
Abstract
Medium chain triglycerides (MCTs) are used to treat neurologic disorders with metabolic impairments, including childhood epilepsy and early Alzheimer's disease. However, the metabolic effects of MCTs in the brain are still unclear. Here, we studied the effects of feeding even and uneven MCTs on brain glucose metabolism in the mouse. Adult mice were fed 35% (calories) of trioctanoin or triheptanoin (the triglycerides of octanoate or heptanoate, respectively) or a matching control diet for 3 weeks. Enzymatic assays and targeted metabolomics by liquid chromatography tandem mass spectrometry were used to quantify metabolites in extracts from the hippocampal formations (HFs). Both oils increased the levels of
INTRODUCTION
Impaired energy metabolism in the brain has a role in many neurologic conditions. Ketogenic diets and even medium chain triglycerides (MCTs) are effective in the treatment of several forms of childhood drug-resistant epilepsy 1 and are currently used in the treatment of early Alzheimer's disease. 2 Uneven MCTs are rare in natural foods, but can be chemically produced from natural oils. They are found to be beneficial in several metabolic disorders, 3 anticonvulsant in four mouse seizure models,4, 5, 6 and improve cognitive function in a mouse Alzheimer's disease model. 7 However, little is known about the effects of even and uneven MCTs on brain energy metabolism.
The majority of energy required by the brain is provided by the oxidation of glucose via glycolysis and the tricarboxylic acid (TCA) cycle. Glucose-derived pyruvate is either metabolized to acetyl-CoA via pyruvate dehydrogenase or is carboxylated by pyruvate carboxylase to provide oxaloacetate, thereby refilling the TCA cycle pool of C4 intermediates (anaplerosis). Thus,
A considerable body of research suggests that glucose is the primary fuel for the brain, and is sufficient to provide all essential brain energy requirements in an individual consuming a regular diet. There is also evidence suggesting that under given conditions, alternate fuels such as medium-chain fatty acids can be used as a fuel for the TCA cycle. Measurements of radioactivity levels after infusion with both [1-11C]- and [1-14C] octanoate have shown that octanoate can readily cross the blood–brain barrier. 8 Further studies have shown that with infusion of [1-14C]- or [1-13C] octanoate at 15% to 25% of caloric requirements, the labeled carbon appears predominantly in glutamine as compared with glutamate. As glutamine is predominantly found in astrocytes, this suggests that medium-chain fatty acids are primarily metabolized by astrocytes.9, 10 In cultures of developing rat brain cells, astrocytes are the only cells capable of metabolizing octanoate through respiration to produce carbon dioxide. 11 Although these experimental settings were artificial, they provide evidence that when feeding >15% to 50% of calories as medium chain fats, the brain has the capacity to use them as energy substrates in the presence of normal blood glucose. Also, feeding oral octanoate at doses of 10 to 30 mmol/kg was anticonvulsant in several models, which is most easily explained by diffusion into the brain. 12 However, even-chain fatty acids such as octanoate can only supply two carbon atoms to the TCA cycle. In contrast, odd-chain fatty acids such as heptanoate can provide the two and three carbon fragments, acetyl-CoA and propionyl-CoA, to the cycle.
Triheptanoin, the triglyceride of the seven carbon-chain fatty acid heptanoate, provides the TCA cycle with both acetyl-CoA and anaplerotic propionyl-CoA. Propionyl-CoA is carboxylated to methyl-malonyl-CoA, which is then metabolized into succinyl-CoA, thereby refilling the C4 TCA cycle intermediate pool. Several studies have shown that triheptanoin can improve cardiac and skeletal muscle function in some long-chain fatty acid oxidation disorders.7, 13, 14 In these reports, a greater improvement was observed with triheptanoin than with even-chain fatty acids. The efficacy and potency of even versus uneven MCTs in the treatment of neurologic disorders is still unexplored; however, both are effective in animal models of seizures. Triheptanoin has been found to be anticonvulsant in four rodent seizure models.4, 5, 6 The exact mechanisms of the anticonvulsant effects are still unknown for even and uneven MCTs. In the present study, we compared the effects chronic trioctanoin and triheptanoin feeding on glucose and TCA cycle metabolism in the mouse hippocampal formation (HF) and the 6 Hz seizure test.
MATERIALS AND METHODS
Animals and Diets
Male CD1 mice (Australian Research Council, Western Australia, Australia) were individually caged under a 12-hour light–dark cycle where food and water were given
All diets were matched in protein, mineral, antioxidant, and vitamin content relative to their caloric densities. Standard diet (SF11-027) or matched diets (SF11-079) in which 35% of the calories are from either trioctanoin or triheptanoin oil 6 were obtained from Specialty Feeds (Western Australia, Australia). Triheptanoin was a gift from Sasol GmbH (Germany) and trioctanoin from Stepan Lipid Nutrition (Maywood, NJ, USA). The oils replaced the sucrose, some of the complex carbohydrates and long-chain fats in the standard diet.
Animals for Metabolite Analysis
The animals used for metabolite analysis were from a larger cohort used to investigate the mouse pilocarpine status epilepticus model of temporal lobe epilepsy. The mice used were injected with methylscopolamine (2 mg/kg intraperitoneally in 0.9% NaCl; Sigma-Aldrich, St Louis, MO, USA) and 15 minutes later pilocarpine (330 to 345 mg/kg subcutaneously in 0.9% saline; Sigma-Aldrich). After a 90-minute observation period, mice were injected with pentobarbital (22.5 mg/kg intraperitoneally in 0.9% NaCl; Provet, Northgate, Queensland, Australia) to stop status epilepticus. Only mice that did not develop status epilepticus were used for metabolic analysis. Those mice are known to behave normally and show no brain abnormalities.15, 16 At 7 to 8 weeks old, and immediately after pilocarpine treatment, mice were initiated on either standard diet or matched diets in which 35% of the calories are from either trioctanoin or triheptanoin oil. Mice were maintained on these diets for three weeks before being killed.
Tissue Extraction
Three weeks after the initiation of feeding of the different diets, the mice were killed by focal microwave fixation to the head at 5 kW for 0.80 to 0.87 seconds, denaturing enzymes and other proteins instantaneously (Model MMW-05, Muromachi, Tokyo, Japan). Afterwards, the mice were decapitated and the cerebral cortices and HF were dissected and stored at −80°C until extracted. All HF samples were sonicated in 0.2 mL of methanol using a Vibra Cell sonicator (Model VCX 750, Sonics and Materials, Newton, CT, USA). Polar metabolites were extracted from samples using a modified Bligh-Dyer water/methanol/chloroform extraction procedure at a 1/2/3 ratio, as previously described. 17 Samples were lyophilized and liquid chromatography tandem mass spectrometry analysis was performed by investigators masked to the treatment. Metabolite concentrations are within reported ranges18, 19 and the average coefficient of variation of metabolite levels was 21%, which is expected for biologic samples, despite no internal standard being used to monitor extraction efficiency.
Liquid Chromatography Tandem Mass Spectrometry Analysis
Reference standards and tributylamine (puriss plus grade) were purchased from Sigma-Aldrich (Sigma Aldrich, NSW, Australia). HPLC Grade acetonitrile and acetic acid (AR Grade) were purchased from RCI Labscan (Bangkok, Thailand) and Labscan (Gliwice, Poland), respectively. De-ionized water was generated via an Elga Purelab Classic water purification unit (Veolia Water Solutions and Technologies, Saint Maurice Cedex, France).
Liquid chromatography tandem mass spectrometry data were acquired on a Dionex UltiMate 3000 liquid chromatography system (Dionex, Sunnyvale, CA, USA) coupled to an ABSciex 4000 QTRAP mass spectrometer (ABSciex, Concord, ON, Canada). The liquid chromatography system was controlled by Chromeleon software (Dionex) and chromatographic separation was achieved by injecting 10
Mobile phase gradient profile
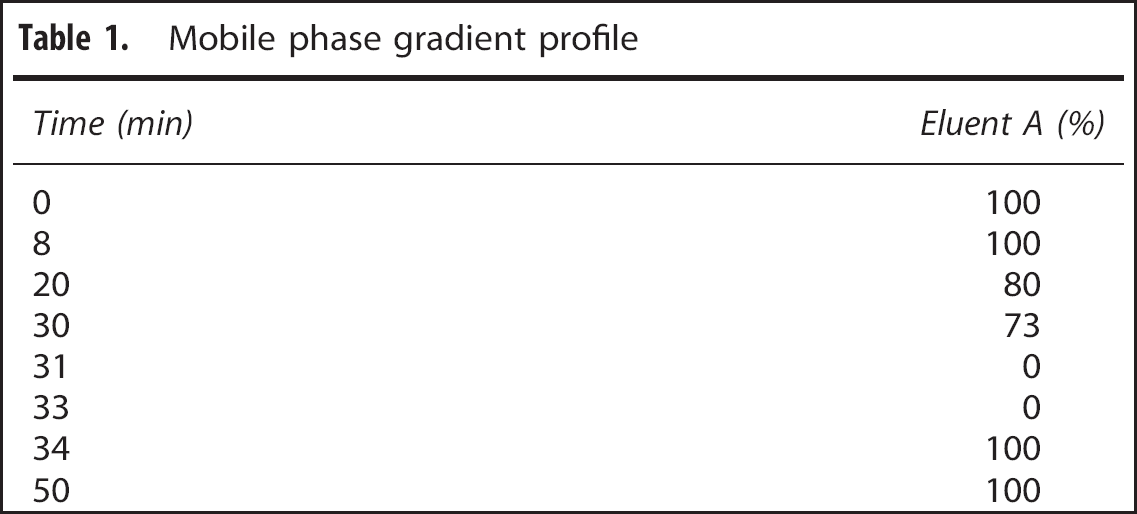
The mass spectrometer was controlled by Analyst 1.5.2 software (ABSciex) and was equipped with a TurboV electrospray source operated in negative ion mode. The following optimized parameters were used to acquire scheduled multiple reaction monitoring data: ionspray voltage−4,500 V, nebulizer (GS1), auxiliary (GS2), curtain (CUR), and collision (CAD) gases were 60, 60, 20, and medium (arbitrary units), respectively, generated via a N300DR nitrogen generator (Peak Scientific, Billerica, MA, USA). The auxiliary gas temperature was maintained at 350°C. The analyte-dependent parameters for the detection of central carbon metabolites are given in Table 2. For all analytes, the entrance potential (EP) was −10 volts.
Analyte-dependent parameters for the transitions used in scheduled multiple reaction monitoring data acquisition
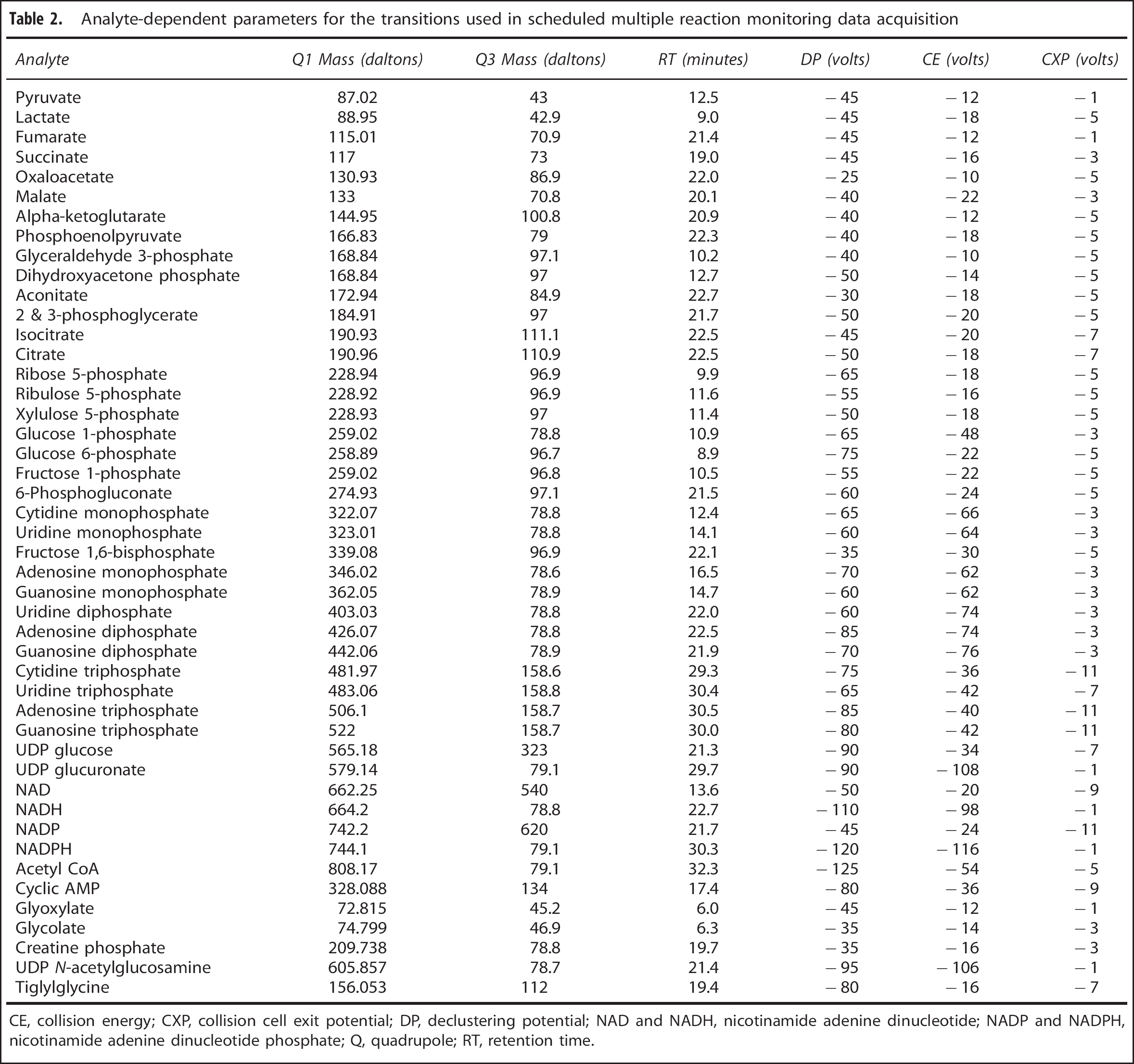
CE, collision energy; CXP, collision cell exit potential; DP, declustering potential; NAD and NADH, nicotinamide adenine dinucleotide; NADP and NADPH, nicotinamide adenine dinucleotide phosphate; Q, quadrupole; RT, retention time.
The samples were run with sample- and analyte-relevant calibration standards and pooled quality control samples21, 22 to control for reproducibility of data acquisition and to ensure data integrity. Analyte stock solutions were prepared in purified water (Veolia) and aliquots of each solution were mixed to achieve a final calibrant solution at 400
Enzymatic Metabolite Assays
Glucose and
6 Hz Seizure Test
Fifteen 7-week-old CD1 mice were placed on either standard diet, or diets containing 35% of calories as triheptanoin or trioctanoin. The 6 Hz seizure model was performed as previously described 20 on the same mice on days 2, 7, and 21 after diet initiation. The critical current required to induce seizures in 50% of mice (CC50) was determined using the ‘up and down method’ varying electrical current intensities at 2-mA intervals. A topical anesthetic (0.5% tetracaine hydrochloride ophthalmic solution) was applied to corneas 15 minutes before a 3-seconds corneal stimulation with 0.2 millisecond-duration pulses at 6 Hz (ECT Unit 57800, Ugo Basile). Mice were held manually during stimulation and released to observe behavioral responses. Seizures were classified as a stunned or fixed posture, rearing, forelimb clonus, or twitching.
Data Analysis
The levels of each metabolite were normalized to the weight of extracted tissue in grams. All statistics were performed by GraphPad Prism software (La Jolla, CA, USA) using a one-way analysis of variance, with a Newman–Keuls
RESULTS
Body Weights and Brain Levels of Glucose and Ketones
The changes in body weights were similar to control diet and found to not be significantly different when feeding 35% trioctanoin or triheptanoin diets for up to 21 days (Figure 1). To assess the effects of the ingestion of the even and odd carbon length MCTs trioctanoin and triheptanoin on metabolic pathways in the mouse brain, concentrations of metabolites were analyzed in HF extracts through liquid chromatography tandem mass spectrometry and enzymatic assays.
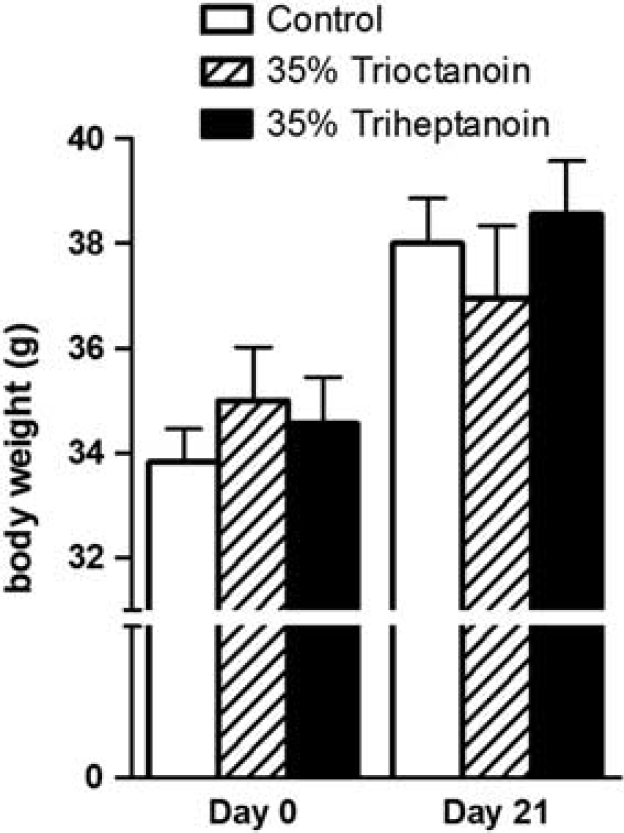
Dietary medium chain triglycerides do not alter body weight in mice. Body weights of CD1 mice, before diet induction and after 3 weeks of feeding a control (white bars,
The levels of glucose were similar in HF extracts of mice in each treatment group (Table 3). In contrast, the levels of BHB were significantly higher in animals that were fed MCTs, specifically 1.7-fold higher for triheptanoin (
Glucose metabolism
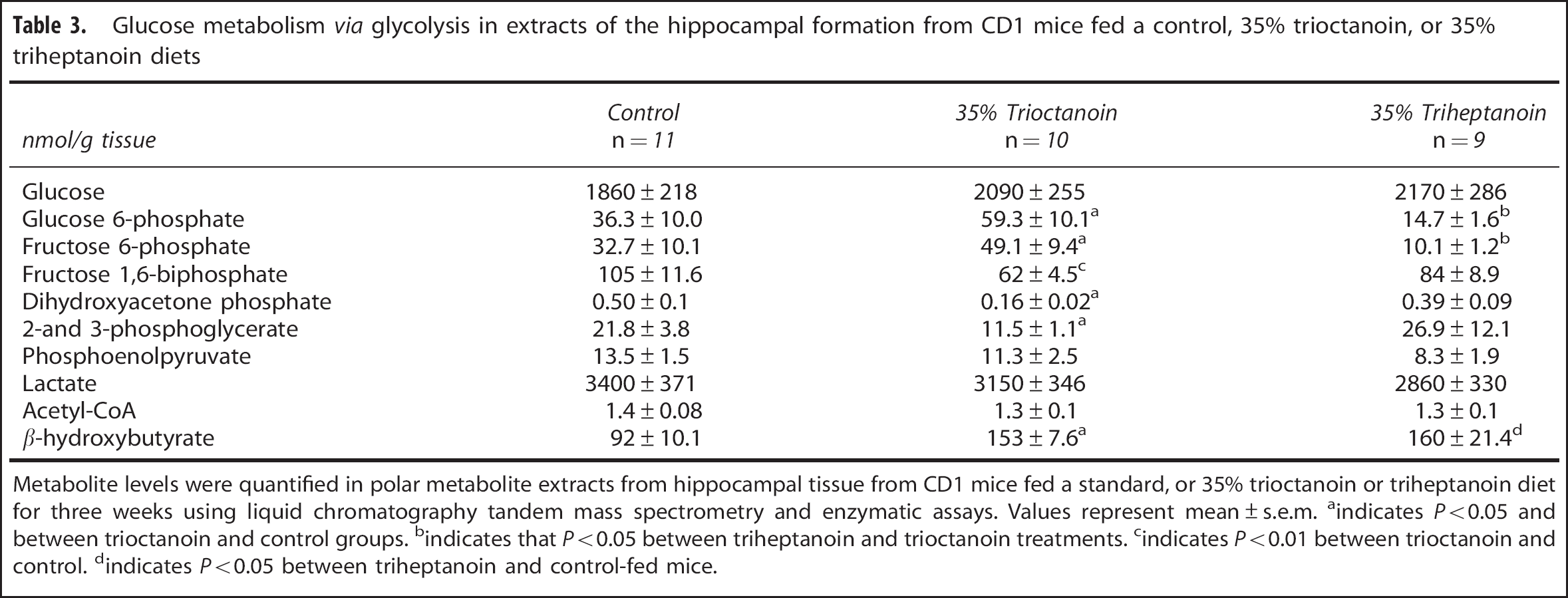
Metabolite levels were quantified in polar metabolite extracts from hippocampal tissue from CD1 mice fed a standard, or 35% trioctanoin or triheptanoin diet for three weeks using liquid chromatography tandem mass spectrometry and enzymatic assays. Values represent mean±s.e.m.
indicates
indicates that
indicates
indicates
Metabolism of Glucose Via Glycolysis
The levels of several metabolites produced through the metabolism of glucose via the glycolytic pathway were measured (Table 3). In mice fed a diet where 35% calories are supplied from trioctanoin, a 1.6- (
Metabolites of the Pentose Phosphate Pathway
Xylulose 5-phosphate, ribulose 6-phosphate, and 6-phosphogluconate, three intermediates of the pentose phosphate pathway (PPP) were quantified in the HF (Figure 2). In the HF extracts of mice fed trioctanoin, the concentration of ribulose 5-phosphate was reduced by 60% (
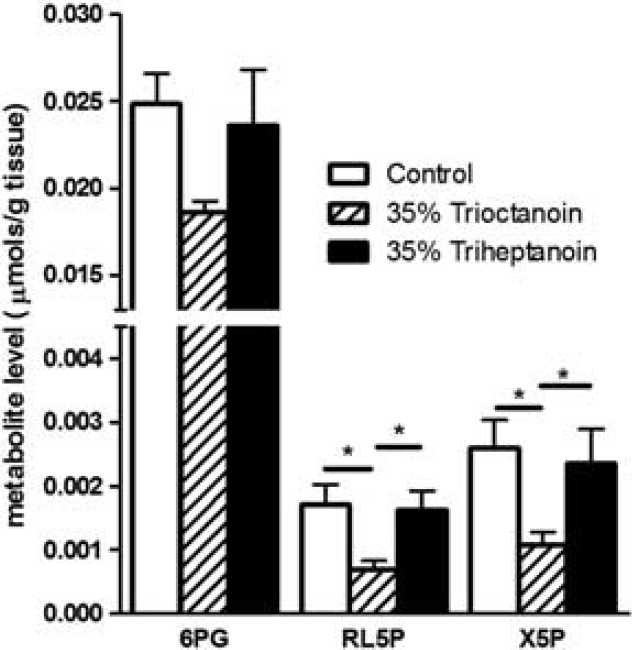
Alterations of levels of the pentose phosphate pathway metabolites after feeding medium-chain triglycerides. Metabolite levels of 6-phosphogluconate (6PG), ribulose 5-phosphate (RL5P), and xylulose 5-phosphate in hippocampal formation brain extracts from mice fed control (white bars,
Tricarboxylic Acid Cycle Intermediates and High-Energy Metabolites
Few changes were observed in the concentrations of high-energy metabolites after even and uneven MCT feeding (Table 4). Levels of ADP were lowered by 17% in mice fed triheptanoin with respect to those fed a control diet (
High-energy metabolite and tricarboxylic acid cycle metabolite levels in CD1 HF extracts from mice fed a control, 35% trioctanoin, or 35% triheptanoin diet
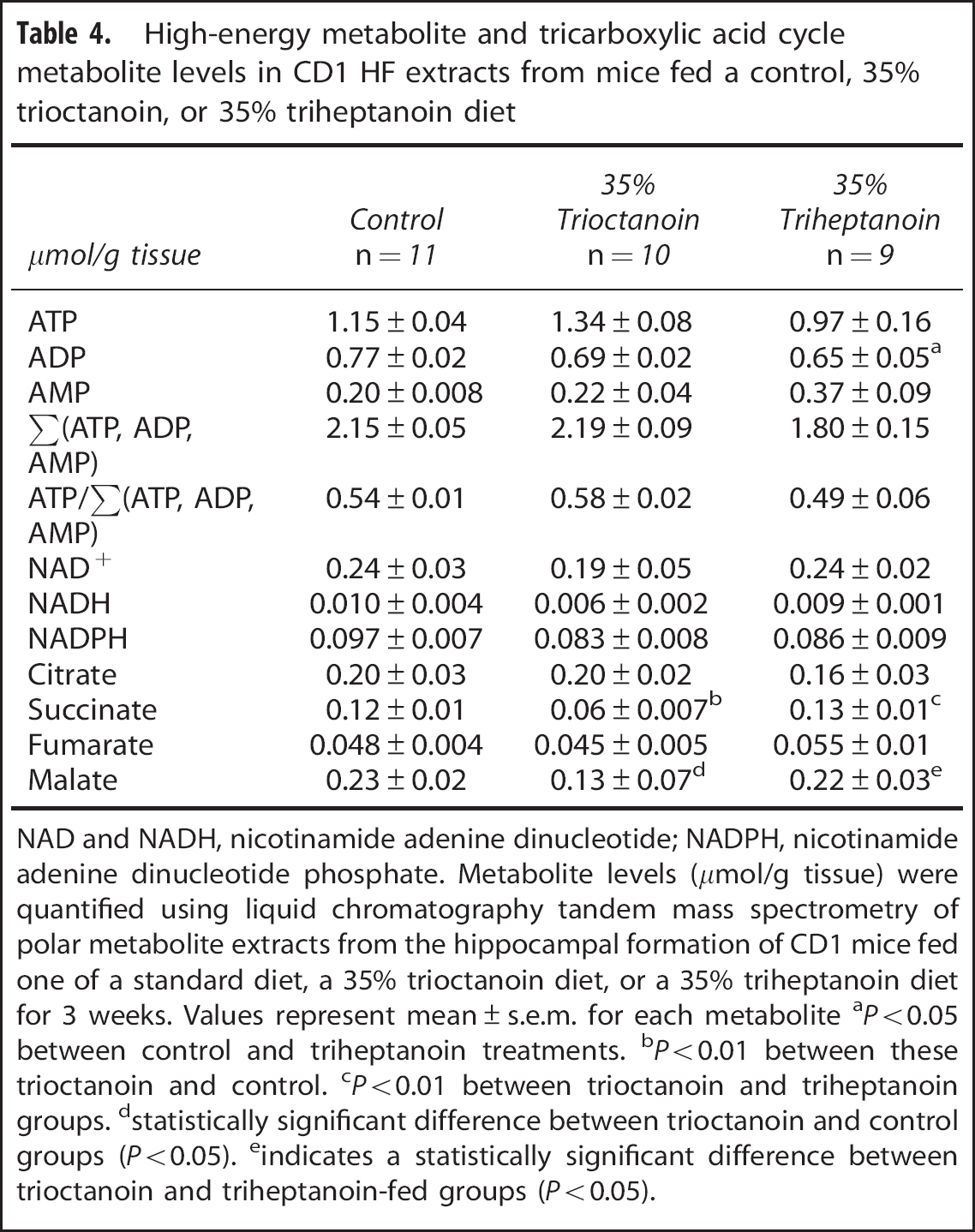
NAD and NADH, nicotinamide adenine dinucleotide; NADPH, nicotinamide adenine dinucleotide phosphate.
Metabolite levels (
statistically significant difference between trioctanoin and control groups (
indicates a statistically significant difference between trioctanoin and triheptanoin-fed groups (
Seizure Susceptibility
The body weights of mice between each treatment group were similar at each given time point (Figure 3A). The 6 Hz test was used to compare the effects of triheptanoin versus trioctanoin on seizure susceptibility in mice after feeding these oils for 2, 7, and 21 days (Figure 3B). After two days of feeding, trioctanoin-fed mice showed a higher seizure threshold by 3.41 mA relative to triheptanoin (
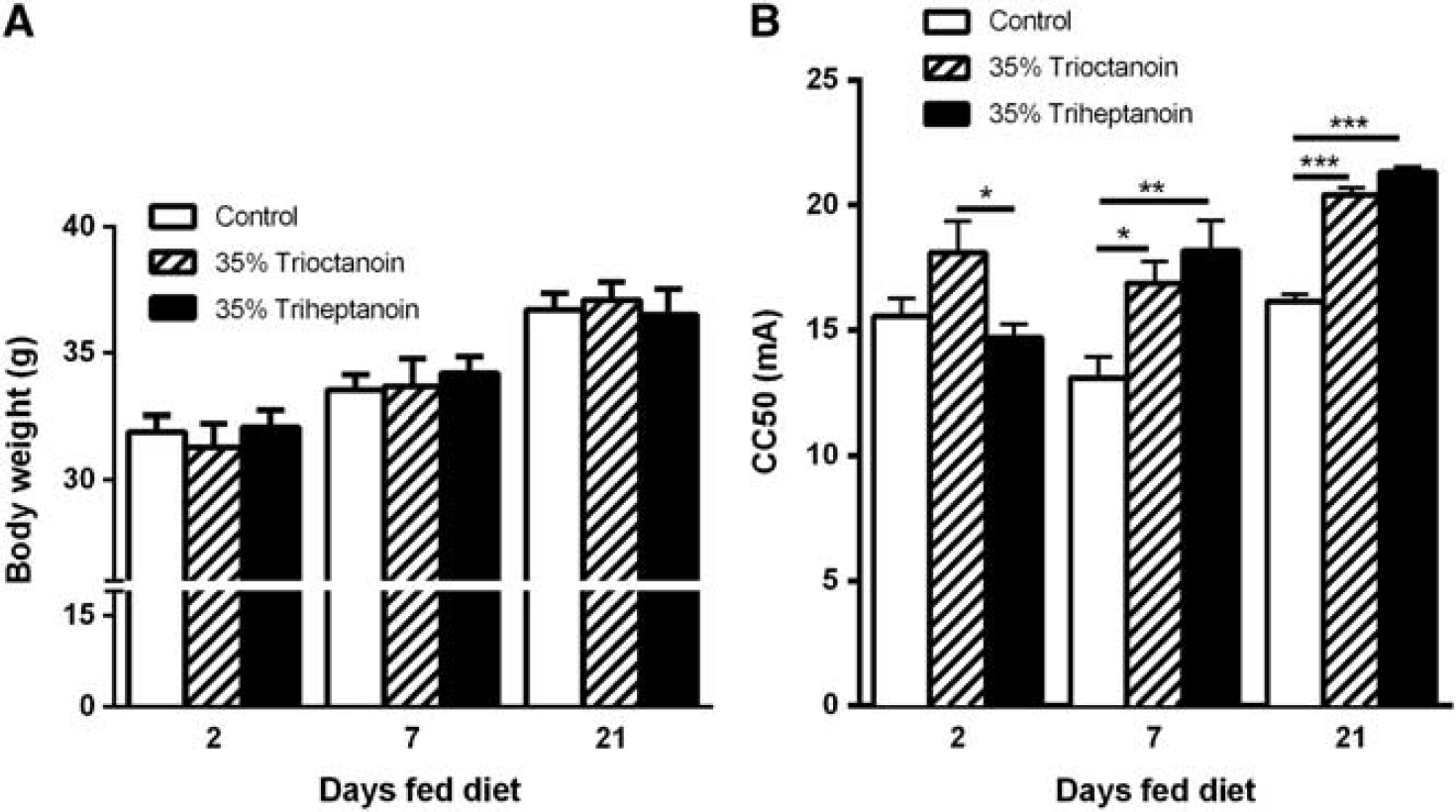
Similar anticonvulsant effects with even and uneven medium-chain triglycerides in the 6 Hz model. (
DISCUSSION
Reduction of Glucose Utilization with Trioctanoin Feeding
The various pathways of glucose metabolism were analyzed through the determination of the concentration of intermediates in extracts of the HF in CD1 male mice that were fed
Triheptanoin does not Appear to Alter Glucose Metabolism
Levels of intermediates of glucose and TCA cycle metabolism were largely unaltered by triheptanoin in the hippocampus of mice without neurologic problems in this and previous studies.
4
Heptanoate is
Reduction of Phosphofructokinase Activity by Trioctanoin
A key finding of this study was the loss of fructose 1,6-bisphosphate and downstream metabolites of the glycolytic pathway in mice fed trioctanoin. This coupled with the findings that fructose 6-phosphate and glucose 6-phosphate levels were increased, indicates that phosphofructokinase activity is inhibited in the HF in mice that were fed trioctanoin but not triheptanoin. This is consistent with previous studies showing that both octanoate and ketone bodies inhibit the activity of phosphofructokinase both
One proposed mechanism for this inhibition of phosphofructokinase is through the accumulation of two negative regulators, acetyl-CoA and citrate, that are produced through the
Even-Chain Fatty Acids Alter Metabolism Through the Pentose Phosphate Pathway
In the HF, the reduction in xylulose 5-phosphate levels coincided with a decrease in ribulose 5-phosphate levels in mice fed trioctanoin. Similar to the glycolytic intermediates, these reductions were not observed with triheptanoin. These findings can be interpreted either as a (1) decrease or (2) increase of the PPP activity with even MCT feeding. (1) Glucose 6-phosphate dehydrogenase, which catalyzes the reaction of glucose 6-phosphate to 6-phosphogluconolactone, is the limiting reaction in the PPP pathway. There is evidence that diets high in polyunsaturated fatty acids limit the synthesis of this enzyme,33, 34 which would lead to a decrease in PPP activity. Little is known of the effect of MCT on brain PPP metabolism. In light of the data from this study, it will be of interest to investigate the effect of medium-chain fatty acids on glucose 6-phosphate dehydrogenase. (2) Alternatively, the loss of PPP intermediates could reflect an increase in PPP activity to compensate for the reduction of glycolytic activity to facilitate the metabolism of glucose. Citrate levels increase acutely after the application of octanoate in guinea pig hearts. 35 This initial increase was associated with the inhibition of phosphofructokinase activity, reducing the production of downstream glycolytic intermediates, such as glyceraldehyde-3-phosphate, which is also a product of the PPP. We did not observe increases in citrate levels in our steady-state measurements. However, it is possible that it was missed because of fast turnover rate of TCA cycling. The loss of glyceraldehyde 3-phosphate may drive the reactions by ribulose 5-phosphate isomerase and transketolase to completion, which would explain the observed loss in xylulose 5-phosphate and ribulose 5-phosphate by an increase in PPP activity (Figure 4).
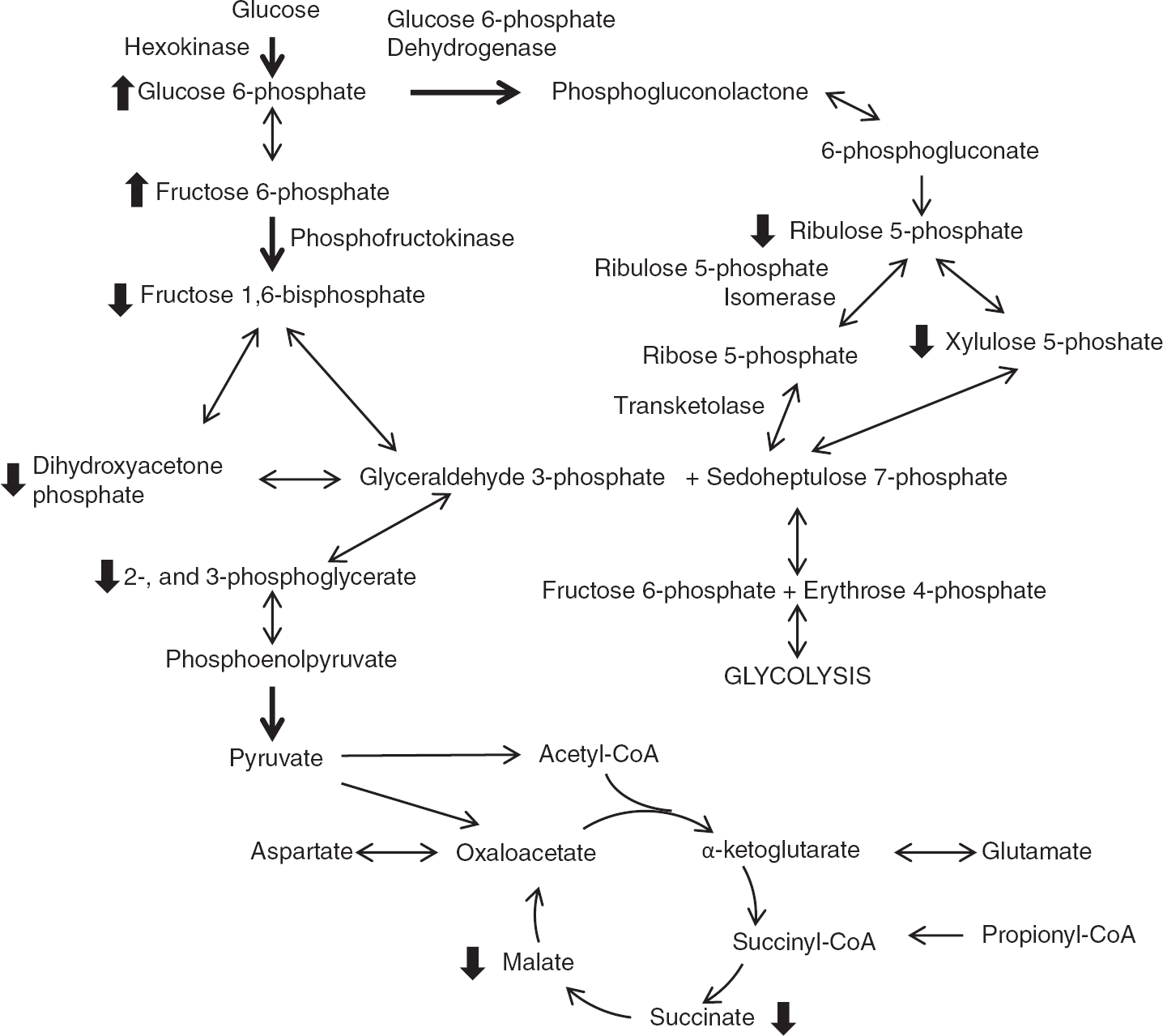
Simplified schematic representation of the alterations within glucose metabolism pathways in the brain after feeding of medium-chain triglycerides. The metabolism of glucose by glycolysis and the pentose phosphate pathway is shown, along with the TCA cycle. Dark arrows represent regulatory steps in the pathway. Only the enzymes involved in reactions where changes in metabolite levels were observed are labeled. Thick arrows next to metabolites indicate significant differences between trioctanoin- and control diet-fed mice in the direction of arrow.
Levels of Tricarboxylic Acid Cycle Intermediates and Adenosine Nucleotides after Medium-Chain Triglyceride Feeding
Feeding of trioctanoin reduced the levels of two of the intermediates in the TCA cycle, succinate and malate, which suggests that the TCA cycling is altered. However, as only static levels of metabolites were measured, we were unable to determine whether this is caused by an increase in flux because of increased acetyl-CoA entry via citrate synthase or a reduction in the cycling resulting from a loss of glucose utilization. Triheptanoin did not result in a reduction of these intermediates levels, which correlates with previous studies in mice fed a 35% triheptanoin diet for 3 weeks. 4 Triheptanoin may restore the levels of the TCA cycle intermediates through the supply of succinyl-CoA through the anaplerotic propionyl-CoA pathway. In addition, as previously stated, triheptanoin appears not to inhibit glycolysis, and thus the flux through the TCA cycle may not be altered in these mice.
Levels of ADP were lowered by 17% in mice fed triheptanoin. This was surprising, because it was not observed in a previous study investigating the metabolism of 13C-glucose after triheptanoin feeding in the same mouse strain (data unpublished). In both studies, the total amount of all adenosine nucleotides and the ratio of ATP against all adenosine nucleotides remained unchanged as compared with control-fed mice.
Anticonvulsant Effects of Even versus Uneven Medium-Chain Triglycerides and Mechanism of Anticonvulsant Effects
Despite the differences in the metabolite concentrations in the HF after trioctanoin versus triheptanoin feeding in this study, both MCTs showed similar anticonvulsant effects in the acute 6 Hz seizure model. These anticonvulsant effects are similar in magnitude to those found after ketogenic diet feeding in this model. 20 In a previous study of CD1 mice, no consistent anticonvulsant effects of triheptanoin were observed in the 6 Hz model. 6 A possible explanation for this discrepancy is that CD1 mice used in previous studies were from a local mouse colony; however, the mice used in this study obtained from a different colony at the Australian Research Council, which is genetically slightly different. As we do not have a comparison of efficacy of trioctanoin versus triheptanoin versus ketogenic diet in several seizure models, it is difficult to speculate on the anticonvulsant mechanism(s) of these dietary treatments. We concluded earlier that the anticonvulsant mechanism(s) of the ketogenic diet in mice may involve a reduction of glucose utilization as fuel. 20 The anticonvulsant actions of 2-deoxyglucose and fructose 1,6-bisphosphate found in animal models36, 37, 38 also suggest that reduced glycolysis and/or increased PPP can be anticonvulsant. As trioctanoin appears to alter glucose utilization through glycolysis and the PPP, it is possible that these metabolic alterations are involved in increasing the seizure threshold in naïve mice in this model with trioctanoin but not triheptanoin. Also, it is possible that the anticonvulsant mechanisms observed by both MCTs in this model are similar and could be explained by anticonvulsant effects of BHB. Similar BHB levels were reached in the HF by both oils to those achieved by the ketogenic diet in a similar mouse strain. 39 Furthermore, this would suggest that the anticonvulsant effects in the 6 Hz model are independent of the possible anaplerotic effects of triheptanoin. However, the anaplerotic potential of triheptanoin is expected to add advantages in chronic disease models with metabolic disturbances, and in clinical settings.3, 4, 13, 14
Previously, it has been hypothesized that anaplerosis is an advantage, specifically to the ‘epileptic’ brain in which TCA cycle intermediates levels are reduced.4, 19 So far, to our knowledge trioctanoin has not been tested in chronic epilepsy models. From this study, one could expect that trioctanoin may be less effective than triheptanoin in chronic epilepsy models because trioctanoin reduces the levels of TCA cycle intermediates and is not anaplerotic. As discussed above, the reduced PPP metabolite levels found after trioctanoin feeding are difficult to interpret. However, if they are an indication of increased flux through the PPP, this would permit glucose catabolism, resulting in increased production of NADPH, which increases the potential to reduce glutathione disulfide by glutathione reductase, and allows production of anaplerotic pyruvate. Although we did not find an increase in the NADPH levels with trioctanoin, the conclusions from these data are limited, as NADP+ levels were below detectable levels. Increased flux through the PPP is a proposed mechanism for the anticonvulsant effect observed by fructose-1,6-bisphosphate. 40
CONCLUSION
In comparison with trioctanoin, triheptanoin feeding had only few significant effects on metabolite levels in the HF, predicting few metabolic side effects for triheptanoin. Both MCTs showed similar anticonvulsant effects in healthy brains, thus the anticonvulsant effects appear to be independent of changes in glucose metabolism. The HF metabolite level changes found with trioctanoin raise several questions, mostly in terms of alteration in PPP activity and its effects and mechanism of action in models of chronic epilepsy and Alzheimer's disease.
Footnotes
KB has filed for a complete US patent regarding triheptanoin as a treatment for seizures. No other authors have any interests/conflicts to disclose.
ACKNOWLEDGEMENTS
The authors are grateful to Mussie Hadera for comments on the manuscript and funding by the Australian National Health and Research Council (Grants 63145 and 104407 to KB). We thank Sasol and Stepan Lipid Nutrition for providing triheptanoin and trioctanoin in kind.