To minimize light-induced damage to the tissue, exposure to excitation light was reduced by obstructing the light path during experimental procedures that did not require continuous measurements. Calibration of BCECF fluorescence ratios to pH units was performed using the high-K+ nigericin method, as previously described. 18 The high-K+ nigericin method has previously been shown to be in good agreement with the null-point technique in isolated arteries. 5 The buffering capacity was estimated from the pHi change observed on washout of NH4Cl as described earlier. 1 No significant difference in the total buffering capacity of VSMCs was seen between arteries from wild-type and NBCn1 knockout mice (P=0.25). Total VSMC buffering capacity (βtot) in the pHi range 6.7 to 7.6 was pHi dependent and approximately described by the one-phase exponential decay function βtot=1.35·108e−2.07·pH. These values are very similar to previous reports from mouse arteries.1, 2, 3, 4 On the basis of the similar buffering capacity of VSMCs in arteries from NBCn1 knockout and wild-type mice, pHi recovery rates were measured at equivalent pHi values and compared directly.
Vasomotion
To analyze the oscillatory component of the tone development, the diameter measurements were transferred to LabChart 7 Pro (AD Instruments, Dunedin, New Zealand). Cyclic height and rate were analyzed at a transmural pressure of 80 mm Hg, using a minimum peak height of 2 μm and a minimum frequency of 2 per minute during the oscillatory phase with greatest amplitude.
Intracellular [Ca2+] measurements
Measurements of [Ca2+]-dependent fluorescence were performed on the same setup described above for pHi measurements, but arteries were loaded with 6 μM Fura2-AM in a loading buffer containing dimethyl sulfoxide (final concentration 0.01%), pluronic F127, and Cremophor EL at 37 °C two times for 30 minutes. The Fura2 fluorescence ratio (F340/F380) was taken as a measure of intracellular [Ca2+]. Further experimental details have been described before. 1
Membrane Potential Measurements
Arteries were mounted in a pressure myograph (DMT) as described above. Membrane potential (Vm) measurements were performed using aluminium silicate microelectrodes (WPI, Hitchin, UK) with a resistance of 40 to 120 MΩ when backfilled with 3 mol/L KCl, recorded with an Intra-767 amplifier (WPI), visualized on an oscilloscope (Gould-Nicolet Technologies, Loughton, UK) and continuously stored with a PowerLab system (ADInstruments). To measure membrane potentials from VSMCs, the electrode was advanced into the vessel wall from the adventitial side. Electrode entry into cells resulted in an abrupt drop in voltage followed by a sharp return to baseline on retraction.
Quantitative Reverse-Transcriptase PCR
The level of NBCn1 mRNA in isolated middle cerebral arteries from wild-type and NBCn1 knockout mice was investigated using two-step TaqMan quantitative reverse-transcriptase PCR as previously described. 1 Glyceraldehyde-3-phosphate dehydrogenase and the transferrin receptor were used as housekeeping genes to correct for variations in RNA isolation efficiency. The primer and probe sequences were: NBCn1, forward 5′-GCA AGA AAC ATT CTG ACC CTC A-3′, reverse 5′-CTC CAC TTC CGT TAC CTT TCA T-3′, probe 5′-TCC TGG AAA CTT GGA CAA TAG TAA AAG TGG TG-3′; glyceraldehyde-3-phosphate dehydrogenase, forward 5′-CAC GGC AAA TTC AAC GGC ACA G-3′, reverse 5′-AGA CTC CAC GAC ATA CTC AGC ACC-3′, probe 5′-AGC TTG TCA TCA ACG GGA AGC CCA TCA CGA-3′; transferrin receptor, forward 5′-TTT GGG CAC TAG ATT GGA TAC CT-3′, reverse 5′-GGT TCA ATT CAA CGT CAT GGG TA-3′, probe 5′-CAG CGG AAG TGG CTG GTC AGC TCA TTA TTA AA-3′.
Statistics
All values are given as mean±s.e.m. Measurements of single variables were compared between arteries from NBCn1 knockout and wild-type mice by unpaired, two-tailed Student's t-tests. When the effect of two variables on the measured variable was analyzed, two-way analysis of variance was performed followed by Bonferroni post tests. The effect of N-nitro-
RESULTS
The NBCn1 knockout mice employed in the current study were generated based on a functional genomics approach using a gene trap vector incorporated into the GC-rich region upstream of exon 1. 2 This approach completely eliminated NBCn1 mRNA expression in the middle cerebral arteries; the relative expression of NBCn1 was 0.004±0.0004 in arteries isolated from NBCn1 knockout mice (n=6) compared with 1.00±0.12 in arteries from wild-type mice (n=6; P<0.001; unpaired, two-tailed Student's t-test). This finding is consistent with previous findings from mesenteric arteries. 2
Intracellular pH Regulation
Regulation of pHi in VSMCs is largely Na+ dependent and is mediated by Na+/H+ exchange and Na+,HCO3− cotransport. 19 To determine the contribution of NBCn1 to Na+,HCO3− cotransport in the middle cerebral arteries, we measured the rate of amiloride-insensitive, Na+-dependent pHi recovery from intracellular acidification (Figures 1A and 1B). An original trace is shown in Supplementary Figure 1. Although Na+,HCO3− cotransport was prominent in arteries from wild-type mice, it was essentially abolished in arteries from NBCn1 knockout mice (Figures 1A and 1B). Further supporting a major role for NBCn1-mediated Na+,HCO3− cotransport in VSMCs of middle cerebral arteries, we found that the resting steady-state pHi of VSMCs was ∼0.3 units lower in arteries from NBCn1 knockout than wild-type mice (Figure 1C).
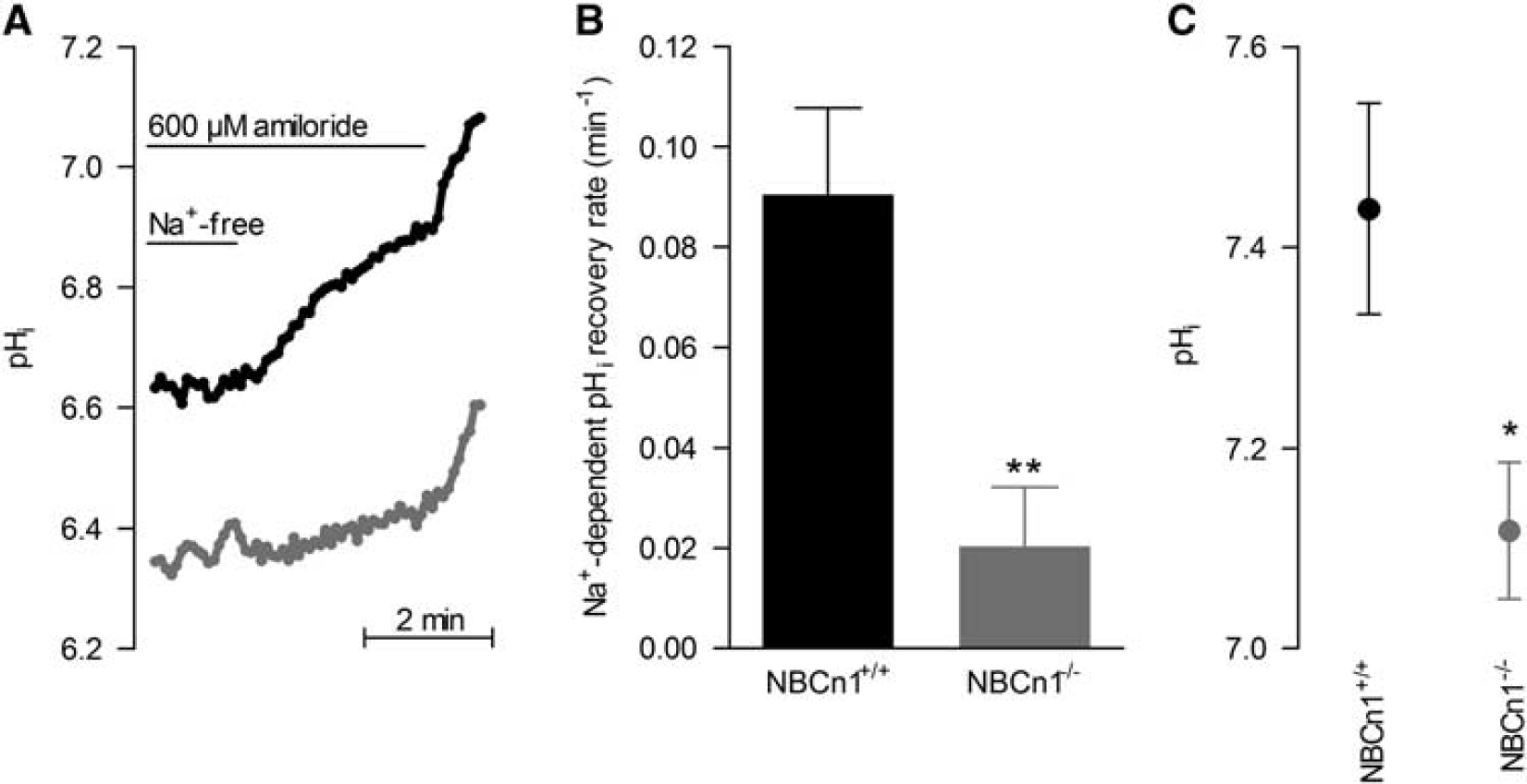
Regulation of intracellular pH (pHi) is impeded in vascular smooth muscle cells (VSMCs) of middle cerebral arteries from NBCn1 knockout mice. (
Myogenic Tone
We have previously shown that even moderate changes in pHi (0.1–0.2 units magnitude) can drastically alter mesenteric artery responses to agonist stimulation. 2 Here we investigated whether the lack of NBCn1 also affected the vasomotor function of middle cerebral arteries. We investigated the level of myogenic tone during stepwise changes in transmural pressure from 20 to 100 mm Hg. The level of myogenic tone was calculated by comparing the diameter under the given condition to the fully relaxed arterial diameter obtained in the presence of 10 μM papaverine and 10 μM Y-27632. In the presence of Y-27632 and papaverine, no difference in diameter was observed between middle cerebral arteries from NBCn1 knockout and wild-type mice (Figure 2), and each square-step increase in transmural pressure elicited a square-step increase in vessel diameter with no evidence of a vasocontractile response to the increased wall stress (Figures 3A and 3B).
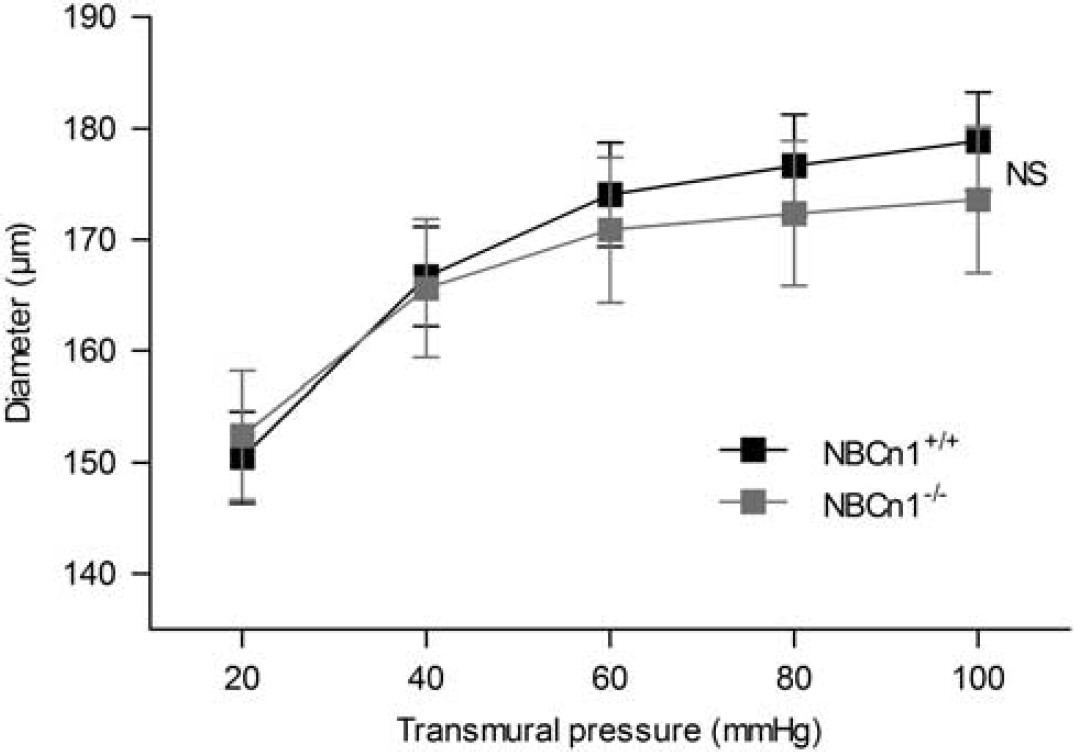
Average relaxed outer diameters of middle cerebral arteries from NBCn1 knockout and wild-type mice (n=8–10). Arteries were investigated in the presence of 10 μM Y-27632 and 10 μM papaverine. Under these conditions, arterial diameters increased when the transmural pressure was raised and no sign of active vasoconstriction was observed. The comparison was performed by repeated measures two-way analysis of variance. NS, not significantly different.
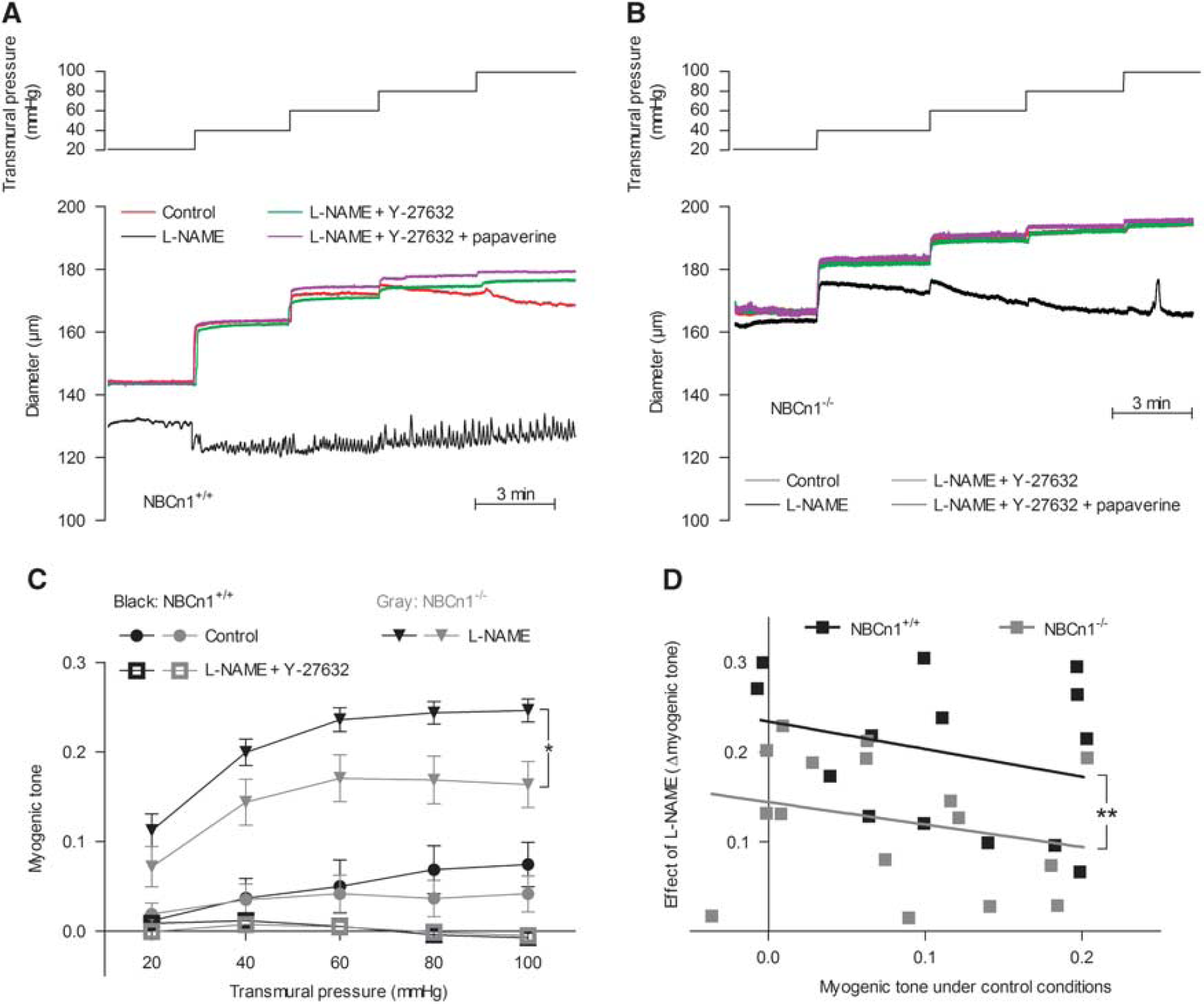
Myogenic tone development in middle cerebral arteries from NBCn1 knockout mice is reduced in the presence of N-nitro-
Under control conditions, surprisingly, low myogenic tone was observed in middle cerebral arteries from both wild-type and NBCn1 knockout mice (Figures 3A–C). This appeared to be consequent to a high basal NO production, as incubation with the NO-synthase inhibitor L-NAME greatly augmented the level of myogenic tone (Figures 3A–C). The response to L-NAME was significantly blunted in arteries from NBCn1 knockout mice compared with arteries from wild-type mice even when the initial level of myogenic tone was taken into consideration (Figure 3D). This result is consistent with our previous finding from mesenteric arteries that NO-synthase activity and NO-mediated vasorelaxation are reduced at low pHi.2, 4
In the presence of L-NAME, the level of myogenic tone was significantly lower in middle cerebral arteries from NBCn1 knockout compared with wild-type mice (Figures 3A–C). To investigate the mechanistic background for this apparent inhibition of contractile function, we investigated the level of intracellular [Ca2+] in the VSMCs. In the presence of 100 μM L-NAME, no difference in the Fura2 fluorescence ratio was seen between VSMCs in middle cerebral arteries from wild-type and NBCn1 knockout mice over the range of transmural pressures investigated (20–100 mm Hg; Figure 4A), suggesting an effect of NBCn1 knockout and consequent low pHi on VSMC Ca2+ sensitivity. We have previously shown that the rho-kinase has a moderate pH sensitivity in vitro and that rho-kinase-dependent signaling is inhibited in mesenteric arteries from NBCn1 knockout mice. 2 On this background, we investigated the effect of the rho-kinase inhibitor Y-27632 (10 μM) on the level of myogenic tone and VSMC [Ca2+] in middle cerebral arteries treated with L-NAME. In the presence of Y-27632, myogenic tone development was completely abolished in arteries from both wild-type and NBCn1 knockout mice (Figures 3A–C). However, intracellular [Ca2+] in the VSMCs still increased when the transmural pressure was raised in the presence of Y-27632 and no difference in the Fura2 fluorescence ratio was observed between arteries from wild-type and NBCn1 knockout mice (Figure 4B). These findings demonstrate that myogenic tone in mouse middle cerebral arteries is absolutely dependent on rho-kinase signaling and are consistent with NBCn1 knockout causing a pHi-mediated inhibition of rho-kinase signaling, ultimately reducing VSMC Ca2+ sensitivity and myogenic responsiveness.
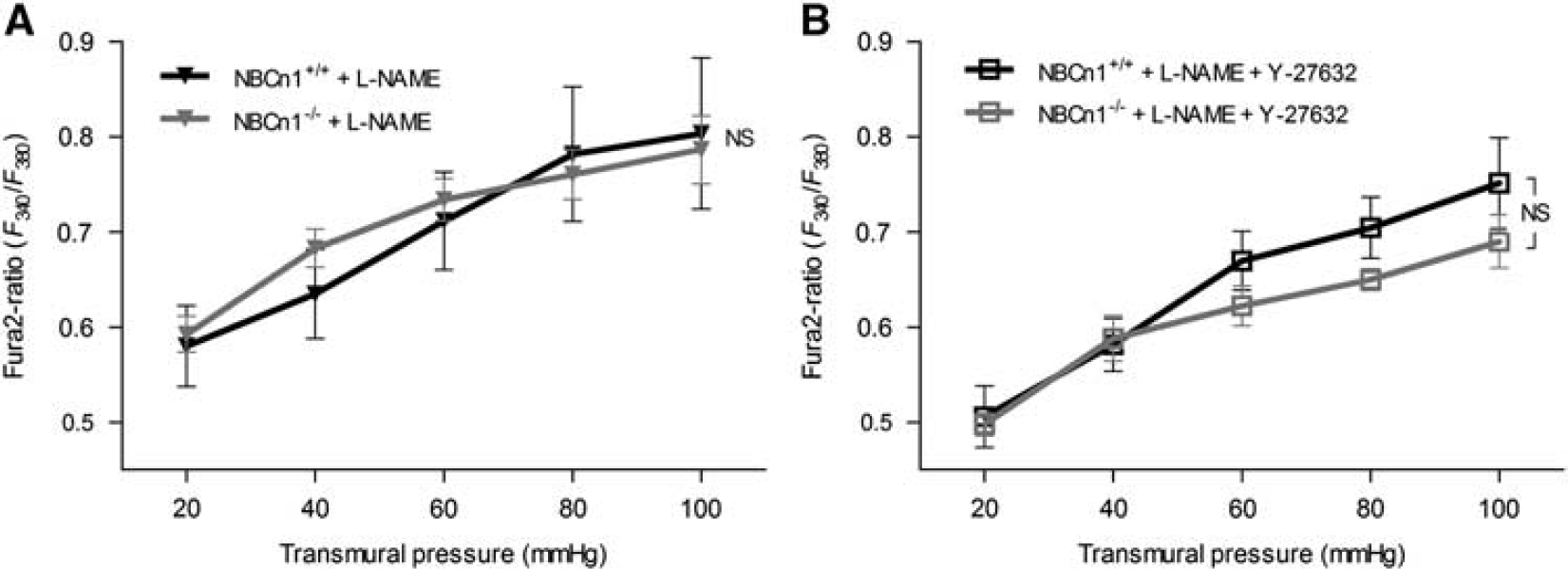
Intracellular Ca2+ responses to changes in transmural pressure are unaffected by knockout of NBCn1. (
Under control conditions, the outer diameter of the middle cerebral arteries was typically steady with no obvious oscillatory behavior (Figures 3A and 3B). On application of L-NAME, 60±13% of arteries from wild-type mice (n=15) and 65±12% of arteries from NBCn1 knockout mice (n=17) developed intermittent vasomotion characterized by rhythmic oscillations in the vessel diameter (Figures 3A and 5). The ability of L-NAME to induce vasomotion in the cerebral vasculature is consistent with previous reports.20, 21 The frequency of the L-NAME-induced vasomotion was very similar between arteries from NBCn1 knockout and wild-type mice (Figures 5A and 5C), whereas the amplitude of the oscillations was reduced to around half in arteries from NBCn1 knockout compared with wild-type mice (Figures 5A and 5B). The magnitude of the diameter oscillations was fairly small compared with the overall level of tone and, consequently, no clear oscillatory pattern (distinct from background noise) could be detected in the Fura2-based intracellular [Ca2+] measurements.
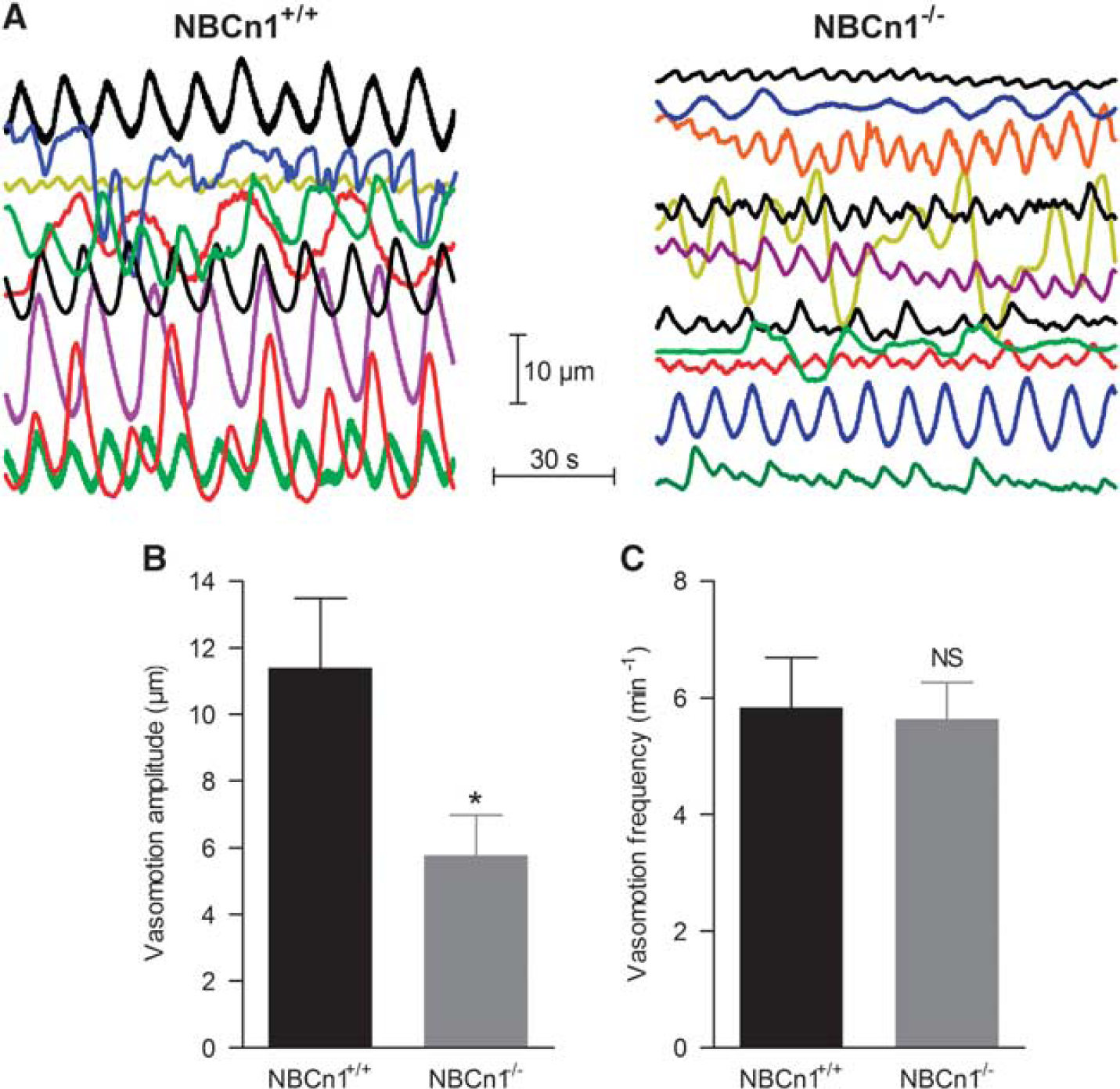
The amplitude of the vasomotion is reduced in middle cerebral arteries from NBCn1 knockout compared with wild-type mice. (
Previous studies by other groups have demonstrated that pHi can modulate ion channel function. 22 Hence, pHi could be expected to modulate VSMC membrane potential and an effect on membrane potential could be predicted to contribute to the reduced myogenic tone observed in middle cerebral arteries from NBCn1 knockout mice in the presence of L-NAME. We found, however, no difference in the resting VSMC membrane potential between arteries from NBCn1 knockout and wild-type mice at a transmural pressure of 80 mm Hg in the presence of 100 μM L-NAME (Figure 6A). Consistent with previous reports, inhibition of NO production with L-NAME depolarized the VSMCs, 23 but although there was a tendency towards a smaller L-NAME-induced depolarization of VSMCs in arteries from NBCn1 knockout mice, this did not reach statistical significance (Figure 6B). Increasing the transmural pressure from 20 to 80 mm Hg in the presence of L-NAME depolarized the VSMCs to a similar extent in middle cerebral arteries from NBCn1 knockout and wild-type mice (Figure 6B). These findings suggest that membrane potential control in VSMCs of arteries from NBCn1 knockout mice is not markedly affected and are consistent with the unaltered levels of intracellular [Ca2+] described above (Figure 4). Taken together, the membrane potential recordings further reinforce the conclusion that sustained low pHi interferes with the contractile function of middle cerebral arteries primarily through inhibition of VSMC Ca2+ sensitivity.
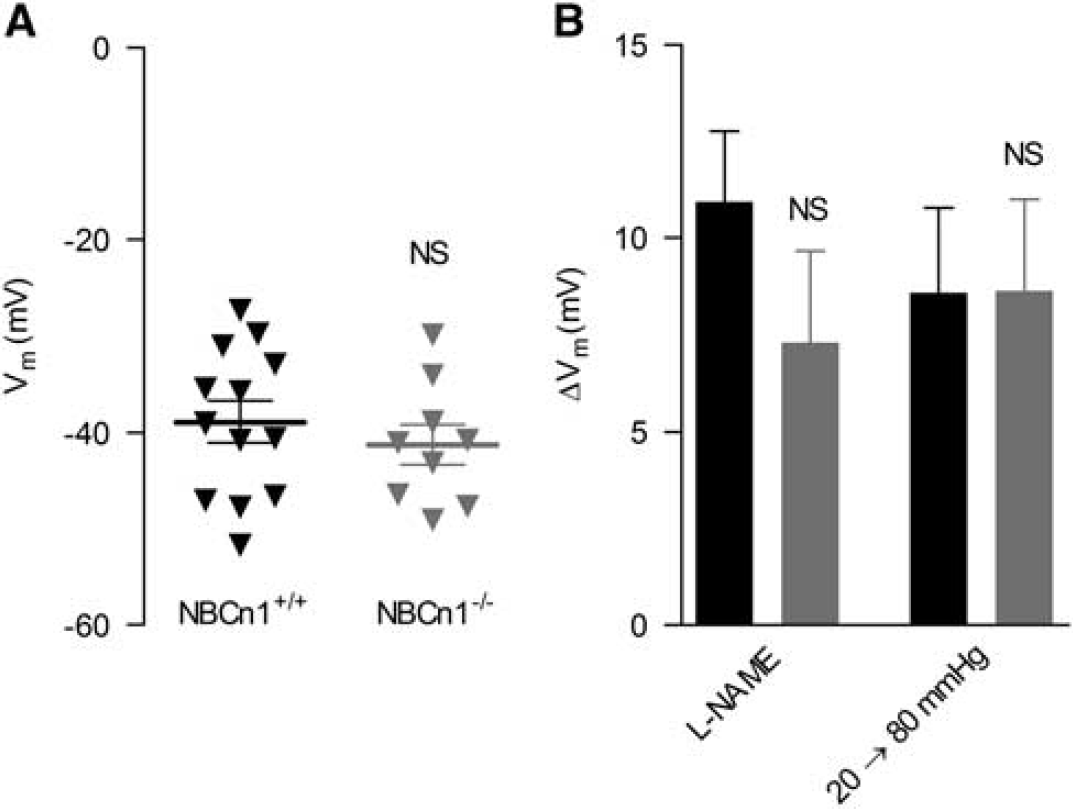
Membrane potential control is unaffected in vascular smooth muscle cells (VSMCs) of middle cerebral arteries from NBCn1 knockout mice. (
Responses to Depolarization and Agonist Stimulation
To further investigate the vasomotor differences between middle cerebral arteries from NBCn1 knockout and wild-type mice, we determined the responses to depolarization, application of serotonin, or stimulation with the thromboxane analog U46619. At a transmural pressure of 80 mm Hg, we found that middle cerebral arteries from wild-type and NBCn1 knockout mice contracted to a similar extent when depolarized by 80 mM extracellular K+ (Figure 7A). In the presence of L-NAME, as described above, basal myogenic tone was significantly reduced in arteries from NBCn1 knockout mice compared with arteries from wild-type mice (Figure 7A). However, depolarization caused by increasing the extracellular [K+] to 80 mM induced strong contractions in both groups of arteries also in the presence of L-NAME (Figure 7A).
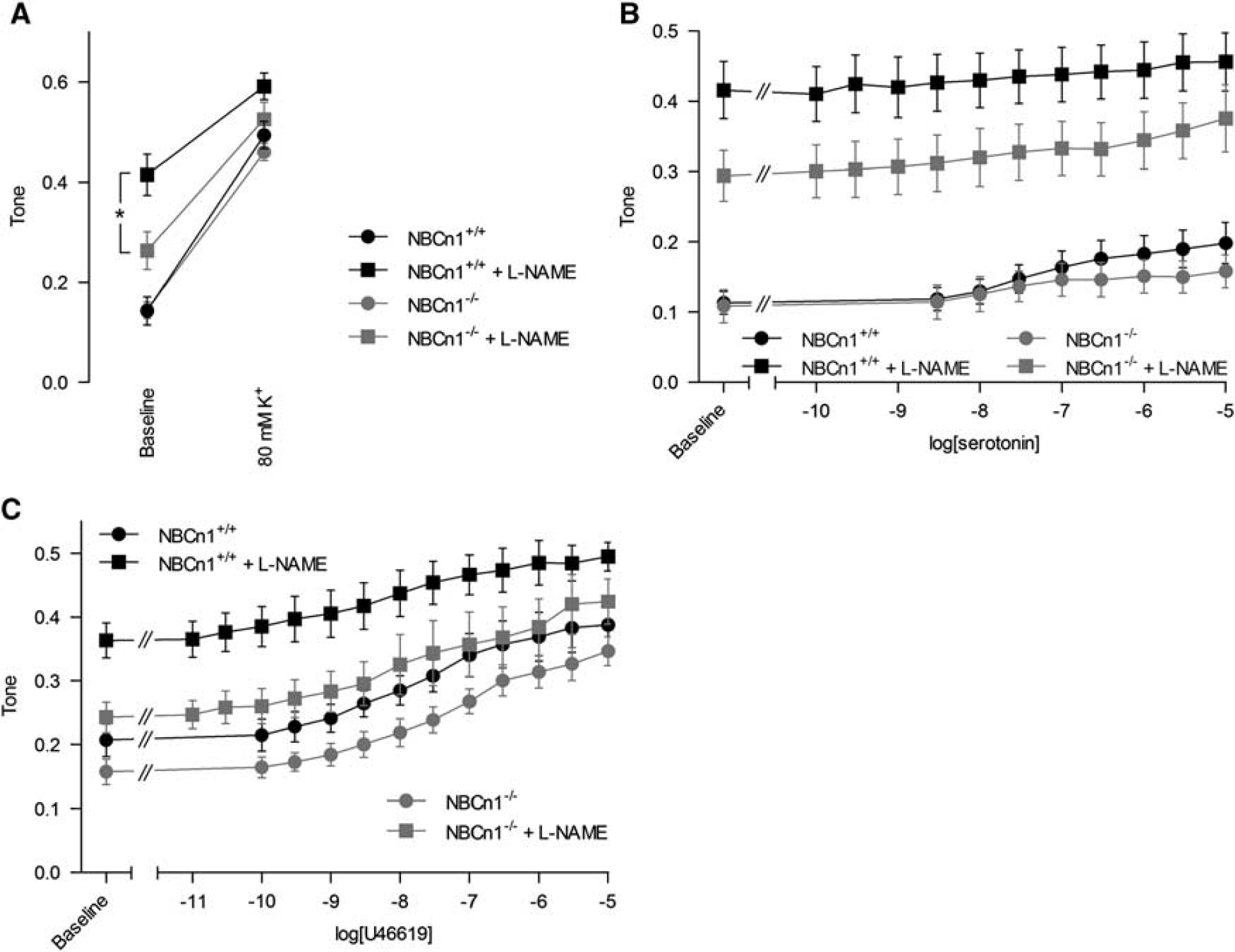
Contractions of middle cerebral arteries from NBCn1 knockout and wild-type mice to agonist stimulation and depolarization are similar under control conditions. In the presence of N-nitro-
Serotonin and U46619 both produced concentration-dependent contractions of the mouse middle cerebral arteries (Figures 7B and 7C). The response to serotonin was, however, relatively small (Figure 7B). In both cases, application of 100 μM L-NAME increased the level of myogenic tone (Figures 7B and 7C1). Because of the difference in baseline myogenic tone between arteries from NBCn1 knockout and wild-type mice in the presence of L-NAME, comparison of the agonist-induced contractile responses was complicated. Overall, these experiments suggest that the sustained intracellular acidification observed in middle cerebral arteries from NBCn1 knockout mice most prominently affects the development of myogenic tone. These findings are consistent with the previously reported pronounced importance of rho-kinase signaling for myogenic tone development. 24
DISCUSSION
We show here that NBCn1 mediates the transplasmalemmal Na+,HCO3− cotransport, which is crucial for pHi control in mouse middle cerebral arteries at steady state and during intracellular acidification. Disruption of NBCn1 expression in the middle cerebral arteries results in intracellular acidification, inhibits NO-mediated and rho-kinase-dependent signaling, and interferes with myogenic vascular reactivity. These findings expand on previous results from mouse mesenteric arteries, demonstrating that a normal pHi in the vascular wall is important for vasomotor responses to vasocontractile (norepinephrine) as well as vasorelaxant (acetylcholine) agonists.2, 4 The current study implies that NBCn1—in addition to being essential for regulation of blood pressure in mice 2 and potentially humans 25 —is important for the regulation of cerebral artery tone in response to changes in transmural pressure and hence is required for autoregulation of cerebral blood flow.
As knockout of NBCn1 inhibits both vasodilatory and vasocontractile signaling cascades, the net result is rather small under control conditions. It should be noted, however, that the lack of normal NO-mediated signaling and reduced rho-kinase-dependent Ca2+ sensitivity will impede the ability of the arteries to respond adequately to relevant external stimuli. Consequently, a normal pHi level is likely to be important for regulating cerebral blood flow in response to changes in hemodynamic conditions.
The development of myogenic tone in middle cerebral arteries from NBCn1 knockout mice takes place over the same transmural pressure range as in arteries from wild-type mice. In congruence, the depolarization of VSMCs caused by an increase in transmural pressure from 20 to 80 mm Hg was similar in arteries from NBCn1 knockout and wild-type mice, and no difference in the levels of intracellular [Ca2+] were observed. These findings imply that the sensing mechanism for transmural pressure (or wall stress) was not affected by the knockout of NBCn1. Instead, the contractile response to raised intracellular [Ca2+] evoked by increased transmural pressure was reduced because of a lower VSMC Ca2+ sensitivity.
Although multiple ion channels expressed in the vascular wall have been found to be activated (e.g., TRPV3) 26 or inhibited (e.g., large conductance Ca2+-activated K+ channels) 22 by low pHi, the VSMC membrane potential was unaffected by a sustained decrease in pHi of ∼0.3 units. In addition, although both Ca2+ transport across the plasma membrane and release from intracellular stores have been reported to be affected by changes in pHi,27, 28 our current and previous findings strongly suggest that overall Ca2+ handling in VSMCs is essentially unaffected by sustained intracellular acidification.1, 2, 4 This is in contrast to acute changes in pHi, which have consistently been shown to cause transient changes in the intracellular [Ca2+] levels, 19 most likely because of combined effects on Ca2+ transport and competition between H+ and Ca2+ for buffer binding.29, 30
Ca2+ sensitization in VSMCs can be mediated by both rho-kinase- and protein kinase C (PKC)-dependent signaling pathways. However, in the mouse middle cerebral arteries, we found that myogenic tone development was completely abolished by rho-kinase inhibition with Y-27632. This is consistent with previous studies on rat posterior cerebral arteries showing a major effect of rho-kinase inhibition15, 16, 31 and a smaller contribution from PKC signaling. 31 Y-27632 has a good selectivity for the rho-kinase over the traditional PKC isoforms. Only PKC-δ is significantly inhibited in the applied concentration range 32 and this Ca2+-independent PKC isoform is unlikely to have a major role in cerebral arteries where PKC activation has been shown to be Ca2+ dependent 33 and PKC-α has been identified as the most important PKC isoform. 34 Nevertheless, it should be noted that PKC-δ has been suggested to contribute to trafficking of TRPM4 to the plasma membrane of VSMCs. 35 As we see no effect of NBCn1 knockout on the VSMC membrane potential or the level of intracellular [Ca2+], it is however unlikely that TRPM4 and PKC-δ have a major role for the difference in myogenic tone observed between arteries from NBCn1 knockout and wild-type mice.
In addition to the reduced overall tone, the amplitude of the oscillatory vasomotor activity, which was observed in a large number of arteries after inhibition of NO synthesis by L-NAME, was strongly attenuated in the arteries from the NBCn1 knockout mice. Although the physiologic role of vasomotion is not comprehensively understood, it has been shown to modify blood flow and suggested to improve tissue dialysis. 36 Thus, an altered vasomotion pattern may contribute to poor tissue oxygenation during metabolic disturbances and acid–base deregulation.
The finding that intracellular acidification in middle cerebral arteries interferes with the same signaling pathways that are affected in mesenteric arteries of NBCn1 and NHE1 knockout mice2, 4 suggests a general applicability of these findings in the resistance vasculature. Although most proteins, including enzymes and ion channels, are influenced by pH to some extent, certain proteins stand out as particularly pH sensitive. Among the most pH-sensitive enzymes with relevance for vascular function, the activities of the NO synthase2, 17 and the endothelin-converting enzyme 37 are inhibited around 30% to 40% by an acidification of 0.2 to 0.3 pH units magnitude, whereas a similarly sized acidification almost completely abolishes the activity of the phosphofructokinase. 38 We have recently shown that the isolated rho-kinase has a moderate pH sensitivity, 2 and our current and previous findings1, 2, 4 support that pHi-induced changes in rho-kinase activity are of physiologic or pathophysiologic relevance. Pinpointing highly pH-sensitive proteins is important to determine relevant targets that may be responsible for cardiovascular complications associated with systemic acid–base disturbances or of consequence for the development of cardiovascular disease in humans with genetic polymorphisms, 25 or possibly mutations in NBCn1 or other proteins involved in pH regulation.
In conclusion, we show that NBCn1 is responsible for the Na+,HCO3− cotransport in mouse middle cerebral arteries. Knockout of NBCn1 inhibits pHi recovery from intracellular acidification and results in a 0.3-pH-unit acidification at steady state. Our findings support that in middle cerebral arteries from NBCn1 knockout mice NO-mediated signaling and rho-kinase-dependent myogenic tone are reduced. These findings suggest that pHi modifies vascular signaling pathways and vascular responsiveness, and therefore may interfere with autoregulation of cerebral blood flow and capillary pressure in response to altered hemodynamic conditions.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.