
Abstract
Sex differences in stroke are observed across epidemiologic studies, pathophysiology, treatments, and outcomes. These sex differences have profound implications for effective prevention and treatment and are the focus of this review. Epidemiologic studies reveal a clear age-by-sex interaction in stroke prevalence, incidence, and mortality. While premenopausal women experience fewer strokes than men of comparable age, stroke rates increase among postmenopausal women compared with age-matched men. This postmenopausal phenomenon, in combination with living longer, are reasons for women being older at stroke onset and suffering more severe strokes. Thus, a primary focus of stroke prevention has been based on sex steroid hormone-dependent mechanisms. Sex hormones affect different (patho)physiologic functions of the cerebral circulation. Clarifying the impact of sex hormones on cerebral vasculature using suitable animal models is essential to elucidate male–female differences in stroke pathophysiology and development of sex-specific treatments. Much remains to be learned about sex differences in stroke as anatomic and genetic factors may also contribute, revealing its multifactorial nature. In addition, the aftermath of stroke appears to be more adverse in women than in men, again based on older age at stroke onset, longer prehospital delays, and potentially, differences in treatment.
INTRODUCTION
Cerebrovascular disease is the second leading cause of death worldwide, accounting for 10% of all deaths. 1 The most common form of cerebrovascular disease is stroke, and the focus of this review. Cerebrovascular disease is generally characterized by increased endothelial damage of the brain blood vessels, leading to inflammation and accumulation of fatty deposits (e.g., low-density lipoproteins (LDLs) and cholesterol) in the damaged arterial wall, thus inducing atherosclerosis. 2 Atherosclerosis reduces the diameter of blood vessels, disturbing blood flow and transport of oxygen and glucose, which leads to deterioration of brain tissue. The term ‘stroke’ is applied when an artery within the brain is blocked by a thrombus causing an ischemic stroke; or the artery bursts, causing a hemorrhagic stroke. 3 Ischemic stroke accounts for 87% of all strokes, while 10% are intracerebral hemorrhage and 3% are subarachnoid hemorrhage strokes. 4 Stroke alone is the third leading cause of death behind diseases of the heart and cancer. More than five million people die of stroke every year, or one person every minute. 5 Other forms of cerebrovascular disease include transient ischemic attack, cerebral small vessel disease, and vascular dementias.
Given the contribution of stroke to the global burden of disease, stroke prevention is of major interest and has high impact. Healthcare costs related to stroke occurrence continue to escalate. Not only are excess costs accrued as a result of the primary stroke, but also as a result of morbidities accompanying stroke and recurring strokes among stroke survivors. 6 Stroke prevention is attainable, since the epidemiologic literature suggests a plethora of modifiable risk factors for stroke. However, published findings also indicate innate differences in stroke occurrence between women and men, and it is not clear why these sex differences exist. Interactions of biologic sex with physiologic, genetic, lifestyle, and other factors may provide clues as to more effective stroke prevention strategies. The aim of this review is to overview sex differences in stroke epidemiology, in underlying pathophysiologic mechanisms identified in humans and rodents, as well as treatments and outcomes.
THE EPIDEMIOLOGY OF STROKE
Epidemiologic studies show differences between women and men in stroke incidence, prevalence, and mortality. In addition, there are differences by age and age group over the life course.4,7,8 Herein, we review the epidemiologic data on sex differences in stroke, highlighting the roles of age and aging, as well as risk factors, stroke subtypes, and stroke severity.
Incidence, Prevalence, and Mortality
Every 40 seconds, a stroke occurs in the United States 4 and each year 15 million strokes are incident worldwide. 9 Despite this, population data for high-income countries revealed a 42% decrease overall in worldwide stroke incidence rates from 163 per 100,000 person-years in 1970 to 1979 to 94 per 100,000 person-years in 2000 to 2008. 10 This decline was especially observed for men. Sex-specific data from the Framingham Heart Study showed decreases of 30.3% for men and 17.8% for women from 1950 to 2004. 11 Overall, the global age-adjusted male/female (m/f) incidence ratio was 1.33 for that period, being highest among those aged 35 to 44 years, and declining after 75 years. 7 Despite an overall decrease in incidence, some studies indicate a recent surge in mid-life stroke incidence among women potentially because of rising obesity and related metabolic syndromes. 12
Similarly, prevalence of stroke increases with age, particularly among women. According to the American Heart Association, worldwide stroke prevalence was 3% in 2008, 4 being most prevalent in men (age-adjusted m/f ratio of 1.41). However, at 85 years of age and older, stroke was more common in women compared with men. 7 Part of this is explained by a lower global life expectancy for men versus women from 85 years: 5.9 versus 6.8 years, respectively. 13 Nevertheless, prevalent stroke increases exponentially in both sexes with age. 14
In terms of mortality, the WHO (World Health Organization) assessed sex-specific mortality rates across 39 countries in Europe and Central Asia,
9
and reported an excess of total deaths because of stroke among women (
Excess stroke in women at high age may arise from longer life expectancy and reaching ages of highest stroke risk compared with men.9,14 Similarly to other cardiovascular risk factors, women experience stroke later than men potentially because of loss of protection by estrogen after menopause (see section ‘Sex-specific pathophysiologic mechanisms’). In addition, women evidence poorer prestroke functionality, multimorbidities, and less social support7,8 at age 75 years and older, which may contribute to observed sex and age differences.
Risk Factors
Traditional risk factors for stroke include hypertension (HTN), Type 2 diabetes mellitus (T2DM), hyperlipidemia, atrial fibrillation, cigarette smoking, overweight, and obesity, other metabolic syndromes, and coronary artery disease. Other suggested risk factors include hyperhomocysteinemia and hypercoagulable states (see Table 1). 15 Modification of these cardiovascular risk factors reduces the risk of stroke, 16 and may be responsible for the decreases in stroke incidence over the past 40 years. Risk factor modification may be achieved pharmacologically through the use of, for example, antihypertensive agents (e.g., thiazide-like diuretics), lipid-lowering medications (e.g., statins), antiplatelet therapies (e.g., aspirin), and anticoagulants (e.g., heparin). 17 Lifestyle modification, as recommended by the American Stroke Association, such as daily exercise, reducing dietary intake of sodium and fat, and weight loss, may also be beneficial. 18 The gradual accrual of modifiable risk factors over time contributes to the higher occurrence of stroke with aging. 19 Innate risk factors for stroke include high age, male sex, non White ethnicity/race, and family history of stroke (see Table 1). 15 However, stroke ultimately affects more women 8 resulting in poorer outcomes 20 as mentioned earlier and explained later on.
Modifiable and nonmodifiable risk factors for stroke 15
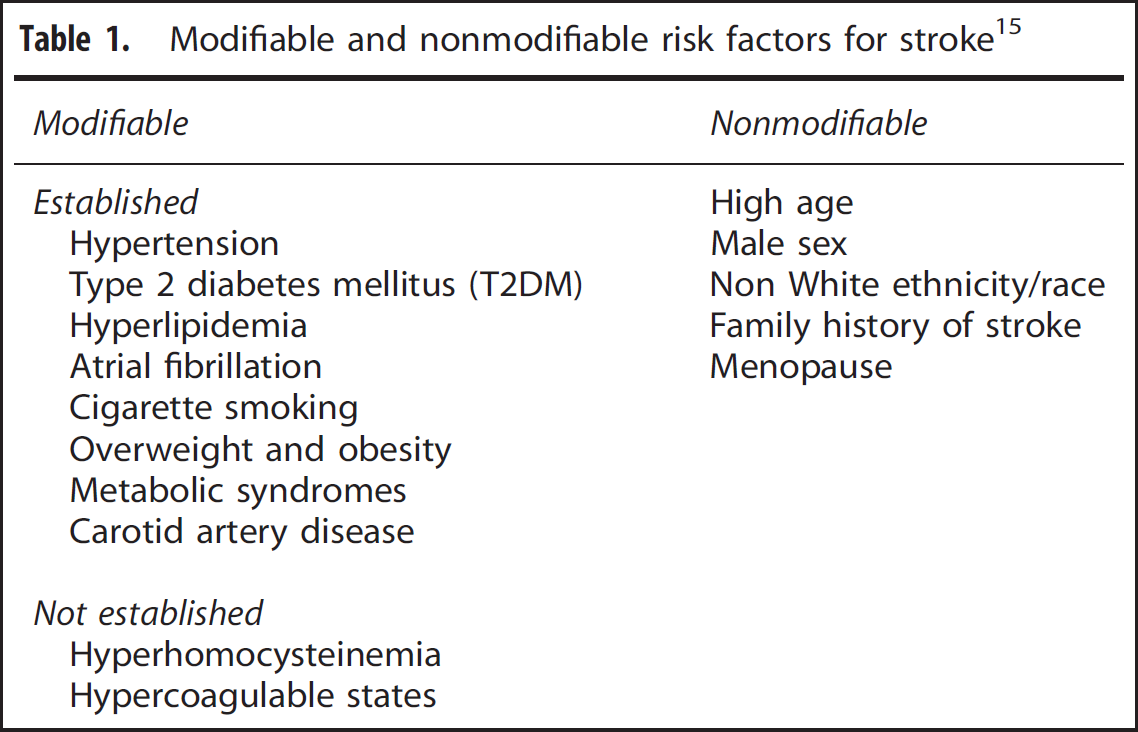
The presence, severity, and timing of risk factors (Table 1) are important determinants of stroke. 16 As aforementioned, accumulation of stroke risk factors with aging may be more likely in women since women are older at stroke onset than men (±4 years)4,7 and live longer. 13 Not only is stroke experienced later by women, but also other cardiovascular risk conditions, such as HTN, central obesity, and hyperlipidemias, are also present at a later age in women than in men. However, it is also important to note that women and men surviving to high age may also comprise a surviving ‘elite’ and be less affected by vascular risk. 21 Earlier and varied pharmacological interventions for the multimorbidities of aging may also contribute to observed differences (see section ‘Differences in stroke treatment and outcome’). There are also sex differences in the stroke risk factor profiles between women and men. For example, atrial fibrillation and HTN (blood pressure (BP) > 140/90 mm Hg) occur more often in female compared with male stroke patients (see Table 2), 22 despite that BP is consistently higher in younger men than in premenopausal women. 23
Examples of reported risk factor prevalences among women and men who experience a stroke
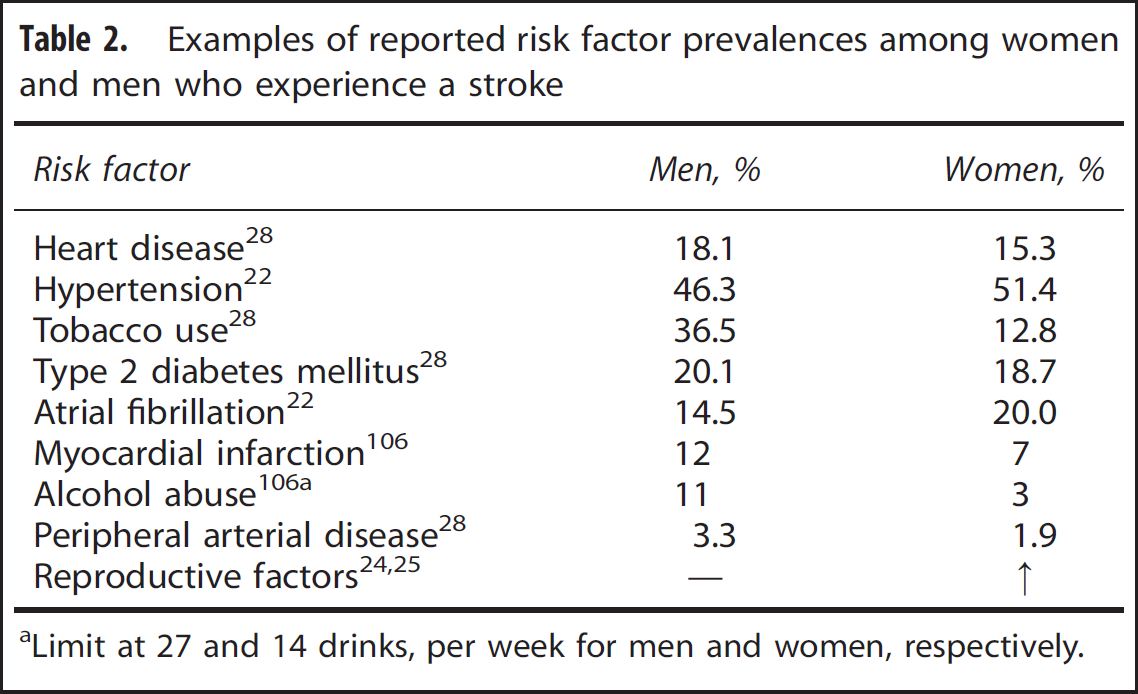
Limit at 27 and 14 drinks, per week for men and women, respectively.
Sex hormone-related factors are also associated with stroke risk in both women and men. In women, use of oral contraceptives, more pregnancies, and the postpartum and peri- and postmenopausal periods increase stroke risk.24,25 Menopause is associated with body weight gain, metabolic syndromes, and higher BPs, all risk factors for stroke. 26 Early menopause, which is linked to cigarette smoking, poor nutritional quality and low socioeconomic status, adversely influences blood vessel integrity.26,27 In contrast, heart disease, myocardial infarction, peripheral arterial disease, T2DM, and alcohol and tobacco use tend to precede stroke in men, 28 as do low levels of circulating testosterone in elderly men. 29 Low testosterone commonly occurs with aging, T2DM, myocardial infarction, and other stroke risk factors. 30 Thus, while low estrogen levels may be commonly associated with stroke in women, low testosterone levels may produce a comparable milieu for men.
Not only do risk factors and their distributions vary between women and men, but also risk effects, as estimated by relative risk (RR). For example, even though T2DM is more prevalent among men with stroke, it may have a stronger effect on stroke risk in women (RR: 2.1 to 6.6 for women versus 1.3 to 2.3 for men). 31 This is similarly observed for metabolic syndrome, which has a greater effect in women (RR: 1.3 to 3.1) versus men (RR: 0.6 to 1.9); 32 and is suggested for smoking (women: RR: 1.78 to 2.33 versus men: 1.66 to 2.00). In contrast, the effect of aging is similar for both women (RR: 1.09 to 1.10) and men (RR: 1.09 to 1.10). 33
Stroke Subtype and Severity
There are numerous stroke classification schemes. These include those originating from the US NIH/NINDS (National Institutes of Health/National Institute of Neurological Disorders and Stroke); OCSP (Oxfordshire Community Stroke Project); TOAST (Trial of ORG 10172 in Acute Stroke Treatment); and Étude du profil Génétique de l'Infarctus Cérébral (GÉNIC). Despite numerous classification schemes, all discriminate between the two primary causes of brain injury related to stroke—ischemia (87% of total stroke number) and hemorrhage (13%).3,4 Ischemic, intracerebral, and subarachnoid hemorrhages account for 87, 10, and 3% of total strokes, respectively. 4 Ischemic strokes are either thrombotic (50% of all strokes) or embolic. 34 Thrombosis is triggered by the Virchow's triad, i.e., hypercoagulability, hemodynamic changes (e.g., stasis), and endothelial injury. 35 The most common pathophysiologic process of this triad is atherosclerosis, or thickening of the vessel wall, which amplifies over years. Eventually, this may lead to ‘arterial wall breakdown’ and release of pro-coagulant and pro-thrombotic factors in the blood. 36 Other processes affecting blood clot formation include fibromuscular dysplasia, arthritis, and dissection of the vessel wall. In some cases, blood clots may detach and travel along the vascular pathway causing embolic stroke distant from the atherosclerotic site. Hemorrhagic strokes are characterized by ruptured vessels that cause leakage of blood into intracerebral areas (10% of all strokes) or the subarachnoid space (3%), primarily as a result of hypertensive-related degenerative changes or cerebral amyloid angiopathy. 37 Ischemic stroke and intracerebral hemorrhage are observed more often in men (m/f: 1.55 and 1.60, respectively); while subarachnoid hemorrhage is more common in women (m/f: 0.84), 7 with little variation by age. 7
According to the NIHSS (National Institute of Health Stroke Scale), significantly more women (44%) than men (36%) appear to experience severe stroke (NIHSS score47). 38 Moreover, the frequency of in-hospital medical complications and death after stroke is higher among women; and if surviving, disability outcomes are more severe.39,40 This is likely because women are older at stroke onset, and more likely to live alone, thus prehospital delay may be longer for women. 8 These differences in disability outcomes are observed for eating, dressing, grooming, and transfer from bed to chair, which may also be responsible for the higher poststroke institutionalization of women. 41
SEX-SPECIFIC PATHOPHYSIOLOGIC MECHANISMS
There is growing evidence of neuroprotective mechanisms of sex steroid hormones in women, supporting the observational epidemiology related to menopause. In this section, we will evaluate current evidence regarding the pathophysiology of stroke with emphasis on differences between women and men and the involvement of sex hormones, genetic, and other factors.
Pathophysiology of Stroke
A common feature of all stroke subtypes is impaired oxygen and nutrient delivery to areas upstream of the stroke site, causing catastrophic effects on brain tissue within minutes to hours of onset. The amount of time from stroke onset to cellular death is related to degree of decrease in blood flow. Cerebral blood flow (CBF) < 10 mL/100 g per minute causes rapid (within minutes) death of most cells, termed as the ‘ischemic core’; while a CBF between 10 and 20 mL/100 g per minute causes neurons to become functionally impaired but remain structurally intact and revivable after blood flow restoration, this area is called the ‘ischemic penumbra’. 42 In the ischemic core, oxygen and glucose deprivation lead to a rapid decline in neuronal ATP production, 43 which triggers the start of a series of biochemical events known as the ‘ischemic cascade’. 44 Briefly, after ATP deprivation, the membrane loses its ionic gradient, which is reflected by increases in cytoplasmic Na+ and Ca2+. This increase triggers glutamate release, causing even more Ca2+ influx via NDMA and AMPA receptors until reaching the excitotoxic threshold. Subsequently, degeneration of organelles and membrane integrity and dissolution of the cell (necrosis) occur. In the ischemic penumbra, reduced blood flow is insufficient to cause neuronal death. However, high extracellular glutamate released by the ischemic core causes accumulation of intracellular Ca2+ and activation of Ca2+ -dependent enzymes such as apoptosis-inducing calpain and caspases in the penumbra. 45 In addition, some of these Ca2+ -dependent enzymes produce, e.g., nitric oxide (NO), arachidonic acid, and superoxide that induce apoptosis.44,45
Sex Steroid Hormone-Dependent Mechanisms
Sex hormones, such as estrogen, progesterone, and testosterone, influence physiologic (e.g., vascular reactivity, CBF, and blood–brain barrier) and pathophysiologic (e.g., atherosclerosis) aspects of cerebral circulation.
46
One of the most extensively studied sex steroid hormones in relation to the physiology and pathophysiology of the circulatory system is the female hormone, estrogen. There is a large amount of evidence that estrogen, particularly 17β-estradiol (E2), is protective against cellular death in premenopausal stroke.
47
Vascular smooth muscle and endothelial cells express specific receptors and metabolic enzymes that interact with sex steroids, triggering genomic and nongenomic effects.
46
Genomic actions of estrogen are mediated through nuclear estrogen receptors (ERs) α and β, which are widely expressed, including on cerebral arterial endothelial and smooth muscle cells (SMCs).
48
On binding estrogen, the conformation of ERα/β changes, leading to recruitment of coactivators, which bind
Total NO production and release is higher in women compared with men and may underlie sex differences in vascular function. 51 NO-dependent mechanisms, such as myogenic tone, are involved in the ‘autoregulation’ of CBF, an important feature of the cerebral circulation for protection against fluctuating blood flow and pressure. Differences in myogenic tone in response to increasing BP were shown in male and female rats and mice that were untreated, ovariectomized (Ovx), or Ovx with estrogen replacement (Ovx + E2). 52 Lower myogenic activity was observed in untreated females and the Ovx + E2 group.
Estrogen also modulates vascular reactivity through cyclooxygenase-dependent prostanoids. 46 Ovx female rats that were chronically treated with E2 exhibited more dilated arteries compared with untreated rats. This difference contributed to a shift in cyclooxygenase-dependent production from vasoconstricting prostaglandin (PGH2) to vasodilating prostacyclin (PGI2). 53 Surprisingly, E2 seems to diminish EDHF-mediated dilations in female rats. 54 This could be presumably because of a higher NOS production since several studies implicated a compensatory role for EDHF when NO is deficient. 55
Together, these results suggest that E2 and most likely also other estrogens contribute to the regulation of different vasoactive factors such as NO, PGI2, and EDHF (see Figure 1). These compounds will promote better tissue perfusion in women than in men during and after cerebral artery occlusion 56 resulting in smaller stroke volumes. 57 Other effects of estrogen are affecting CBF, 58 decreasing permeability of the blood–brain barrier, 59 reducing oxidative stress and increasing neurogenesis and angiogenesis. 47 However, estrogen also generally affects pathophysiologic processes of the vascular system. There is compelling evidence that estrogen suppresses different aspects of atherogenesis. 60
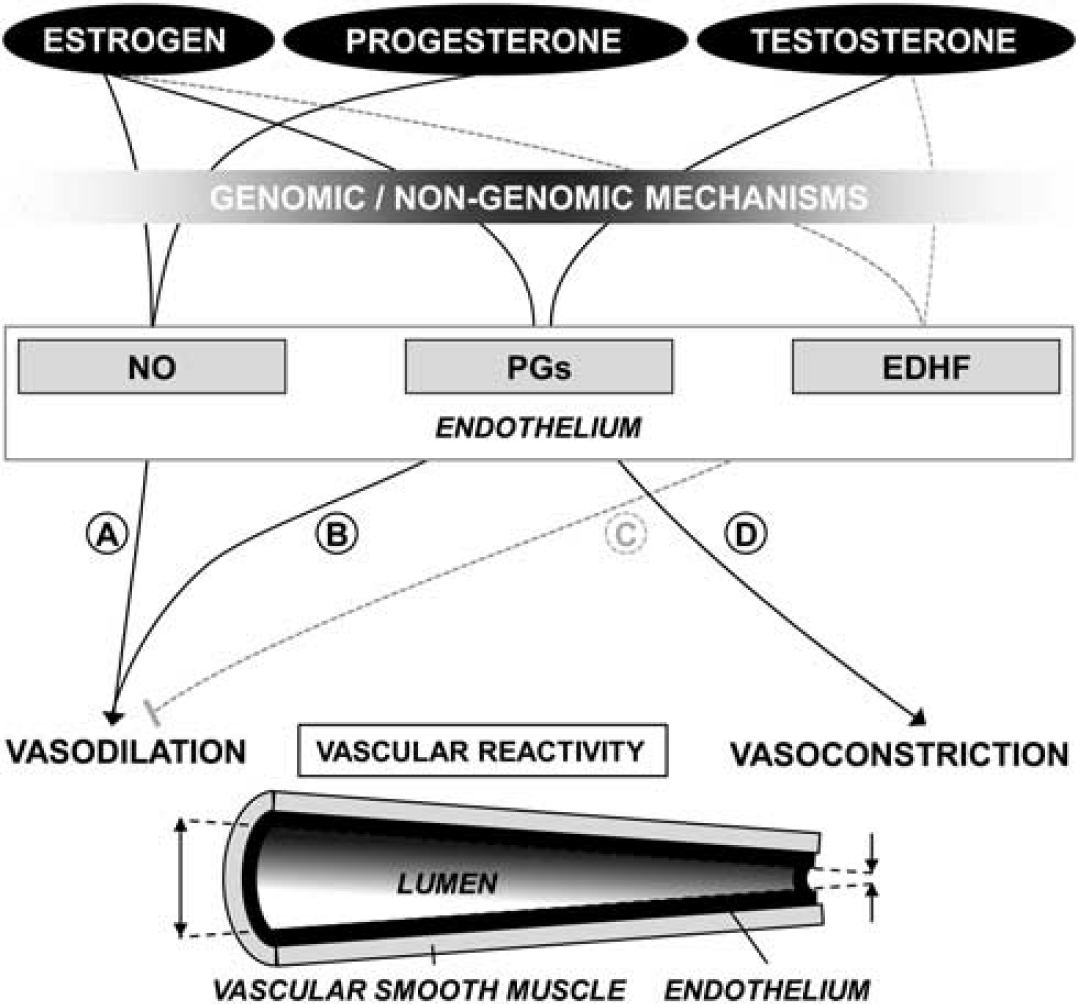
Observed sex steroid hormone effects on vascular reactivity. Estrogen and progesterone promote vasodilation while testosterone has opposing effects. Vasodilation is induced through increased nitric oxide (NO) production by estrogen and progesterone (
Formation of atherosclerotic plaques is inhibited by estrogen through many actions. Animal studies show that estrogen causes (1) antiproliferation of vascular SMCs, (2) decreased lipoprotein(a) accumulation and oxidation, (3) reduced monocyte adherence via modified expression of adhesion molecules (e.g., vascular cell adhesion molecule-1) and apical NO, (4) inhibition of monocyte differentiation, and (5) prevention of platelet thrombi. An extensive review concerning these and other influences of estrogen on atherosclerosis has been compiled by Nathan and Chaudhuri. 60 Only a small aspect of data related to the influence of estrogen on pathophysiologic functions is highlighted here. According to animal studies, it appears that higher levels of circulating estrogen in females are largely responsible for protection against cerebral and peripheral vascular diseases. This is also suggested by the increased stroke risk among postmenopausal versus premenopausal women, 7 which is likely related to decreases in estrogen levels during menopause. The menopausal transition, or perimenopause, is known to have a variety of effects on female physiology 61 and heralds onset of several vascular risks. 26
Progesterone also exhibits different properties with regard to cerebral and peripheral vascular function and dysfunction, when binding to its receptor, which is expressed in vascular SMC and endothelial cells.62,63 Rats treated with progesterone 4 weeks after ovariectomy experienced restored NO-mediated control of vascular tone in the mesenteric artery. 64 In addition, endothelial nitric oxide synthase expression was induced in human endometrial epithelial cells (HES) by progesterone through a nuclear progesterone receptor-mediated mechanism. 65 Like estrogen, progesterone inhibited cultured human vascular SMC proliferation, 66 which is important in atherogenesis. Several other effects of progesterone include decreased inflammation, reduced oxidative stress, and decreased edema. 47 These mechanisms of action may be responsible for progesterone's neuroprotective properties. Experiments with reproductive senescent female (RSF) rats revealed that progesterone-treated RSF rats had smaller cortical infarcts compared with untreated RSF rats. 67 This reduction was also observed in Ovx rats treated with progesterone both before middle cerebral artery occlusion and during reperfusion. 68 In addition, combined estrogen and progesterone administration to RSF rats attenuated cortical and total brain ischemic injury. 69 Reduced cortical infarct volume was also observed in Ovx rats through combined estrogen and progesterone treatment. 70 Despite the experimental evidence, the beneficial effects of estrogen and progesterone in women remain controversial. The WHI (Women's Health Initiative) reported a significant 31% increase in stroke risk in 16,608 healthy postmenopausal women after ingestion of estrogen + progestin for 5.6 years. 71 These results are in sharp contrast to observed protective effects of estrogen and progesterone in animal studies. However, as with any clinical trial in humans, there are caveats when interpreting the WHI data 72 related to inclusion and exclusion criteria, as well as experimental design. In WHI, it is important to be aware of the advanced age of the women when the intervention occurred, timing of hormone exposure in relation to menopausal age, hormonal preparations, differences in acute and long-term effects of hormonal preparations, and stroke susceptibility. These considerations are not unique to clinical trials in human. Similar phenomena have also been observed in Ovx RSF rats. Estrogen treatment increased cortical and striatal infarct volumes significantly compared with untreated Ovx RSF rats. 73 In contrast, estrogen treatment reduced neuron loss in another study, an effect that was blocked by continuous co-administration of progesterone. 74 Furthermore, progesterone exacerbated striatal stroke injury in Ovx female rats 75 confirming the complexity of steroid-mediated mechanisms. Divergent data may be linked to differences in choice of animal model, gender, age, hormone type and dose, and duration and timing of treatment. Ongoing follow-up studies should provide clarification.
Neuroprotective mechanisms attributed to estrogen and progesterone are less apparent in men compared with women, because men have lower circulating levels of these hormones. Therefore, men may be more influenced by the actions of androgens, most importantly testosterone. However, much less is known about the possible effects of testosterone on vascular functionality. Cellular signaling by testosterone occurs through the androgen receptor, which is expressed on cerebral blood vessels. 76 Via genomic and nongenomic pathways several physiologic functions, such as vascular reactivity, HTN, and atherosclerosis, are affected by the dominant bioactive form of testosterone, dihydroxytestosterone. 77 In contrast to estrogen, testosterone administration to orchiectomized male rats, shifts cyclooxygenase-dependent prostanoid production toward vasoconstricting thromboxane (TxA2), causing enhancement of vascular tone in cerebral arteries. 78 Moreover, TxA2 possesses potent platelet-aggregating actions that may promote thrombogenesis. 79 Similarly to estrogen, 54 though less pronounced, testosterone reduces EDHF contribution to vascular resting tone in orchiectomized rats. 80
Removal of androgens by castration provides protection from ischemic injury but is restored after testosterone replacement. 81
Data obtained with spontaneously hypertensive rats (SHRs) reveal a testosterone-mediated exacerbation of HTN in male SHRs. Female Ovx SHRs chronically treated with testosterone manifest increased BP levels comparable to those observed in male SHRs. 82 Androgens positively influence atherogenesis by (1) promoting vascular SMC proliferation, 83 (2) increasing monocyte adherence via enhancing vascular cell adhesion molecule-1 expression, 84 and (3) upregulating atherosclerosis-related genes, for example, those involved in lipoprotein processing, cell-surface adhesion and coagulation and fibrinolysis, in male but not in female macrophages. 85 Conversely, androgen deficiency promotes pathogenic processes leading to atherosclerosis (e.g., oxidative stress, endothelium dysfunction, and inflammation proliferation) primarily by altering lipid profiles (increased levels of triglycerides, cholesterol, LDLs, and decreased levels of high-density lipoproteins). 86 To conclude, it is far from clear whether the effects of androgens are positive or negative on stroke pathophysiology. It should also be remembered that testosterone is converted to E2, which may account for the observed beneficial effects of testosterone. 47
Sex Steroid Hormone-Independent Mechanisms
Genomic factors may also contribute to differences in stroke pathology between women and men. A recent study by Tian et al (2012) suggests that sex-specific expression of genes related to poststroke immune, inflammatory, and cell death responses may be responsible for differences between women and men.
87
For example, testosterone appears to induce expression of genes accounting for cell death through enhanced inflammation, dysregulation of blood–brain barrier and extracellular matrix, apoptosis and ionic imbalance.
81
In addition, genes on the Y-chromosome are partially involved in the higher BP and HTN observed in men. Human population studies reveal a Y-chromosome polymorphic
The vascular anatomy is also different among women and men. Women have smaller arteries and hearts than men, partly because of a smaller body size. The larger body size of male humans and rats compared with their female counterparts leads to enlargement of the left atrium which in humans is correlated with an increased risk for stroke onset. 92
Sex differences observed in stroke epidemiology and pathophysiology in both humans and animals may be attributed to several factors, including traditional risk factors as well as physiologic (genetic, hormonal, and anatomic) factors. In human populations, sex differences in lifestyle factors such as physical activity levels, levels and patterns of dietary intake, and smoking behavior may contribute to the risk for stroke (see Figure 2).
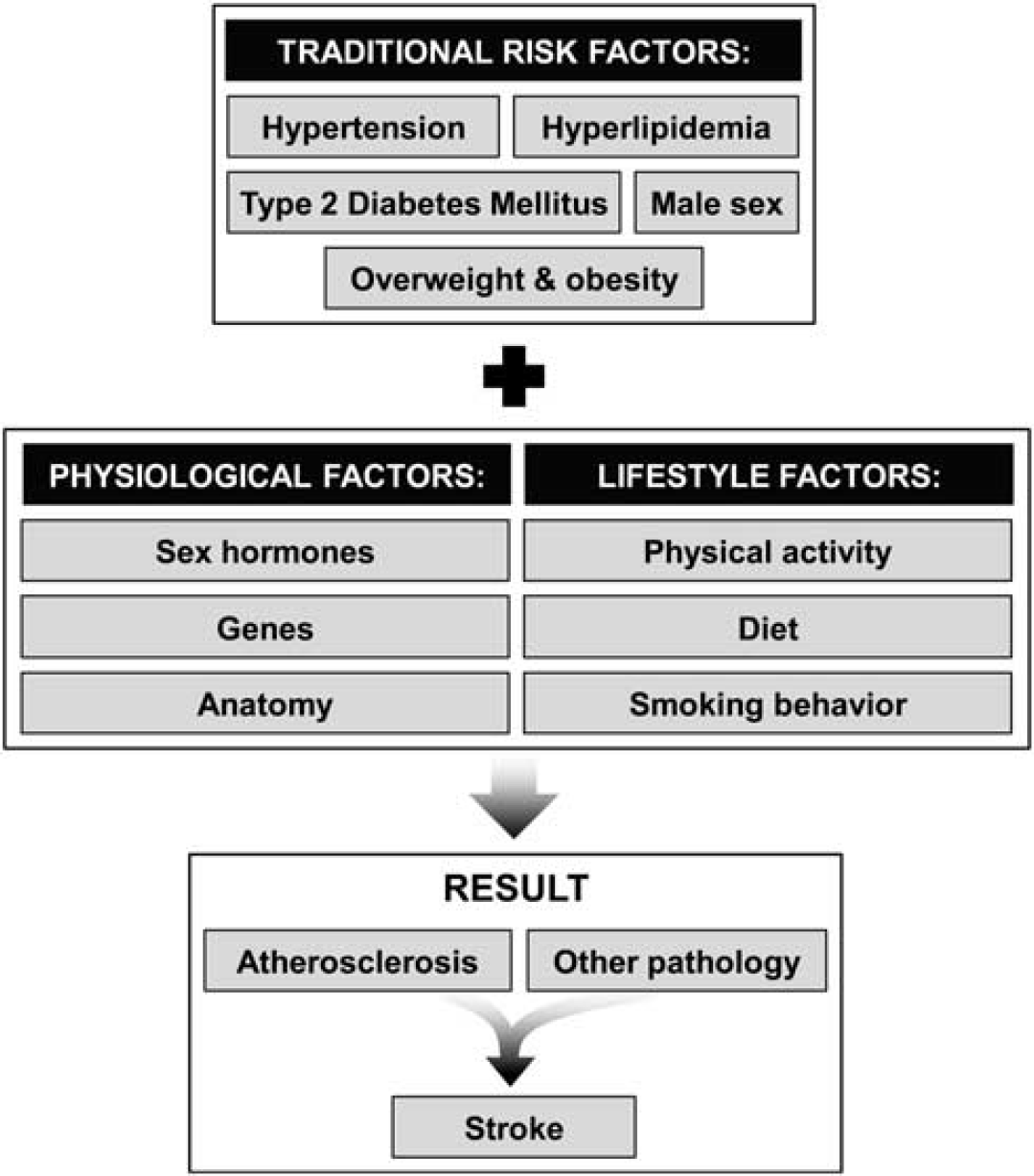
The dynamic interplay among factors related to stroke onset.
DIFFERENCES IN STROKE TREATMENTS AND OUTCOMES
There is growing evidence, although variable and inconsistent, of sex differences in stroke treatments and outcomes.38,93 In part, this is explained by differences in prestroke characteristics and clinical presentation. As described earlier, women are older at stroke onset and more likely to live alone, therefore evidencing longer prehospital delays after symptom onset compared with men. 8 The first response of others witnessing a stroke is an independent predictor of delay time. 94 In addition, women evidence worse prestroke disability and higher institutionalization rates. At clinical presentation, women have shown to be significantly less likely suffering dysarthria, ataxia, or paresthesia than men, but more often had urinary incontinence, loss of consciousness, visual deficits, and dysphasia.38,95 The presence and number of morbidities accompanying stroke also differs by sex, which is important for choice of secondary prevention. 95
Studies report a variety of acute and chronic stroke treatments—thrombolysis, stroke unit care, carotid surgery, rehabilitation, and forms of tertiary prevention. Secondary outcomes also vary and include quality of life, depression, dependency and addictions to drugs and alcohol, recurrent stroke, and death.8,20 Appelros
With regard to functional outcomes after stroke, women suffer more severe strokes than men 38 resulting in a lower quality of life (e.g., more physical limitations and decreased mental health) and higher dependency afterward. This leads to more discharges to nursing homes. 97 Furthermore, poststroke depression is more common among women than men. These differences in outcomes might be linked to demographic factors, poorer prestroke (psychosocial and physical) functionality, multimorbidities and less social support.7,8
To conclude this section, gaining insight into sex-related differences with respect to risk factor epidemiology, clinical presentation, treatment, and outcome will provide opportunities for better prevention and optimization of treatment by sex. For example, better monitoring of women transitioning through the menopause in relation to vascular risk (i.e., trends in BP, body weight, and blood lipids) and compliance to treatment regimens related to stroke risk would be beneficial. This will positively influence quality of care and outcome.
ANIMAL MODELS FOR SEX-SPECIFIC DRUG DISCOVERY IN STROKE
Given discrepant outcomes of stroke in women versus men, better clinical research is needed, as is further research using animal models to develop sex-specific treatments for stroke. Animal models provide a fundamental tool for this by mimicking (1) stroke against the background of different biologic risk environments (e.g., models of atherosclerosis, hypercholesterolemia, or arterial HTN) 98 or (2) the effect of stroke on physiologic functionality by cerebral occlusion (e.g., middle cerebral artery occlusion). 99 For example, hypercholesterolemia and atherosclerosis are induced in apolipoprotein E-deficient (apoE−/–) and LDL receptor-deficient (LDLR−/–) mice and these processes are enhanced when animals are fed a Western-type diet. 100 The SHR stroke-prone model is the most widely used animal model in stroke-related HTN studies since 80% of SHR-RP rats experience stroke within 8 to 13 months. 101 Despite the lack of a decent animal model that simulates perimenopause, which affects female physiology and facilitates the onset of vascular risk, 61 the accelerated ovarian failure model and RSF rats model seem promising. In the latter RSF model, natural aging of female rats causes hormonal fluctuations and so-called estropause. This model confirms the neuroprotective properties of female hormones since differences in infarct size after middle cerebral artery occlusion disappeared in middle-aged RSF versus age-matched male rats, 67 but failed to achieve significant low levels of estrogen as observed in women. In contrast, in the accelerated ovarian failure model, follicle depletion is induced by repeated 4-vinylcyclohexene (VCD) injection and results in a significant decrease in estrogen levels over a period of time. 102 Yet, studies using this model with stroke induction are still absent. Overall, this first group of models provides opportunities to investigate the effect of sex steroid hormone (in combination with ovariectomy and orchiectomy) and pretreatment on pathologic factors related to stroke. The second group of models includes mechanical occlusion of the MCA, embolic models (injection of large-sized synthetic thrombi) and inducing endothelial damage through phototrombosis. 103 These are especially suitable for testing therapeutic methods, but also for studying cellular consequences and functional outcome after stroke.
Standardization of stroke-related animal research is important since this may contribute to the diversity of results observed regarding the effectiveness of stroke treatments in humans versus animals and females versus males. Therefore, the STAIR (Stroke Treatment Academic Industry Roundtable) provided guidelines and standards to improve the translational applicability of current animal stroke models. 104 Attention should be paid to guidelines related to dose of drug used, window of opportunity, choice of animal model, physiologic monitoring, and outcome measures. Despite the establishment of these STAIR criteria over 10 years ago, in 1999, no optimal animal model is available that mimics perimenopause. However, the accelerated ovarian failure model potentially allows studying optimal approaches (i.e., dose, window of opportunity) for restoring sex hormone levels to reduce stroke risk in women. 105
CONCLUSION AND FUTURE PERSPECTIVES
This review highlights important sex differences in the epidemiology, pathophysiology, treatments, and outcomes related to stroke. Epidemiologic studies have revealed a clear age-by-sex interaction leading to several mechanistic hypotheses of stroke risk and onset. Premenopausal women appear less vulnerable to stroke than similarly aged men. However, after menopause the m/f ratios for prevalence and incidence decrease, indicating an increase in stroke among postmenopausal women (or decrease in men). This shift is reflected in mortality and case fatality rates, which are higher for women at older ages. When evaluating these data it should be taken into account that women have longer life expectancy, are older at stroke onset, and suffer more severe strokes. These two latter observations may be because of the gradual accrual of aging-related risk factors. The prevalence and severity of these risk factors is different for men and women. Women experiencing stroke show higher prevalence of HTN and atrial fibrillation; while men experience more heart disease, T2DM, and alcohol and tobacco use. In addition, women appear to have a worse prognosis after stroke, such as a lower quality of life and more poststroke depression.
Premenopausal women are most likely protected against stroke because of sex steroid hormone-dependent mechanisms. This is a natural conclusion, since there are dramatic changes in the female sex hormone milieu before, during, and after menopause. Estrogen, testosterone, and progesterone affect different physiologic and pathophysiologic functions of the cerebral circulation. Estrogen promotes blood flow by decreasing vascular reactivity while testosterone has opposite effects. Both are involved in the development of atherosclerosis, where inconsistent effects are reported. Clarifying the impact of sex hormones on the cerebral circulatory system using suitable animal models, for example, those mimicking the perimenopausal stage is essential to elucidate female–male differences in stroke pathophysiology and for developing sex-specific treatments. Not to be forgotten are other factors, such as anatomic and genetic factors, which may also contribute to observed differences.
Much remains to be learned about differences in stroke between women and men. In the end, women are affected most negatively by stroke. Further research is needed to improve stroke risk profiles and treatments for both women and men.
DISCLOSURE/CONFLICT OF INTEREST
The authors declare no conflict of interest.