
Abstract
The adaptation of the cerebral circulation to pregnancy is unique from other vascular beds. Most notably, the growth and vasodilatory response to high levels of circulating growth factors and cytokines that promote substantial hemodynamic changes in other vascular beds is limited in the cerebral circulation. This is accomplished through several mechanisms, including downregulation of key receptors and transcription factors, and production of circulating factors that counteract the vasodilatory effects of vascular endothelial growth factor (VEGF) and placental growth factor. Pregnancy both prevents and reverses hypertensive inward remodeling of cerebral arteries, possibly through downregulation of the angiotensin type 1 receptor. The blood–brain barrier (BBB) importantly adapts to pregnancy by preventing the passage of seizure provoking serum into the brain and limiting the permeability effects of VEGF that is more highly expressed in cerebral vasculature during pregnancy. While the adaptation of the cerebral circulation to pregnancy provides for relatively normal cerebral blood flow and BBB properties in the face of substantial cardiovascular changes and high levels of circulating factors, under pathologic conditions, these adaptations appear to promote greater brain injury, including edema formation during acute hypertension, and greater sensitivity to bacterial endotoxin.
INTRODUCTION
The adaptation of the maternal cardiovascular system to pregnancy is substantial and necessary for normal growth and development of the fetal-placental unit.1,2 Systemically, pregnancy is a high volume, low resistance state characterized by a large increase in cardiac output (CO) driven by circulating hormones that rise throughout the course of gestation.1–4 Plasma volume increases 40% to 50% during singleton gestation (and up to 100% for twin gestation) that is associated with a drop in systemic vascular resistance.1–4 As a consequence, blood pressure declines mid-gestation then rises to normal levels near term.2,4 In addition to systemic cardiovascular changes, several organs have substantial increases in blood flow during pregnancy, including the kidneys, ovaries, and uterus.5,6 The distribution of CO to individual organs also changes over gestation with the greatest increase to the uteroplacental circulation. 6
The adaptation of the brain and cerebral circulation to pregnancy is importantly unique from other organs because of the need for a constant blood supply and the relative intolerance to increased blood volume. The progressive demand on the cardiovascular system and high levels of circulating factors that rise over the course of pregnancy poses challenges for the brain, an organ with high metabolic requirements and the need for tight water and ion homeostasis. The normal physiological adaptation of the cardiovascular system to pregnancy includes changes that affect vessel wall integrity, hemodynamics, and coagulation that increase the risk of stroke, edema, and other neurovascular complications.1–4,7–9 Compared with other organs, we have a limited understanding of the adaptation of the cerebral circulation to pregnancy and the underlying mechanisms that drive it. However, understanding how normal pregnancy, and conditions such as preeclampsia, affect the cerebrovascular wall is important considering neurologic complications are a leading cause of maternal morbidity and mortality worldwide. 10
This review will describe current knowledge about the adaptation of the cerebral circulation to pregnancy, including changes in cerebral blood flow (CBF), hemodynamics, vascular structure, and blood–brain barrier (BBB) properties. Although both human and animal studies are discussed, it is worth noting that human studies on brain blood flow and cerebrovascular structure are difficult to perform and thus robust information on the adaptation of the cerebral circulation in humans is lacking. We have used animal models to characterize the adaptation of the cerebral circulation to pregnancy and these will be described. In addition, how the adaptation of the cerebral circulation during pregnancy may predispose to neurologic symptoms, especially during pathologic states such as preeclampsia and eclampsia, is also discussed.
CHANGES IN CEREBRAL BLOOD FLOW AND AUTOREGULATION DURING NORMAL PREGNANCY
Measurement of Cerebral Blood Flow and Cerebrovascular Resistance During Pregnancy
The substantial increase in plasma volume and CO during pregnancy is distributed unequally to several organ systems. Most notably, uterine blood flow increases 10-fold over the nonpregnant state by late-gestation, with the percent of CO received by the uteroplacental unit increasing from ~0.5% to 15%.5,6 Undoubtedly, such a dramatic increase in CBF cannot be tolerated by the brain. However, the magnitude by which CBF does change during pregnancy, if at all, has been difficult to assess in humans. Transcranial Doppler (TCD) ultrasound has been widely used to study cerebral hemodynamics during human pregnancy because it is noninvasive and can measure changes in blood flow velocity.11–15 However, it cannot measure vessel diameter and thus the validity of extrapolating CBF from TCD measurements is of concern.
16
One recent crosssectional study used dual-beam angle-independent digital ultrasound to measure blood flow changes in the internal carotid artery (ICA) over the course of pregnancy in healthy women.
17
This study also measured ICA diameter and thus calculated changes in cerebrovascular resistance (CVR) and global CBF. In this study, CVR decreased from a nonpregnant value of 0.141 to 0.112 mm Hg × mL/100 g/min in the third trimester, with CBF increasing 22% from 42.2 mL/100 g/min in nonpregnant women to 51.8 mL/100 g/min in the third trimester. The strength of this study is that ICA diameter and blood flow volume were measured; however, it is limited by a crosssectional analysis and that there were eightfold more patients measured in the third trimester compared with nonpregnant, and fivefold more than in the first trimester. In addition, how the authors normalized to brain weight is unclear. The finding that CBF increases 22% by late-gestation is in contrast to a longitudinal study by Zeeman
We and others have used microspheres in animals to measure absolute changes in CBF during pregnancy.6,19,20 There are obvious limitations to animal studies as well, including the use of anesthesia in some19,20 but not all 6 studies that can affect CVR and CBF. Further, the use of microspheres is terminal, thus precluding longitudinal studies. In unanesthetized, standing sheep instrumented with indwelling catheters, CBF was found to decrease from 48 mL/100 g/min in the nonpregnant state to 38 mL/100 g/min by late-pregnancy (130 to 140 days). 6 Other studies in anesthetized rats found little change in CBF at late-gestation compared with nonpregnant: 92 versus 88 mL/100 g/min 19 and 58 versus 60 mL/100 g/min. 20
Measurement of Autoregulation of Cerebral Blood Flow During Pregnancy
Autoregulation of CBF is an important mechanism that provides relatively constant blood supply during changes in perfusion pressure. It is a highly protective mechanism in the brain that has limits. In normotensive adults, CBF is ~50 mL/100 g/min provided that cerebral perfusion pressure is between ~60 and 160 mm Hg.21–23 Above and below these limits, CBF becomes dependent on perfusion pressure linearly. During normal pregnancy, cerebral autoregulation appears to be intact and similar to nonpregnant women, as assessed by transient hyperemic response and TCD. 24 However, whether or not the limits of autoregulation are shifted during human pregnancy is not known, but important to understand considering both hypertensive and hypotensive episodes occur frequently in pregnant women. For example, hypertension is one of the most common complications of pregnancy. 25 If the upper limit of autoregulation is shifted to lower pressures during pregnancy, autoregulatory breakthrough would occur at lower pressures, potentially causing vasogenic edema. In fact, edema formation in response to autoregulatory breakthrough has been proposed as an underlying mechanism of eclampsia.10,26,27 The lower limit of CBF autoregulation is also important to understand because substantial hemorrhage occurs during parturition often lowering blood pressure. 28 If the lower limit of autoregulation is shifted to higher pressures, CBF may fall with decreasing pressure, leading to neurological symptoms such as dizziness, confusion, loss of consciousness, and ultimately ischemic brain damage.29–31
We have measured the limits of CBF autoregulation during normal pregnancy in anesthetized rats using laser Doppler to measure changes in CBF. Using pentobarbital as an anesthetic with acute infusion of phenylephrine to raise blood pressure, we found no difference in the pressure at which autoregulatory breakthrough occurred between nonpregnant and late-pregnant rats. 32 However, because laser Doppler measures relative changes in CBF, whether or not CBF was at the same level after breakthrough of autoregulation occurred could not be determined. Thus, in a separate study we used microspheres to measure absolute changes in CBF basally before infusion of phenylephrine and then after blood pressure was acutely increased to 203 ± 3 mm Hg for nonpregnant and 193 ± 3 mm Hg for late-pregnant rats. We found that while CBF was similar in late-pregnant versus nonpregnant rats at baseline, there was an ~40% increase in CBF with acute hypertension in the pregnant animals (Figure 1A). 20 The increase in CBF at the higher pressures was due to a decrease in CVR that was greater in the pregnant animals: 0.70 ± 0.07 mm Hg × mL/100 g/min for nonpregnant versus 0.45 ± 0.05 mm Hg × mL/100 g/min for pregnant (Figure 1B). The decreased CVR in pregnant animals with acute hypertension is likely due to increased vascular volume that occurs in pregnancy secondary to outward remodeling of brain arterioles and increased capillary density (see below). Autoregulation of CBF was recently measured in nonpregnant and late-pregnant rats using chloral hydrate anesthesia instead of pentobarbital. 33 It was found that the upper limit of autoregulation is somewhat shifted to higher pressures in late-pregnant rats in both the anterior and posterior cerebral cortices (Figures 2A and 2B). However, the shape of the CBF autoregulatory curves is different with the different anesthesia, likely because chloral hydrate did not produce the same change in CBF, suggesting that there was some decrease in CVR compared with pentobarbital before the start of phenylephrine infusion. Regardless, when brain water content was measured, only the pregnant animals had significant edema formation in response to acute hypertension, similar to pentobarbital anesthesia.20,33 Thus, it appears that the brain is more susceptible to edema formation during pregnancy when there is an acute elevation in blood pressure. This finding is significant considering edema is a primary mechanism by which seizure is thought to occur during hypertensive pregnancy.26,27
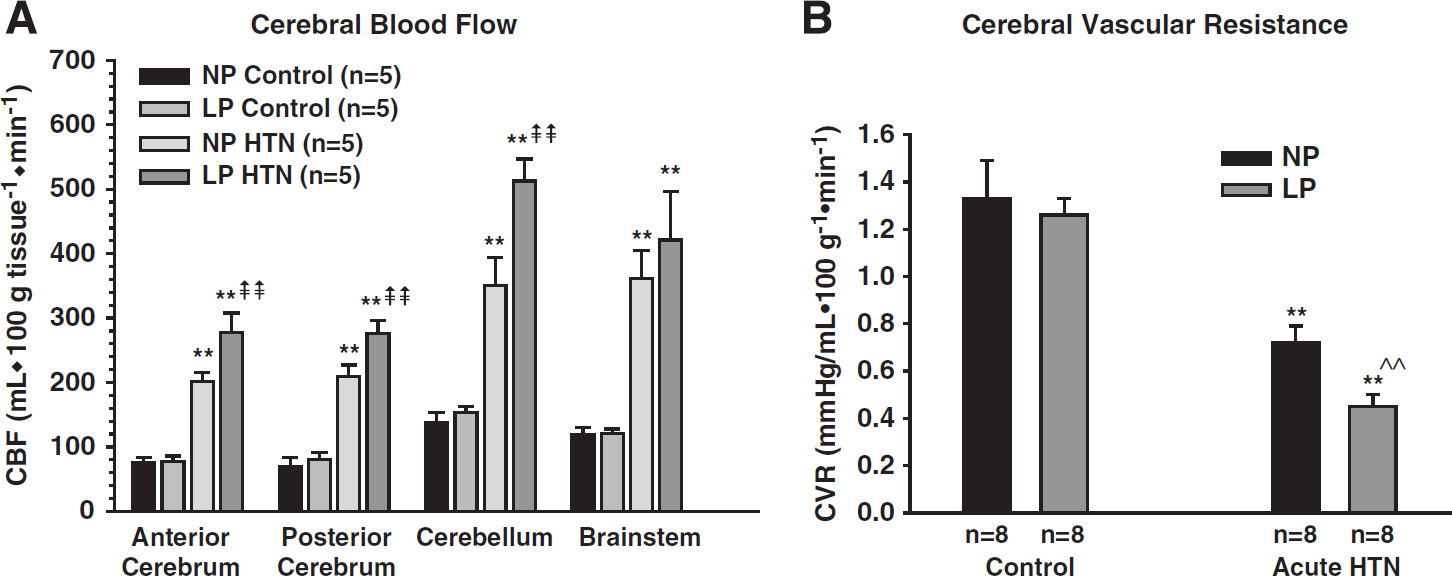
Effect of pregnancy on cerebral blood flow (CBF) and cerebrovascular resistance (CVR) in response to acute hypertension. (
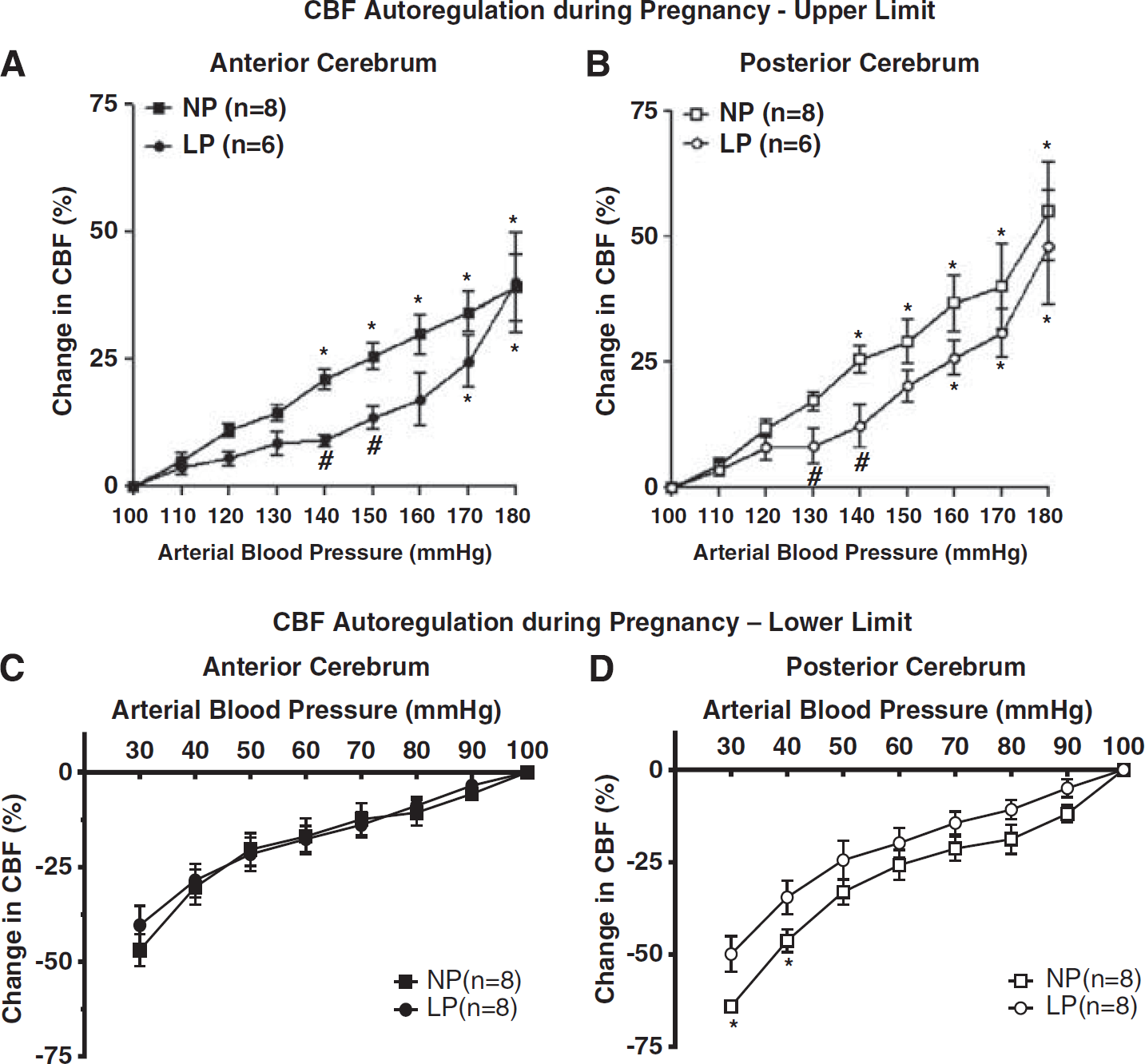
Changes in cerebral blood flow (CBF) autoregulation during pregnancy. (
In addition to shifting the upper limit of CBF autoregulation, pregnancy also appears to shift the lower limit. Cerebral blood flow autoregulation during hemorrhagic hypotension was measured in nonpregnant and late-pregnant rats under chloral hydrate anesthesia. Unlike the upper limit of CBF autoregulation that was shifted in both anterior and posterior cerebral cortices during pregnancy, the lower limit of autoregulation was shifted to lower pressures only in the posterior cerebral cortex (Figures 2C and 2D). The extension of the autoregulatory curve to lower pressures during pregnancy may be a protective mechanism against hypoxia/ischemia during hemorrhagic hypotension that occurs during parturition. The mechanism by which this occurs preferentially in the posterior cortex, or the consequence thereof, is not completely understood. However, gestation-induced changes in endothelial and neuronal nitric oxide synthase (eNOS and nNOS) specifically in the posterior cerebral cortex may have a role. 33
ADAPTATION OF THE CEREBRAL CIRCULATION DURING NORMAL PREGNANCY
Changes in Reactivity and Structure of Posterior Cerebral Arteries Despite considerable changes in renal and uterine artery structure and function during gestation that has significant impact on blood flow to those organs, how pregnancy influences cerebral arteries is less understood. Unlike other organs, large cerebral arteries that lie within the subarachnoid space (pial) are important contributors to vascular resistance in the brain. 35 Because these vessels can influence CVR and CBF, their adaptation to pregnancy is important to understand. In addition, remodeling of cerebral arteries during gestation could impact the integrity of the vascular wall, a consequence that is also important to consider, especially in the presence of comorbidities such as diabetes or hypertension that are known to increase the risk of hemorrhagic stroke.36,37
The majority of studies during pregnancy has been on the PCA, mostly because of the propensity for edema to form in the posterior cortex during conditions such as early-onset preeclampsia, HELLP syndrome (hemolysis, elevated liver enzymes, low platelets) and eclampsia.26,27 Late-pregnancy has been particularly studied because this is when neurologic symptoms occur most often. 38 Posterior cerebral artery structure, including passive diameters and distensibility, is not different in late-pregnant rats, however, they undergo forced dilatation at lower pressures.20,39 Our original interpretation of these results was that the autoregulatory curve was shifted to lower pressures during pregnancy. We now know that this is not the case (see above) and reminds us that myogenic activity of pial arteries is not the only factor influencing autoregulation of CBF. How pregnancy might alter neuronal or metabolic components that influence autoregulation of CBF has yet to be explored.
It is well-known that pregnant animals and humans are more sensitive to bacterial endotoxin than in the nonpregnant state, responding to low-dose lipopolysaccharide (LPS) with disseminated intravascular coagulation.40,41 The hypersensitivity of the peripheral and renal vasculature to LPS is thought to be due to changes in both the innate and adaptive immune systems during pregnancy. 42 Recently, we explored whether the cerebral circulation also has increased sensitivity to LPS by infusing 1.5 μg/kg on day 14 of rat pregnancy and then studying changes in PCA structure and function on day 20 of a 22 day gestation (i.e., late-pregnancy). Compared with similarly treated nonpregnant animals, PCA from late-pregnant animals responded to LPS with decreased tone and increased sensitivity to nitric oxide (Figure 3). 43 In addition, inducible nitric oxide synthase (iNOS) expression was increased threefold in PCA from late-pregnant animals under control conditions (saline-infused). Furthermore, while vessels from both nonpregnant and pregnant animals responded to LPS infusion with an increase in iNOS expression, the increase was greater during pregnancy. When cytokine mRNA expression was measured in pial arteries from nonpregnant and late-pregnant animals, pregnancy alone caused an increase in the expression of the proinflammatory cytokines tumor necrosis factor α (TNFα) and interferon γ but also an increase in the antiinflammatory cytokine interleukin-10. 43 Thus, cerebral arteries during pregnancy appear to be in a state of inflammation, with a substantial increase in iNOS and pro-inflammatory cytokine expression. However, the increase in interleukin-10 may provide an antiinflammatory balance such that function remains near normal.
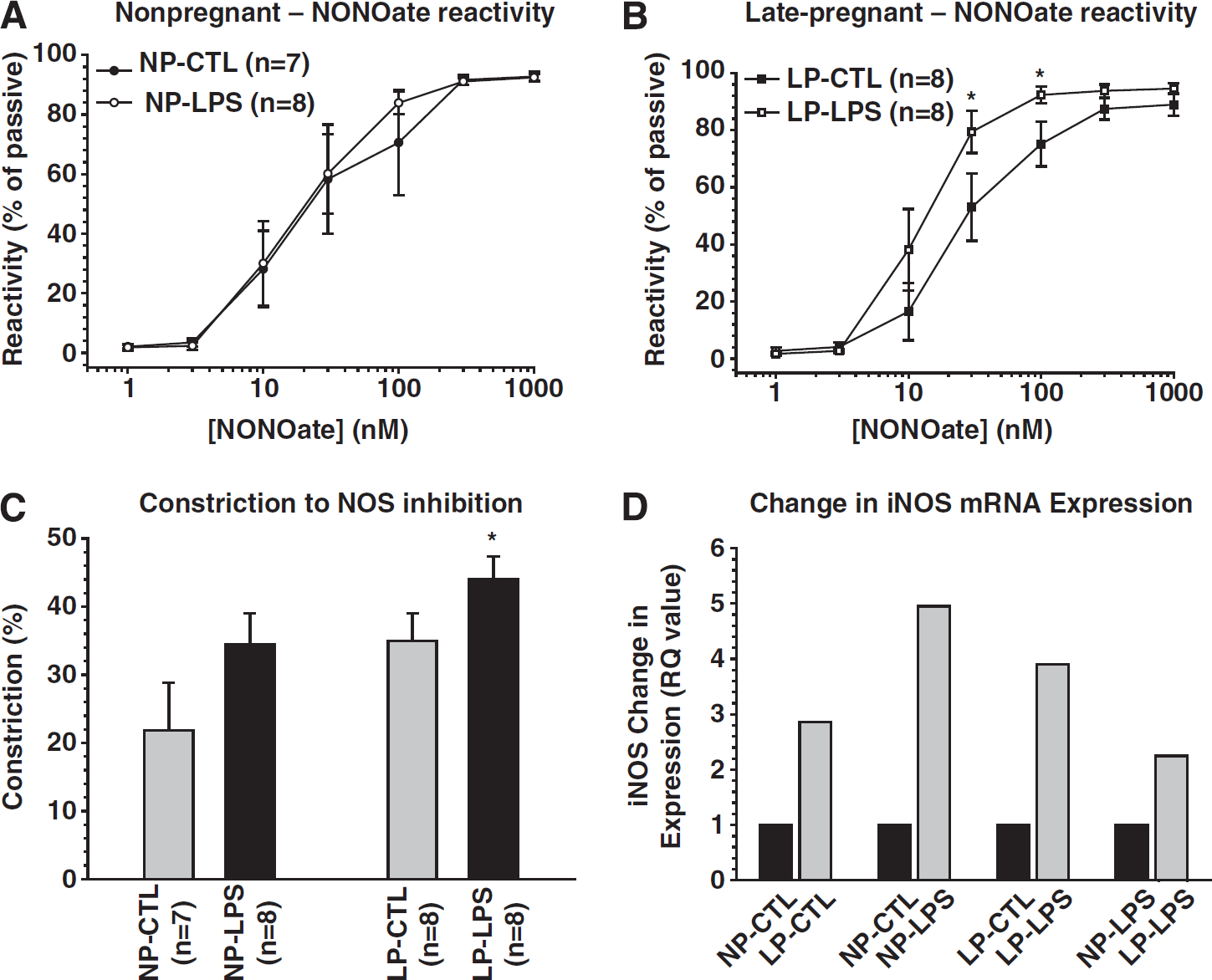
Effect of low-dose lipopolysaccharide (LPS) on nitric oxide (NO) vasodilation, contribution to tone and iNOS expression in cerebral arteries from pregnant and nonpregnant animals. (
Pregnancy and Cerebral Artery Remodeling During Chronic Hypertension
During chronic hypertension, the cerebral arteries undergo inward hypertrophic remodeling, that is, have smaller lumen diameters and thicker walls.44,45 Chronic hypertension also increases basal tone of cerebral arteries and together with inward remodeling, increase CVR. 46 The increase in CVR during chronic hypertension is considered protective of the downstream microcirculation from potentially damaging hydrostatic pressure that is increased during hypertension. 47 There is also a shift in the CBF autoregulatory curve to higher pressures during chronic hypertension that is protective of the microcirculation. 48 One interesting aspect of pregnancy is that it prevents hypertensive inward remodeling in female rats. 49 Female rats that were given the NOS inhibitor L-NAME in their drinking water for the last week of pregnancy had PCA that were similar in lumen diameter to controls whereas nonpregnant females treated with L-NAME for the same duration as pregnant animals had PCA that were significantly smaller in diameter with increased wall thickness. The lack of remodeling in the pregnant animals was not due to lower blood pressure as both nonpregnant and pregnant animals had a similar degree of hypertension with NOS inhibition. That pregnancy prevents hypertensive remodeling of cerebral arteries was confirmed in Dahl salt sensitive rats. 50 The mechanism by which pregnancy prevents inward hypertensive remodeling of cerebral arteries is not known, but may be related to the finding that pregnancy downregulates the angiotensin type 1 receptor (AT1R) in cerebral arteries (Figure 4A). 51
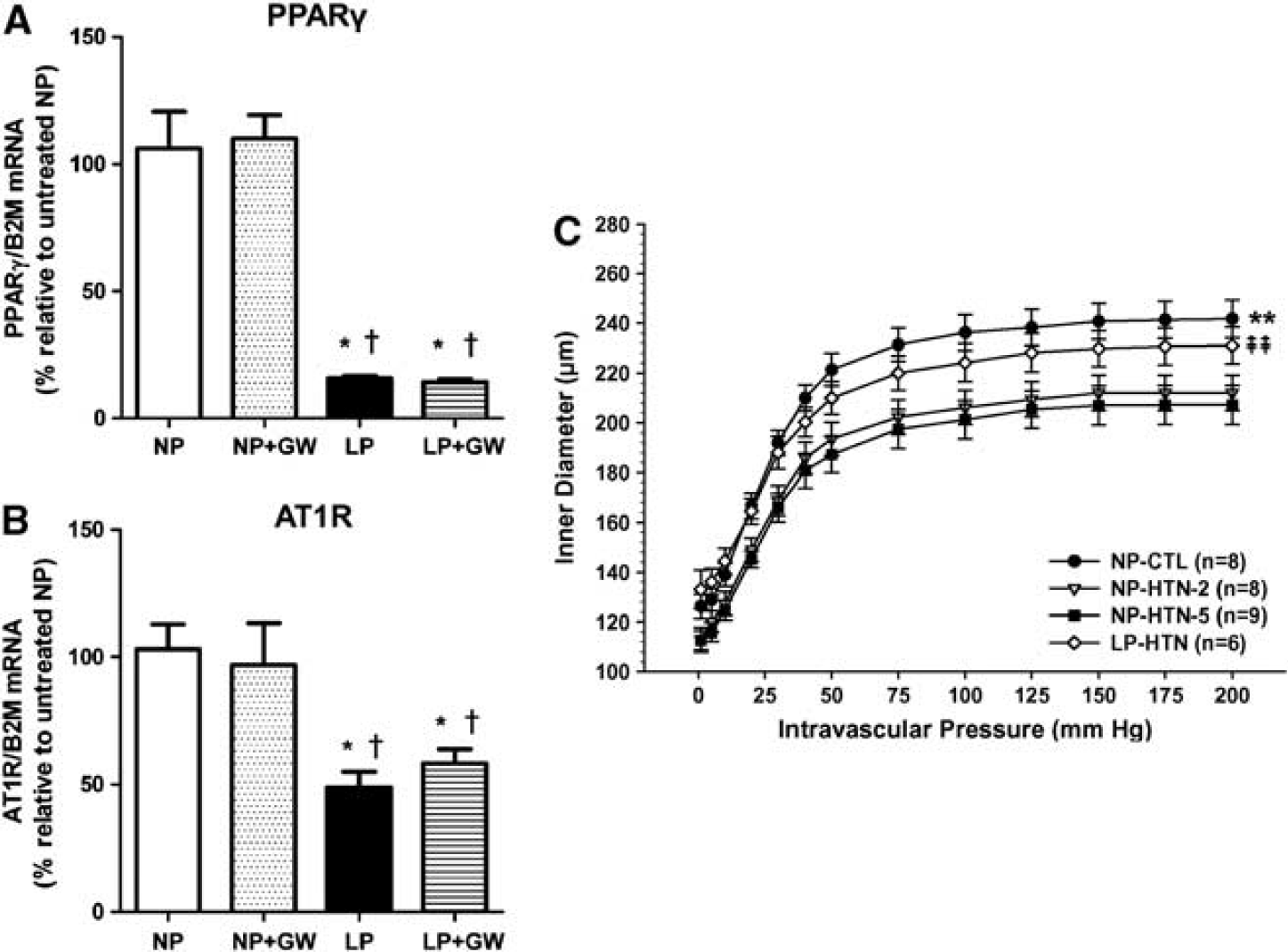
The effect of pregnancy on expression of peroxisome proliferator-activated receptor γ (PPARγ), angiotensin type 1 receptor (AT1R) and hypertensive remodeling of cerebral arteries. (
Probably even more interesting is that pregnancy can reverse preexisting hypertensive remodeling without lowering blood pressure. 52 Female rats that were hypertensive by NOS inhibition for 2 weeks were bred and PCA structure and biomechanical properties were measured 3 weeks later (late-gestation) and compared with nonpregnant rats that were hypertensive for 2 or 5 weeks. Nonpregnant rats that were hypertensive for 2 or 5 weeks had considerable inward hypertrophic remodeling that was similar, suggesting PCA from hypertensive animals before pregnancy had undergone remodeling. After 3 weeks of pregnancy, PCA had similar lumen diameters and wall thicknesses to normotensive controls (Figure 4C). Thus, one adaptation of the cerebral circulation to pregnancy is to limit the response to chronic hypertension by reversing and preventing inward hypertrophic remodeling. This may not be beneficial, however, given that blood pressure is still increased and therefore the protective effect of increased CVR that occurs during chronic hypertension is not present in the pregnant state.
The role of increased activation of peroxisome proliferator-activated receptor γ (PPARγ) during pregnancy as a mechanism underlying remodeling has also been explored. Peroxisome proliferator-activated receptor γ is a ligand-activated transcription factor expressed in numerous cell types and regulates genes involved in adipogenesis, glucose homeostasis, and lipid metabolism.53,54 Peroxisome proliferator-activated receptor γ is also expressed in vascular cells and has direct protective effects that are antihypertensive, antiinflammatory, and antiatherogenic.51,54–56 Peroxisome proliferator-activated receptor γ is highly activated during pregnancy and important for placental development and changes in maternal metabolism.57,58 Peroxisome proliferator-activated receptor γ activation suppresses expression of the AT1R in vascular smooth muscle and inhibits AT1R signaling involved in Ang II-mediated vascular remodeling.59,60 However, the role of PPARγ activation during pregnancy in cerebral artery remodeling is far from clear. Inhibition of PPARγ with the ligand inhibitor GW9662 in nonpregnant rats causes cerebral artery inward hypertrophic remodeling, similar to hypertension but without an increase in blood pressure. 51 Peroxisome proliferator-activated receptor γ inhibition in nonpregnant rats also impaired dilation to acetylcholine and sodium nitroprusside and increased basal tone, suggesting that activation of PPARγ in normotensive, nonpregnant rats is important for normal endothelial and smooth muscle function. Similar effects of PPARγ inhibition on the cerebral vasculature has been shown in several other studies.61–63 However, the effect of PPARγ inhibition during pregnancy has a very different effect than in the nonpregnant state. Peroxisome proliferator-activated receptor γ inhibition during pregnancy has little effect on cerebral arteries and does not affect endothelial or smooth muscle function, or inner diameter. 51 It does, however, cause a modest increase in wall thickness and outer diameter, suggesting some outward remodeling. The relative lack of effect of PPARγ inhibition during pregnancy may be related to the substantial decrease in PPARγ mRNA expression in cerebral arteries in response to pregnancy (Figure 4B). 51 Thus, pregnancy is associated with decreased PPARγ and AT1R expression in cerebral arteries that may underlie the propensity of these vessels to resist hypertensive remodeling (Figure 4C). The association between PPARγ and AT1R expression and signaling remains to be determined.
Changes in Reactivity and Structure of Parenchymal Arterioles and Capillaries During Normal Pregnancy
While the decreased level of PPARγ expression in cerebral arteries may explain the response of those vessels to PPARγ during pregnancy, this does not seem to be the case for brain parenchymal arterioles (PA). Parenchymal arterioles undergo a substantial increase in lumen diameter during pregnancy that is PPARγ-dependent; however, the expression of PPARγ mRNA is considerably decreased in this vessel segment compared with pial arteries. 20 Unlike pial arteries that change structurally and functionally very little during pregnancy, the PA undergo outward remodeling that increase lumen diameter (Figure 5). The increase in lumen diameter of PA during pregnancy is accompanied by little change in outer diameter but a significantly thinner vascular wall. Thus, pregnancy causes outward hypotrophic remodeling of PA (defined as an increase in lumen diameter at the expense of the vascular wall). One consequence of PA remodeling and vessel wall hypotrophy is that it substantially increases wall tension and wall stress in that segment of the vascular even without a change in intraluminal pressure. However, during acute hypertension when there is autoregulatory breakthrough and forced dilatation of upstream pial vessels, the increase in vascular volume and wall stress seriously compromises the integrity of the vessel wall and can promote edema formation (Figure 6). In fact, late-pregnant animals develop significant edema formation in response to acute hypertension, a response that is not seen in nonpregnant animals.32,33 While we cannot be certain that outward hypotrophic remodeling of PA is the underling or only mechanism by which this occurs, there is little direct effect of pregnancy on BBB permeability (see below), suggesting that hemodynamic changes are more likely to be the cause. In support of changes in hemodynamics as an underlying mechanism of edema formation in response to acute hypertension in pregnant animals, CVR is decreased to a greater extent in pregnant animals with autoregulatory breakthrough (Figure 1B). 20 The decrease in CVR during pregnancy is associated with an ~40% increase in CBF in pregnant animals during acute hypertension. Thus, it is possible that outward remodeling of PA during pregnancy decreases CVR during acute hypertension, increasing hydrostatic pressure and promoting edema formation.
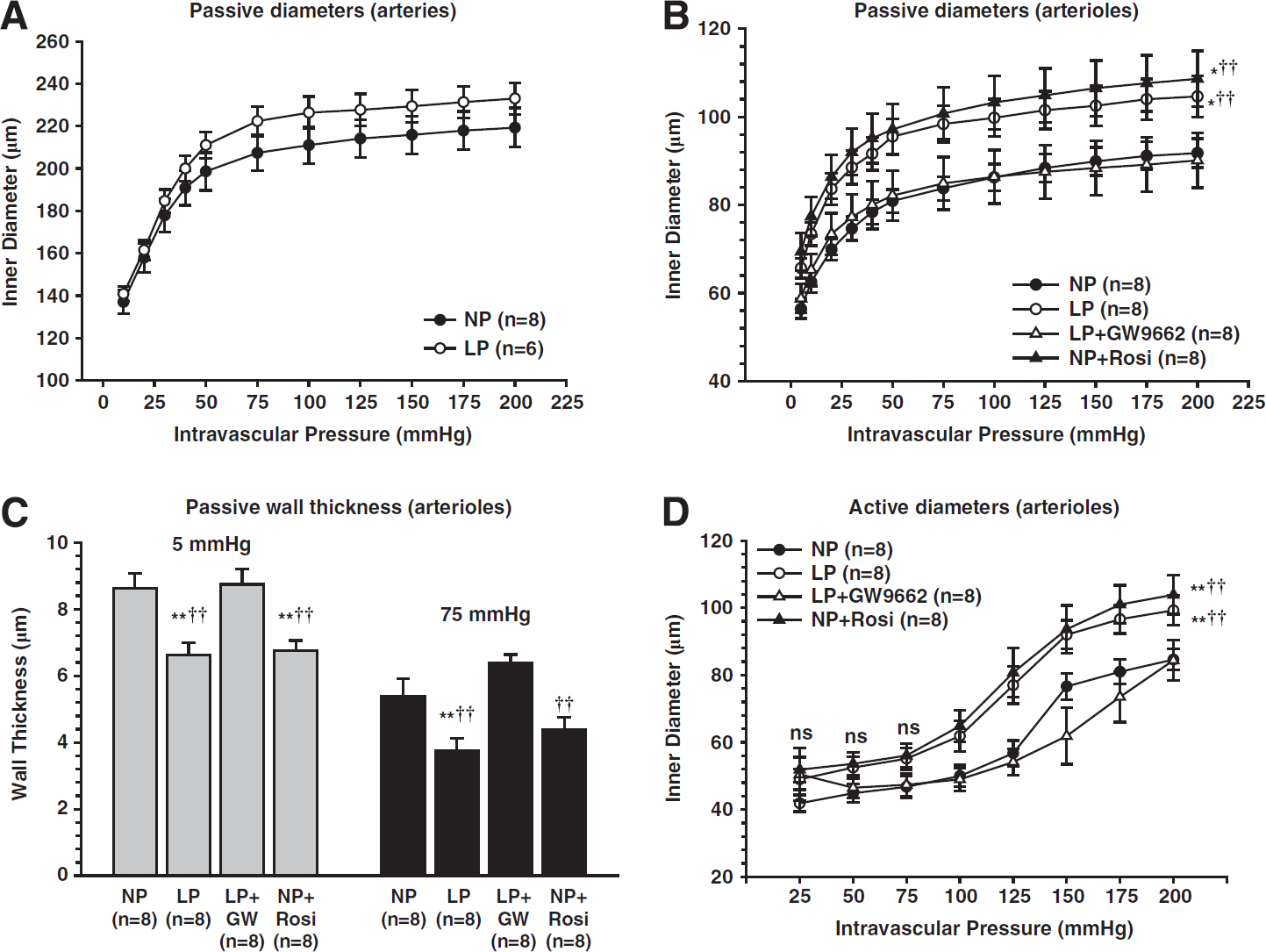
Effect of pregnancy and peroxisome proliferator-activated receptor γ (PPARγ) activation on remodeling of brain penetrating arterioles. Penetrating brain arterioles isolated from nonpregnant control (NP), late-pregnant control (LP), NP treated with the PPARγ agonist rosiglitazone for 3 weeks to mimic pregnancy (NP + Rosi), or LP treated with the PPARγ inhibitor GW9662 (LP + GW9662) for the last half of pregnancy were used to measure lumen diameter and wall thickness under pressurized conditions. (
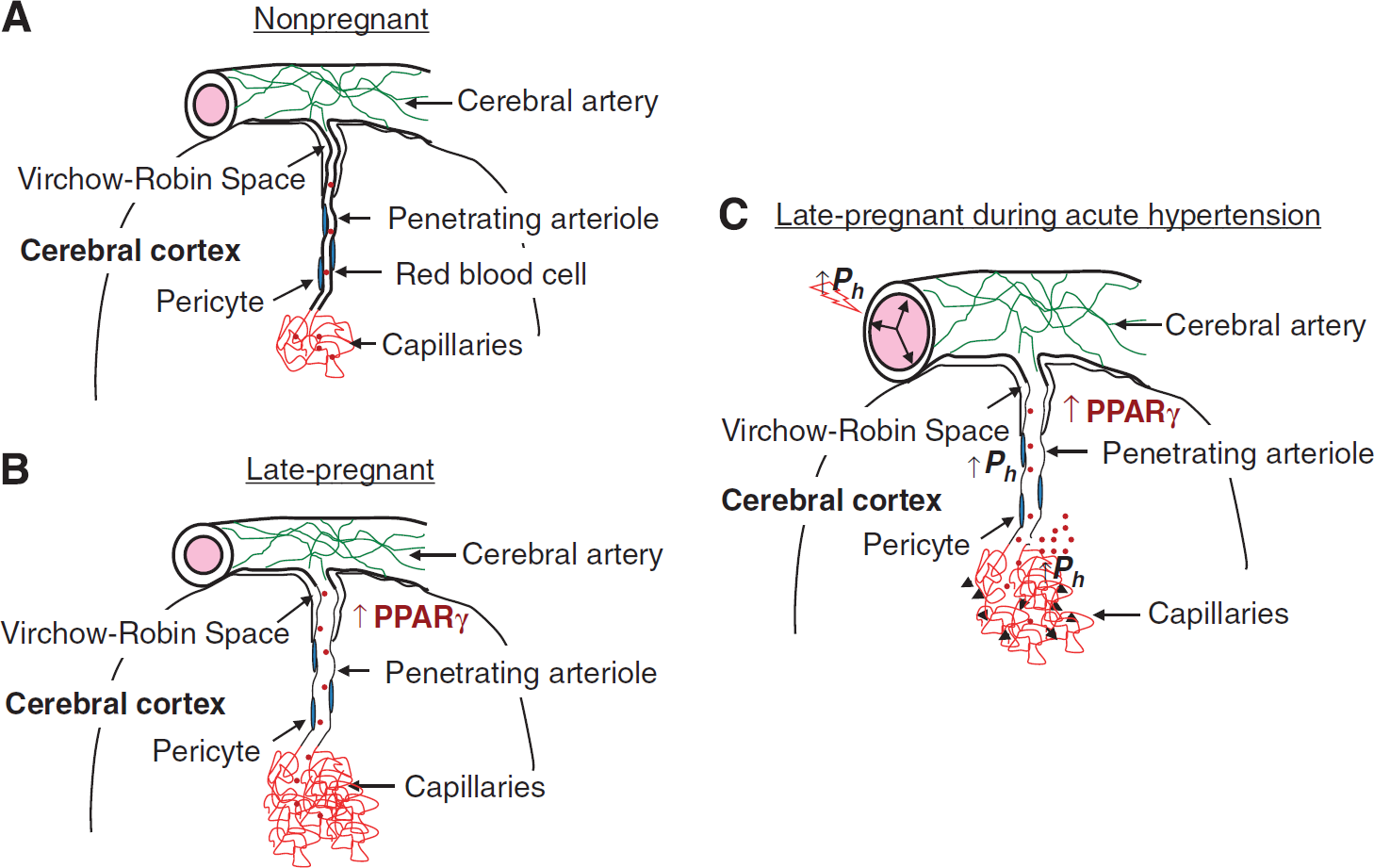
Summary diagram of cerebral vascular adaptation to pregnancy and the effect of acute hypertension. (
The outward remodeling of PA during pregnancy is prevented by PPARγ inhibition and mimicked by activation of PPARγ in nonpregnant animals, suggesting that increased activation of PPARγ during pregnancy is responsible for the change in these vessels (Figure 5). 20 Peroxisome proliferator-activated receptor γ has been shown to be more activated during pregnancy, although the endogenous activators have not been identified. 57 Recently, we investigated the role of the peptide hormone relaxin on PPARγ activation and outward remodeling of PA during pregnancy. This investigation was prompted because relaxin levels are high during pregnancy and has been shown to activate PPARγ in cells in culture.64,65 Relaxin treatment of nonpregnant rats to the level of mid-pregnancy caused selective outward remodeling of PA, similar to pregnancy, that was prevented by inhibition of PPARγ. 66 Thus, it appears that relaxin may be one of the endogenous activators of PPARγ that is elevated during pregnancy.
Capillary density is also affected by pregnancy in a region-specific manner. In the posterior cerebral cortex, but not anterior, pregnancy increases capillary density. 20 The increase in capillary number was mimicked in nonpregnant animals by PPARγ activation with rosiglitazone, but is not prevented by inhibition of PPARγ during the last 10 days of pregnancy. It is possible that the lack of effect of PPARγ inhibition on preventing the increase in capillary density during pregnancy is related to when during gestation capillaries increase, that is, the treatment with GW9662 may have started too late to affect angiogenesis. Similar to enlargement of PA lumen diameter, an increase in capillary density increases vascular volume and transvascular filtration that may contribute to edema formation that occurs during pregnancy in responses to acute hypertension.
Perivascular Innervation
Cerebral subarchnoid (pial) arteries are extrinsically innervated with sympathetic, parasympathetic and trigeminal nerve fibers. 67 Under physiological conditions, these perivascular nerves do not have a major impact on resting CBF.68,69 However, under pathological conditions they appear to have a more important role. For example, during acute hypertension, sympathetic nerve activity has a protective function that limits hyperperfusion and increased BBB permeability.70,71 Trigeminal nerve activation causes vasodilation in dural and pial arteries during migraine, and sense noxious stimuli.72–74 Thus, the interest in how pregnancy might influence perivascular nerve density is more related to pathologic conditions such as acute hypertension that occurs during preeclampsia/eclampsia, and severe and persistent headache that is common in preeclampsia. 75
Pregnancy has been shown to significantly increase the density of calcitonin gene-related peptide (CGRP)-containing nerves on PCA, suggesting an effect of pregnancy on trigeminal innervation. 76 This effect was not found with tyrosine-hydroxylasecontaining nerves, suggesting no effect of pregnancy on sympathetic innervation of pial arteries (Figure 7). The consequence of an increase in CGRP-containing nerves during pregnancy is not clear, but may relate to the appearance of headache during preeclampsia/eclampsia. In fact, headache is the most common neurological symptom of these conditions. 75 In addition, women with migraine often have their symptoms disappear during pregnancy, which again may be related to change in trigeminal innervation during pregnancy. 77 It is worth noting that only PCA were studied and not dural vessels, one of the most important vessels in the pathogenesis of migraine. Interestingly, there were little CGRP-containing nerves in PCA from male rats.
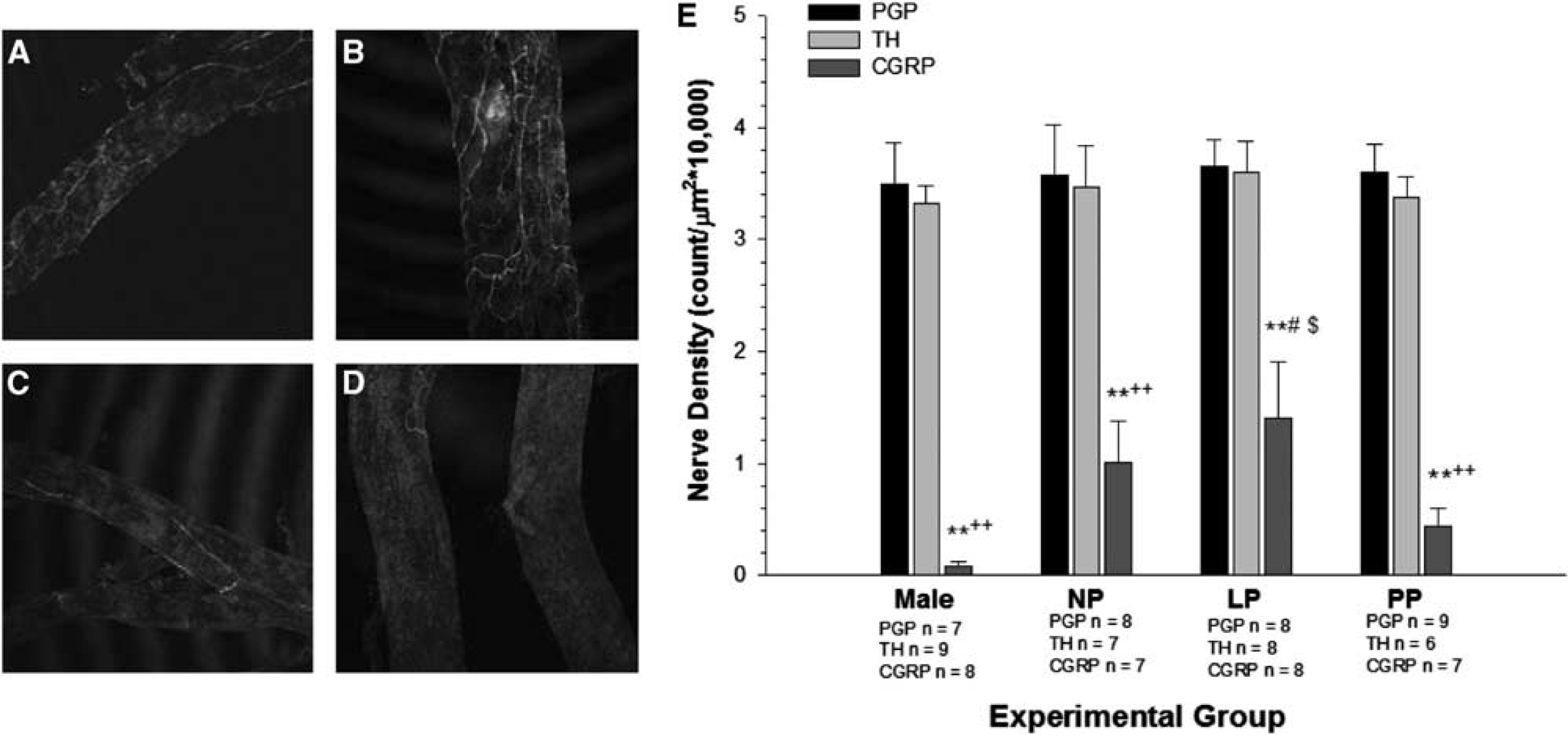
The effect of pregnancy on perivascular innervation of cerebral arteries. Photomicrographs (10 ×) of posterior cerebral arteries stained for calcitonin gene-related peptide (CGRP)-containing nerve fibers from (
The Cerebral Veins
In addition to cerebral arteries, cerebral veins have an important role in control of hemodynamics under normal physiological conditions and have been implicated in the development of several pathological states. Unlike the arterial side of the vasculature, 70 to 80% of cerebral blood volume resides on the venous side. 78 Thus, changes in venous outflow can significantly affect cerebral blood volume and intracranial pressure.79–81 In addition, thrombosis of cerebral veins and/or venous sinuses can cause serious neurological complications such as venous infarction and intracranial hemorrhage. 82 Pregnancy increases the risk of several complications associated with cerebral veins, including intracranial venous thrombosis, stroke, and cerebral hemorrhage, partly due to pregnancy being a hypercoaguable state.82–84 However, how pregnancy changes cerebral veins in addition to its effect on the coagulation cascade is largely unknown.
A recent study investigated how pregnancy affects the cerebral vein of Galen, a vein from the deep venous system. 85 This large vein provides drainage for the medial areas of the diencephalon, basal ganglia, midbrain, medial aspect of the cerebral hemisphere, and the corpus callosum. 86 Thus, it serves a great territory of the brain. This vein is surrounded by a layer of large smooth muscle cells and has a low level of basal, pressure-induced tone (4 to 6%) in the nonpregnant state that decreases during pregnancy to only 1 to 2% (Figure 8). The decrease in tone in the vein of Galen during pregnancy causes these vessels to have larger lumen diameters. However, the increase in lumen diameter is not only due to decreased tone during pregnancy. Under passive conditions, the vein of Galen is also larger with a significantly thinner vascular wall that substantially increases wall stress. Thus, the vein of Galen undergoes outward hypotrophic remodeling during pregnancy, similar to PA. Whether or not pregnancy-induced remodeling of the vein of Galen is due to PPARγ activation or relaxin is not known. However, the enlargement of cerebral veins during pregnancy may promote venous pooling or stasis, an effect that would be exacerbated by increased coagulation. In addition, the increase wall stress in the veins during pregnancy could also promote rupture or dissection, especially under hypertensive conditions. It is important to note that it is not known what effect there is on other cerebral veins during pregnancy such as the parenchymal veins and venules that are the effectors of petechial hemorrhage that occurs during eclampsia. 10
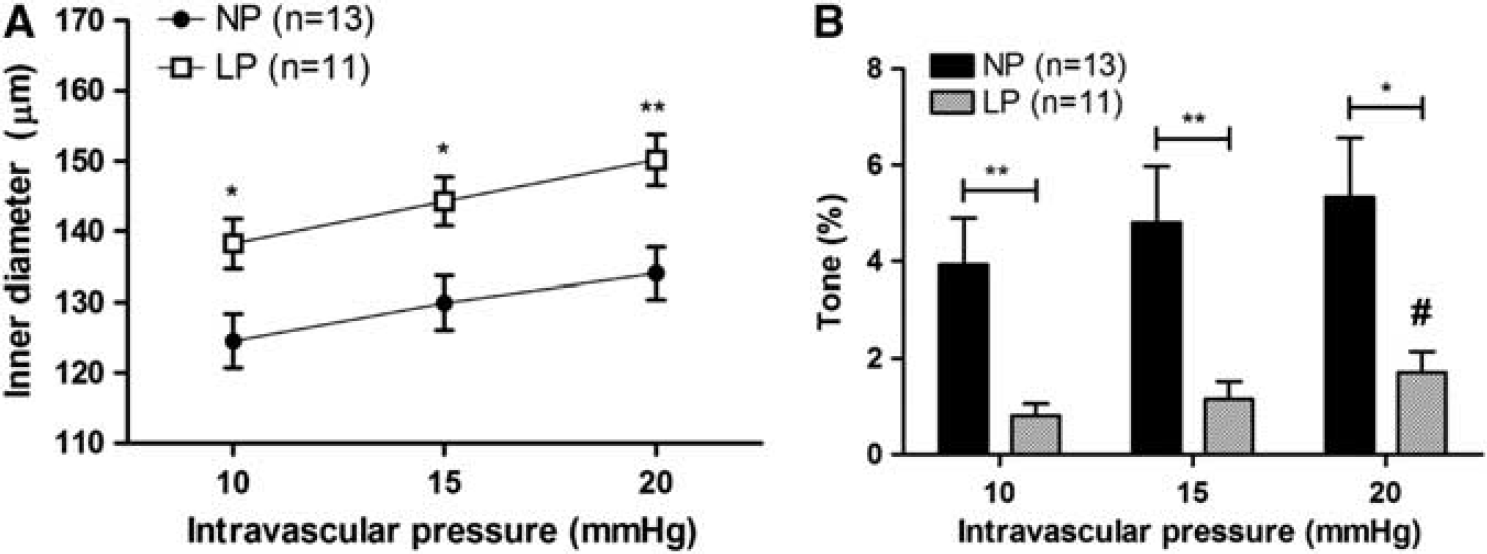
The effect of pregnancy on active diameters and tone of cerebral veins. (
THE ADAPTATION OF THE BLOOD–BRAIN BARRIER TO PREGNANCY AND THE EFFECT OF CIRCULATING FACTORS
The cerebral endothelium that comprises the BBB is structurally and functionally distinct from endothelium outside the CNS in that it contains high electrical resistance tight junctions that restrict not just paracellular flux of proteins, but also ions. 87 This modifies Starling's forces such that the ion concentration of plasma opposes hydrostatic pressure to control transvascular filtration because ions are orders of magnitude greater in number than proteins. 88 The BBB also does not contain fenestrations and has low transcellular transport compared with endothelium in the periphery. 87 Lastly, passage of many proteins, including cytokines and growth factors, are regulated by numerous transporters and receptors that are either influx or efflux. 87 Together, these properties provide a highly regulated interface between the blood and the brain that is protective against vasogenic edema and passage of damaging serum constituents into the brain parenchyma.
The adaptation of the BBB during pregnancy is of considerable interest for several reasons. First, during normal pregnancy, the endometrium, decidua and placenta produce large amounts of cytokines (e.g., TNFα) and angiogenic growth factors (e.g., vascular endothelial growth factor, VEGF) that are released into the circulation.89–91 These hormones are essential for normal intrauterine development of the fetal-placental unit and drive the cardiovascular adaptation to pregnancy. However, several of these factors are vasoactive and have the potential to increase vascular permeability, including BBB permeability to water and solutes. 90 Remarkably, despite high levels of circulating permeability factors, vasogenic brain edema does not normally develop over the course of gestation, suggesting that there may be adaption of the BBB to pregnancy that limits the impact of these factors. Second, hypertension can have a significant effect on BBB permeability and is common during gestation. Pathologic conditions such as early-onset preeclampsia and HELLP syndrome produce severe disease that are associated with neurologic symptoms, with BBB disruption as a central etiology.10,26 However, women who develop preeclampsia and HELLP syndrome are by definition normotensive and asymptomatic before pregnancy, suggesting that pregnancy alone may impact the BBB in these conditions. Lastly, BBB disruption also has a central role in the appearance of seizure during pregnancy.10,26,27 In fact, the primary explanation for the neurologic symptoms associated with eclampsia, most notably seizure, is that it arises from an acute hypertensive episode that causes autoregulatory breakthrough, decreased CVR and hydrostatic brain edema.10,26,27 Thus, how pregnancy alters the permeability of the BBB under normal conditions, as well as in response to acute hypertension, is important to understand if conditions such as eclampsia are to be avoided.
Effect of Pregnancy on Blood–Brain Barrier Permeability to Solutes and Water
In
The effect of pregnancy on the blood–brain barrier (BBB).
The Role of Circulation Factors in the Adaptation of the Blood–Brain Barrier to Vascular Endothelial Growth Factor During Pregnancy
One of the most important growth factors for successful pregnancy is VEGF. 91 It is produced in a variety of cells at higher levels during pregnancy including endothelial cells.96,97 VEGF was initially discovered as a permeability factor but is now recognized as having a role in angiogenesis, vascular growth, endothelial cell survival, and vasodilation through complex interaction of VEGF with its two receptors, FMS-like tyrosine kinase 1 (Flt1) or VEGF receptor 1 (VEGFR1) and fetal liver kinase 1 (Flk1) or VEGF receptor 2 (VEGFR2).98–100 In the uterine circulation, VEGF expression and its receptor VEGFR1 are increased during pregnancy that promotes growth and increases permeability of that vascular bed. 101 In cerebral veins, VEGF mRNA expression is also increased threefold over the nonpregnant state, without an increase in receptors VEGFR1, VEGFR2, or neuropilin (Figures 10A and 10B). 94 Similar to the uterine circulation, VEGF increases BBB permeability of cerebral veins; however, circulating factors during pregnancy control its actions. When cerebral veins are perfused with plasma from nonpregnant animals, VEGF induces a significant increase in BBB permeability. However, when a cerebral vein from a late-pregnant rat is perfused with late-pregnant plasma and similarly treated with VEGF, there is no increase in BBB permeability (Figure 10C). The lack of VEGF-induced permeability occurred despite a threefold increase in VEGF expression in those veins. That circulating factors are responsible for preventing VEGF-induced permeability was shown by perfusing a vein from nonpregnant animals—that does have increased permeability in response to VEGF—with plasma from late-pregnant animals and showing that it prevents the increase in permeability. 94 Although numerous factors are increased in late-pregnant plasma, soluble Flt1 (sFlt1) is the most likely candidate. In fact, addition of sFlt1 to nonpregnant plasma prevents VEGF-induced permeability similar to late-pregnant plasma (Figure 10D). 94 SFlt1 is significantly increased in the maternal circulation during pregnancy and controls the biologic actions of VEGF. 102 In the cerebral circulation, the importance of sFlt1 in controlling VEGF-induced permeability is paramount and one of the most important adaptations that occurs during pregnancy because it prevents cerebral edema in the face of high circulating permeability factors.
The role of circulating factors in the adaptation of the blood–brain barrier (BBB) to vascular endothelial growth factor (VEGF) during pregnancy. (
Excessively elevated sFlt1 levels during pregnancy has been implicated in the pathogenesis of preeclampsia.
103
Large amounts of sFlt1 can alter the angiogenic balance by removing too much VEGF from the maternal circulation, causing endothelial dysfunction. However, in the cerebral circulation, high levels of sFlt1 did not promote endothelial dysfunction or affect permeability. In fact, elevated sFlt1 to the level of preeclampsia has a similar protective effect on VEGF-induced permeability as levels produced during normal pregnancy (Figure 10D).
94
It is possible that longer periods of exposure to excessive sFlt1
The Adaptation to Seizure Provoking Serum During Pregnancy In addition to increased circulating angiogenic factors, normal pregnancy is associated with large amounts of hormones and cytokines that produce a state of mild peripheral inflammation.104–106 The production of pro-inflammatory cytokines is important for normal fetal-placental development during pregnancy. However, peripheral inflammation can affect the brain and promote seizure by passage of activated leukocytes through the BBB, activation of microglia, and production of TNFα that causes neuronal hyperexcitability. 107 The hyperexcitable nature of pregnant plasma was demonstrated using a hippocampal slice culture model in which the normal horse serum that the slices are normally grown in was replaced by serum from nonpregnant or late-pregnant rats. The late-pregnant serum caused hyperexcitability of neurons when evoked potentials were measured, an effect that did not occur with nonpregnant serum. 95 Neuronal hyperexcitability in response to exposure to late-pregnant serum was associated with microglial activation and prevented by addition of soluble TNF receptor 1. However, TNFα was not increased in the late-pregnant serum, suggesting another circulating factor in the serum activated the microglia and production of TNFα to promote hyperexcitability. What that factor might be is not known.
One important aspect of this study is that the pregnant animals from which the serum was taken were not having seizure, pointing to an important role for the BBB in protecting the brain from seizure provoking serum during pregnancy. It is possible that the BBB adapts to the high levels of circulation factors over the course of gestation to prevent passage into the brain and protect it from hyperexcitability. It is not known if certain receptors or transporters adapt to pregnancy to control or limit exposure of these factors to the brain. Alternatively, the BBB may be sufficiently equipped to handle the rise in these circulating factors during gestation. Regardless, the importance of this finding may relate to the ~20% of women who have unexplained seizure during seemingly normal pregnancy, that is, without the diagnosis of preeclampsia.38,108,109 It is possible that in these cases either the adaptation to these factors is not sufficient or the BBB transporters/receptors are overcome by the high levels of seizure provoking serum.
Effect of Pregnancy on AQP4 in the Brain
The aquaporins (AQPs) are a family of channel-forming transmembrane proteins that facilitate the movement of water, glycerol, and other solutes across the plasma membrane of cells.110–113 AQP4 is the predominant AQP in the brain and is localized in the astrocytic endfeet surrounding blood vessels, but not cerebral endothelium.114–116 The involvement of AQP4 in brain edema has been demonstrated using knockout mice for both AQP4 and α-syntrophin, a membrane protein anchoring AQP4 to astrocytic endfeed. In α-syntrophin knockout mice, there was decreased edema after focal ischemia, suggesting that the presence of AQP4 in astrocytes enhances edema. 117 In addition, AQP4 knockout mice were found to have reduced cytotoxic edema after ischemic stroke and acute hyponatremia. 118 Thus, AQP4 regulates brain water content under pathologic conditions.
Because of its role in promoting brain edema, how pregnancy affects AQP4 is of interest to conditions such as early-onset preeclampsia and eclampsia in which edema formation is the major pathologic event that causes brain injury. 10 AQP4 protein and mRNA are significantly increased in the brain during pregnancy with a peak mid-gestation compared with late-pregnancy or postpartum.92,119,120 The consequence of increased AQP4 in the brain during pregnancy is not clear; however, pregnancy causes increased edema formation in response to acute hypertension that is not due to increased BBB permeability.20,32,33 Whether increased AQP4 during pregnancy is involved in edema formation in response to acute hypertension is not known. One important aspect of AQP4 in the brain is that it affects seizure threshold. AQP4 knockout mice have a higher seizure threshold, suggesting that conditions that increase AQP4 in the brain, such as pregnancy, would have a lower seizure threshold. 121 The effect of pregnancy on seizure threshold has not been measured but is a potentially important adaptation to pregnancy that may predispose to seizure, especially during conditions that increase BBB permeability such as preeclampsia and eclampsia.
PERSPECTIVES
Pregnancy poses a unique challenge to the brain and brain circulation that is distinct from other organs. While other organ systems undergo substantial increases in blood flow, the adaptation of the cerebral circulation appears to be to maintain relative constancy in the face of large increases in plasma volume and CO. However, the cerebral circulation is not unchanged during pregnancy. Similar to other vascular beds, the cerebral circulation is more sensitive to the effects of LPS and has increased expression of iNOS and pro-inflammatory cytokines during normal pregnancy. Although the large subarchnoid pial arteries do not undergo substantial changes in structure or function during normal pregnancy, the PA undergo outward hypotrophic remodeling. In addition, pregnancy promotes an increase in capillary density in the posterior cerebral cortex. The increase in PA lumen diameter in conjunction with a thinner vascular wall substantially elevates wall tension that could predispose to edema or hemorrhage during excessive elevations in blood pressure, as seen with severe preeclampsia and eclampsia. In addition, the increase in capillary density could also serve to increase transvascular filtration in response to increased hydrostatic pressure and may underlie the increase in edema formation that occurs in pregnant animals during acute hypertension. The enlargement of PA and greater capillary density in the brain may also have a beneficial effect by contributing to the shift in the lower limit of autoregulation of CBF to lower pressures. The shift in the autoregulatory curve to lower pressures may be protective of hypoxic/ischemic injury to the brain during hemorrhagic hypotension that occurs during parturition. Pregnancy also downregulates AT1R and PPARγ in the cerebral circulation that are likely responsible for both preventing and reversing hypertensive inward remodeling. Lastly, the BBB adapts to high levels of circulating angiogenic and permeability factors that rise over the course of gestation to prevent edema formation. Although VEGF expression is increased threefold in cerebral veins during pregnancy, an increase in VEGF-induced BBB permeability is prevented by high levels of circulating anti-angiogenic factors such as sFlt-1. The adaptation of the BBB to circulating factors during pregnancy is probably one of the most important adaptations that occur to prevent increased permeability and limit passage of seizure provoking serum into the brain.
DISCLOSURE/CONFLICT OF INTEREST
The authors declare no conflict of interest.