
Abstract
Recent stem cell technology provides a strong therapeutic potential not only for acute ischemic stroke but also for chronic progressive neurodegenerative diseases such as Alzheimer's disease, Parkinson's disease, and amyotrophic lateral sclerosis with neuroregenerative neural cell replenishment and replacement. In addition to resident neural stem cell activation in the brain by neurotrophic factors, bone marrow stem cells (BMSCs) can be mobilized by granulocyte-colony stimulating factor for homing into the brain for both neurorepair and neuroregeneration in acute stroke and neurodegenerative diseases in both basic science and clinical settings. Exogenous stem cell transplantation is also emerging into a clinical scene from bench side experiments. Early clinical trials of intravenous transplantation of autologous BMSCs are showing safe and effective results in stroke patients. Further basic sciences of stem cell therapy on a neurovascular unit and neuroregeneration, and further clinical advancements on scaffold technology for supporting stem cells and stem cell tracking technology such as magnetic resonance imaging, single photon emission tomography or optical imaging with near-infrared could allow stem cell therapy to be applied in daily clinical applications in the near future.
Despite numerous studies and active challenges for the treatment of major neurologic diseases such as ischemic stroke, Alzheimer's disease, Parkinson's disease, amyotrophic lateral sclerosis, and multiple sclerosis, only a few treatments have ameliorated the neurologic symptoms with conventional symptomatic therapies for neurorepair. Among various organs, brain is the most sensitive organ to various injuries such as ischemia, hypoglycemia, infection/inflammation, trauma, aging, and degeneration. In the brain, neurons are particularly sensitive to such injuries, and the vulnerability is different even within the neuronal populations (Abe et al, 1991, 1995; Abe and Kogure, 1993). These vulnerabilities of neurons make it difficult to cure patients suffered from the above diseases in clinical settings.
However, the stem cell approach could provide an alternative choice for neuroregeneration and disease-modifying therapy. For neuroregenerative therapy, the activation of intrinsic neural stem cells (NSCs) or the exogenous transplantation of NSCs/neural progenitor cells (NPCs) can be applied (Abe, 2000; Iwai et al, 2003). To support stem cell migration, an artificial scaffold can be implanted into the injured brain for promoting ischemic brain repair and regeneration (Deguchi et al, 2006). The addition of neurotrophic factors greatly enhanced the intrinsic migration or invasion of stem cells into the scaffold, which could provide a future regenerative potential against ischemic brain damage at the chronic stage (Zhang et al, 2008). Especially, granulocyte-colony stimulating factor (G-CSF) is regarded as a promising drug candidate which can reduce neuroinflammation and potentiate both neurogenesis and angiogenesis after ischemic stroke by promoting bone marrow (BM) cell migration into the ischemic brain (Sehara et al, 2007a, 2007b). Recent studies have demonstrated that cord blood mononuclear cells, BM mononuclear cells, and BM stromal cells (BMSCs) can survive in postischemia tissue, and reduce neuronal damage when transplanted into rodents subjected to cerebral infarction (Brenneman et al, 2010; Chen et al, 2006; Hokari et al, 2008; Prockop et al, 2003). Recent studies suggest an important interaction between neuronal cells and vascular component as neurovascular unit and a potential therapeutic target for ischemic stroke, Alzheimer's disease, and amyotrophic lateral sclerosis (del Zoppo, 2009; Kurata et al, 2011; Miyazaki et al, 2011; Yamashita et al, 2009; Zlokovic, 2010). In addition to necrosis and apoptosis, additional neuronal cell death mechanisms such as autophagy and transcriptional repression-induced atypical death have recently been pointed out as important for the therapeutic approach of both neurorepair and neuroregeneration (Morimoto et al, 2009; Tian et al, 2011). Potential therapeutic benefits of stem cell therapy on these neurovascular units and additional cell death mechanisms are future topics to be studied. The purpose of this review is to summarize the current progress of basic stem cell science and its early clinical applications for advanced stem cell therapy.
Part 1. Intrinsic neurogenesis and exogenic stem cell transplantation
Intrinsic Neural Stem Cells
To supply new neurons into the infarcted brain, two tactics are proposed. One is the transplantation of extrinsic NSCs/NPCs derived from stem cells such as embryonic stem (ES) cells and induced pluripotent stem (iPS) cells. The other is the activation of intrinsic NSCs. It was already reported that persistent neurogenesis occurs in two restricted regions of the adult mammalian brain: the subgranular zone (SGZ) of the hippocampal dentate gyrus (Gage, 2000) and the subventricular zone (SVZ) of the lateral ventricle (Alvarez-Buylla and Garcia-Verdugo, 2002). In the SGZ, newly born neurons migrate into the granule cell layer and integrate into the existing neuronal network. In the SVZ, which is a thin cell layer in the lateral walls of lateral ventricles, NSCs continuously give rise to NPCs migrating into the olfactory bulb (Doetsch et al, 1999). To know whether the ischemic condition affects endogenous neurogenesis, we studied the temporal profile of NSC division, migration, and differentiation in the SGZ and the SVZ by using the transient forebrain ischemia gerbil model. We found that the ischemic condition increased the division of NSCs of the SGZ with a peak 10 days after ischemic induction, following which cells migrated into the granule cell layer and differentiated mainly into neuronal cells (Iwai et al, 2002; Figure 1A). Furthermore, we also found that transient forebrain ischemia enhances NSC proliferation in the SVZ with a peak 10 days after ischemia, leading to the migration of more NPCs into the olfactory bulb (Iwai et al, 2003). These results suggest that forebrain ischemia increased NSC number and resulted in increased neurogenesis, mostly in the two restricted lesions, the SGZ and the SVZ.
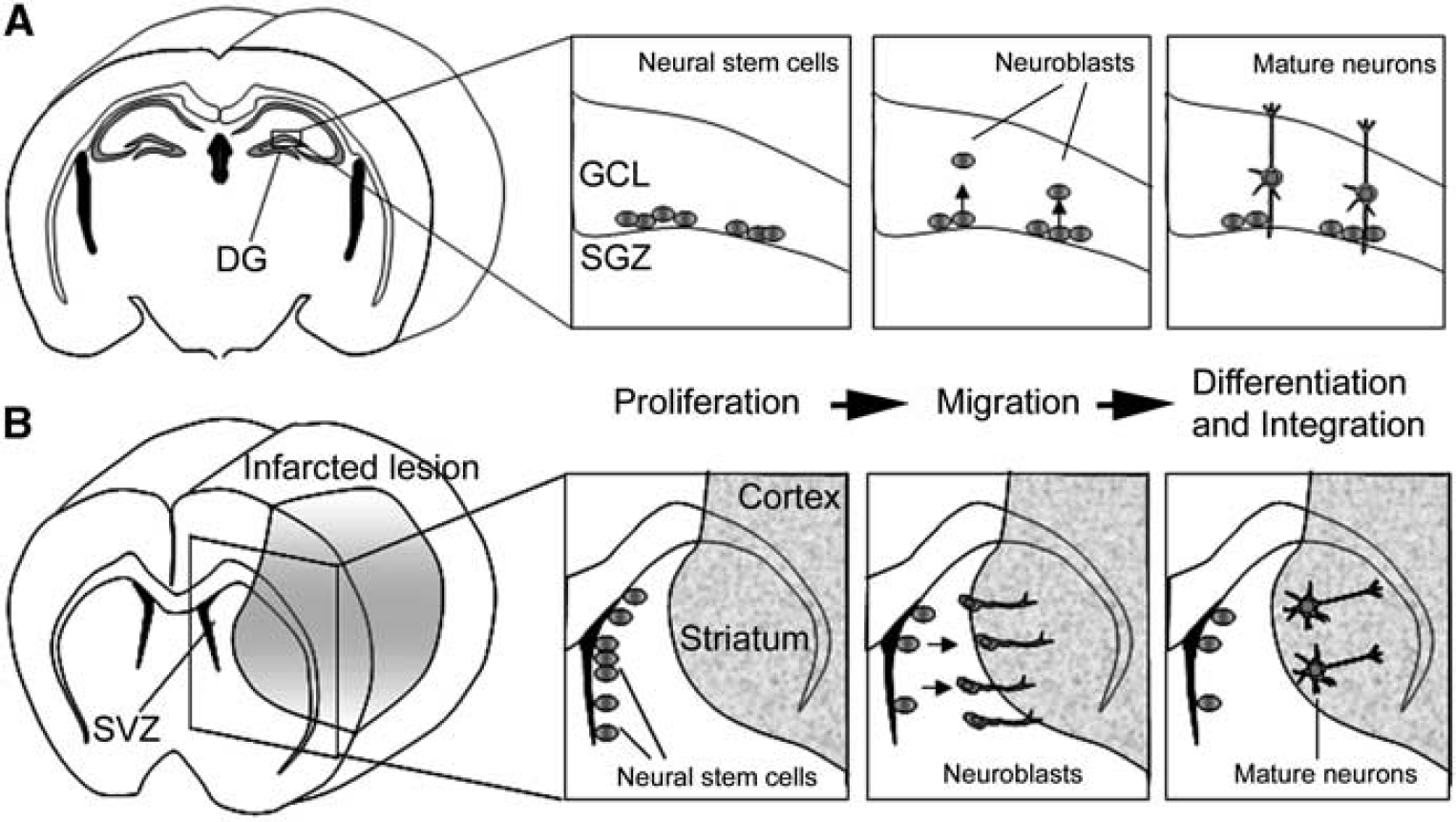
The stage of endogenous neurogenesis can be divided into three steps: proliferation, migration, and differentiation. (
Many researchers reported that newly born neurons can be found in the postinfarcted lesion including the striatum and cortex in another animal model, the transient focal ischemia model (Arvidsson et al, 2002; Teramoto et al, 2003), which is closer to the pathophysiology of human cardioembolic stroke. To clarify whether SVZ NSCs supply new neurons to areas injured by ischemia, several study groups have performed region-specific cell labeling and long-term tracing experiments. Subventricular zone-derived NPCs were also reported to migrate toward the injured striatum after middle cerebral artery occlusion. A long-term tracing study showed that the SVZ-derived NPCs differentiated into mature neurons in the striatum, in which they formed synapses with neighboring striatal cells (Yamashita et al, 2006; Figure 1B), implying that the SVZ has an important role as a cell source supplying newborn neurons to brain lesions damaged by focal ischemia. Recently, NPCs supplying GABAergic neurons were found even in the neocortical layer 1 of adult rats, and their proliferation was highly activated in the ischemic condition (Ohira et al, 2010).
In the postischemic brain, newly born neurons can be supplied from the SVZ, the SGZ, and the neocortical layer, but the number is too small for recovery of neurologic functions. For example, newly born neurons could replace only 0.2% of the dead striatal neurons even in the rat middle cerebral artery occlusion model (Arvidsson et al, 2002). Appropriate interventions need to be added to enhance the proliferation, survival, and neuronal maturation of intrinsic NSCs and their progeny, so as to use their intrinsic neural cell source for therapeutic purposes.
Cell Transplantation Therapy with Embryonic Stem, Induced Pluripotent Stem, and Induced Neuronal Cells
Human ES (hES) cells were first generated from the inner cell mass of blastocysts in 1998 (Thomson et al, 1998), and human-induced pluripotent stem (hiPS) cells were established by introducing four transcriptional factors (
Scientific reports showing that each specific type of neuron could be generated from human skin fibroblasts
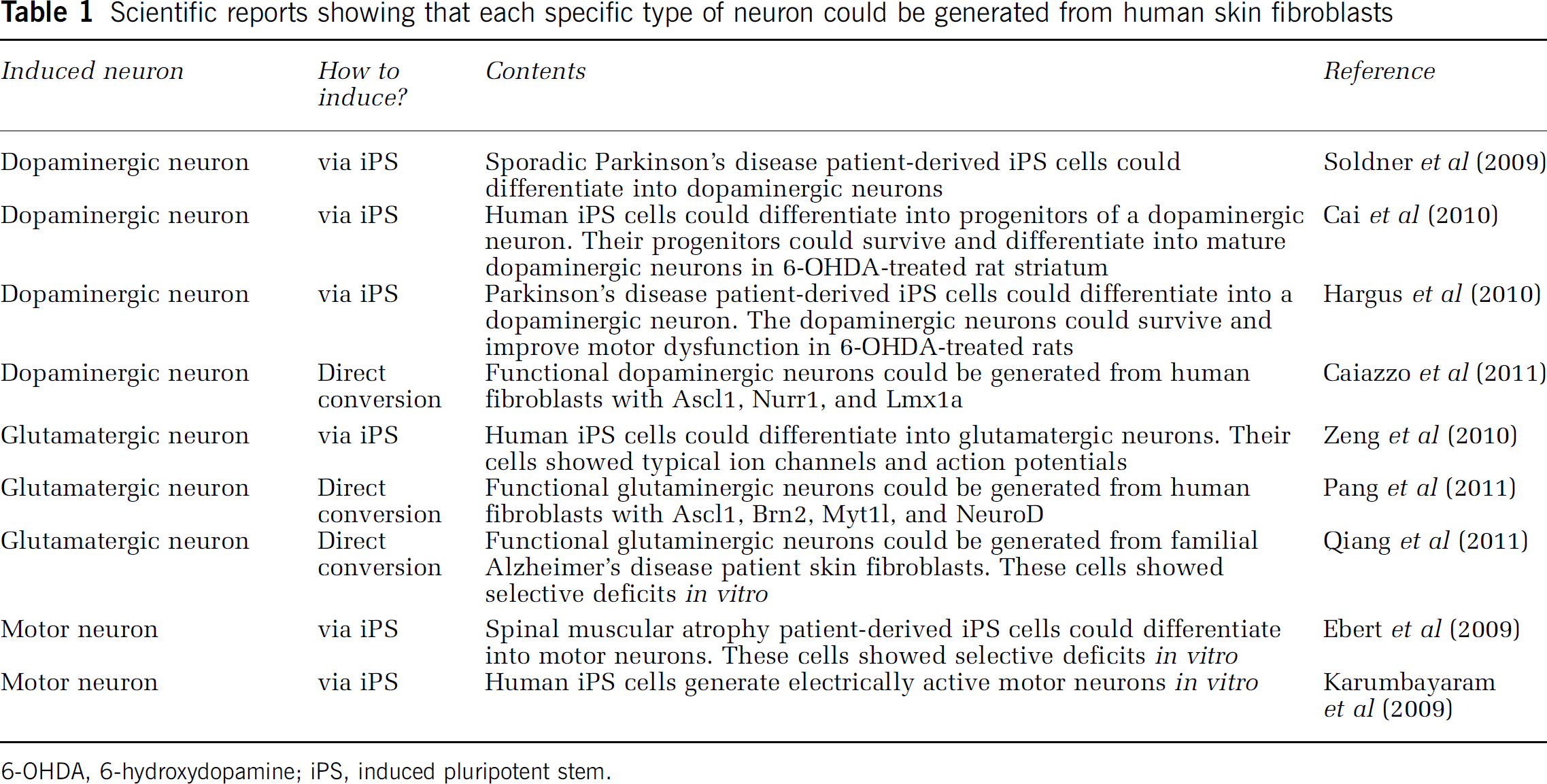
6-OHDA, 6-hydroxydopamine; iPS, induced pluripotent stem.
To achieve iPS cell therapy in a clinical setting, iPS cells tumorigenicity is a critical problem that clearly needs to be overcome. Germline-competent chimera mice with iPS cells developed tumors in which the integrated exogenous
Part 2. Granulocyte-colony stimulating factor in basic and clinical sciences
Research on stem and progenitor cells has the potential to yield new treatments for ischemic stroke, but transplantation of these cells faces a variety of problems, such as infection, rejection, and risk of malignancy, and there are also ethical and political issues (Lo and Parham, 2009). Granulocyte-colony stimulating factor, which is in widespread clinical use for treatment of chemotherapy-associated neutropenia (Cavallaro et al, 2000), is a new candidate for neuroprotection and neuroregeneration. As its profiles of pharmacological and adverse effects are well known, clinical application of G-CSF is expected to be straightforward, compared with stem/progenitor cell therapy. This review focuses on the neuroprotective and neuroregenerative properties of G-CSF in experimental ischemic models (Figure 2). The evidence from current clinical trials is also reviewed.
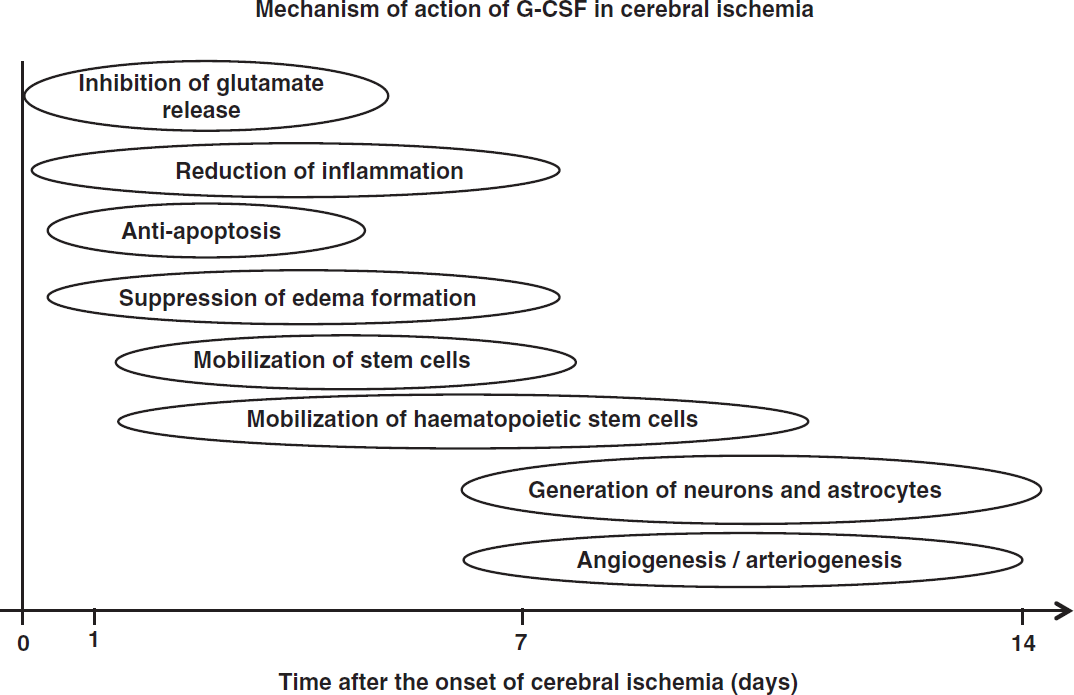
Mechanism of action of granulocyte-colony stimulating factor (G-CSF) in cerebral ischemia.
Neuroprotective Mechanisms of Granulocyte-Colony Stimulating Factor in Cerebral Ischemia
It is well established that G-CSF reduces infarct volume in experimental cerebral ischemia (Schäbitz et al, 2003; Schneider et al, 2005; Six et al, 2003). However, most studies assessed transient rather than permanent ischemia, and short-term rather than prolonged ischemia (England et al, 2009). In the acute phase of cerebral ischemia, the neuroprotective mechanisms of G-CSF include inhibition of glutamate release, reduction of inflammation, antiapoptosis activity, and suppression of edema formation.
Ischemia causes impairment of brain energy metabolism, with the release of excessive amounts of glutamate into the extracellular space. Granulocyte-colony stimulating factor can attenuate the release of extracellular glutamate in transient focal ischemia (Han et al, 2008). Further, G-CSF has a direct protective effect against glutamate-induced excitotoxicity in cultured neurons (Schäbitz et al, 2003). Both G-CSF and G-CSF receptors are widely expressed by neurons in the central nervous system (CNS), and their expression is upregulated in ischemia, suggesting the involvement of an autocrine protective signaling mechanism (Schneider et al, 2005). At 6 hours after cerebral ischemia, endogenously released G-CSF is presumably active on the upregulated G-CSF receptor in the penumbra, and may provide protection against apoptotic cell death of neurons. After interaction with G-CSF receptor through JAK signaling, G-CSF activates several antiapoptotic pathways involving signal transducer and activation of transcription-3, extracellular signal-regulated kinase, and phophatidylinositol 3-kinase-Akt (Schneider et al, 2005). Furthermore, G-CSF upregulates Bcl-2 and Pim-1 expression, and downregulates cytochrome c reductase translocation to the cytosol, Bax translocation to mitochondria, and the level of cleaved caspase-3 in neurons (Solaroglu et al, 2006).
Microglia/macrophages are considered to be major sources of inflammatory cytokines in the ischemic brain. Granulocyte-colony stimulating factor decreased the migration of BM-derived monocytes/macrophages into the peri-infarct and core regions (Komine-Kobayashi et al, 2006; Lee et al, 2005). It also suppressed inducible nitric oxide synthase activity and nitrotyrosine expression (Komine-Kobayashi et al, 2006). At 24 and 72 hours after transient focal ischemia, G-CSF also reduced expression of tumor necrosis factor-
In contrast, an increased amount of peripheral neutrophils may result in aggregate formation in cerebral microvasculature, leading to a breakdown of blood flow, and may worsen brain damage (del Zoppo and Mabuchi, 2003). A negative effect of G-CSF on outcome, associated with enhanced brain atrophy and an exaggerated inflammatory response, was also reported in permanent cerebral infarction (Taguchi et al, 2007). Although administration of G-CSF was accompanied by a significant increase of circulating neutrophils 2 days after ischemia, leukocytosis was restricted to the vessel compartment with no elevation of intraparenchymal neutrophil count, and there was no deleterious effect on lesion formation or functional recovery (Strecker et al, 2010).
Neuroregenerative Mechanisms of Granulocyte-Colony Stimulating Factor in Cerebral Ischemia
In the subacute phase (and partly also the chronic phase) of cerebral ischemia, neuroprotective mechanisms include the mobilization of hematopoietic stem cells, generation of neurons and astrocytes, angiogenesis and arteriogenesis.
Granulocyte-colony stimulating factor mobilizes stem cells from BM to circulating blood. CD34+ hematopoietic stem cells from BM migrated into the injured brain when administered intracerebrally or intravenously (Sykova and Jendelova, 2005). It improves the recovery of motor function in spinal cord injury models (Urdzikova et al, 2006). Piao et al (2009) reported that numerous BM-derived cells migrated into the brain parenchyma when G-CSF+ stem cell factor (SCF) was applied 16 weeks after ischemia. However, G-CSF+SCF treatment in the subacute phase predominantly increased BM-derived microglia, and slightly increased neuronal and endothelial cells, in the peri-infarct region (Kawada et al, 2006). The migration of BM-derived monocytes/macrophages was slightly reduced in the peri-infarct region after G-CSF treatment (Komine-Kobayashi et al, 2006). Thus, the capacity of BM-derived cells to restore function in the injured brain has been demonstrated, but there is some doubt as to whether mobilization of stem cells is the main neuroprotective mechanism of G-CSF.
Cerebral ischemia induces the generation of new neurons, which may potentially be used to restore brain function, from precursor cells. Administration of G-CSF immediately after middle cerebral artery occlusion increased newly generated neurons in the SGZ (Schneider et al, 2005; Sehara et al, 2007b). Administration of G-CSF+SCF in the subacute phase (days 11 to 20) after middle cerebral artery occlusion significantly stimulated proliferation of intrinsic NSCs/NPCs in the rostral SVZ, compared with administration in the acute phase (days 1 to 10) (Kawada et al, 2006). The subacute-phase treatment elevated the expression of IL-10 mRNA and antiinflammatory cytokines at 14 days after occlusion, suggesting that hematopoietic cytokine treatment in the subacute phase may provide a favorable microenvironment for neurogenesis (Morita et al, 2007). Even in the chronic phase of ischemia, significant functional improvement was seen at 1, 5, and 17 weeks after SCF+G-CSF treatment (Zhao et al, 2007). Thus, proliferation of intrinsic NSCs/NPCs is considered to be the main protective role of G-CSF rather than mobilization of stem cells.
Angiogenesis is a process in which new vessels arise from preexisting ones (Pandya et al, 2006). Lee et al (2005) found that G-CSF enhanced angiogenesis in stroke, measured in terms of endothelial cell proliferation, vascular surface area, number of branch points, and vascular length. Granulocyte-colony stimulating factor-induced angiogenesis may be caused by direct activation of brain endothelial cells (Bussolino et al, 1991) and mobilization of endothelial progenitor cells to the ischemic boundary zone. The effect of G-CSF was more pronounced when treatment was initiated earlier, but it remained even when treatment was delayed up to 7 days after ischemia (Lee et al, 2005). The G-CSF+SCF treatment during chronic stroke also elevated BM-derived endothelial cells (Piao et al, 2009), suggesting a prolonged therapeutic time window.
Arteriogenesis is a process in which preexisting collateral arterioles transform into functional collateral arteries. Granulocyte-colony stimulating factor promoted leptomeningeal collateral growth after common carotid artery occlusion, and increased circulating blood monocytes and Mac-2-positive cells, suggesting that mechanisms coupled to monocyte upregulation might be involved (Sugiyama et al, 2011c).
Current Clinical Trials of Granulocyte-Colony Stimulating Factor
There have been several clinical studies of G-CSF, as shown in Table 2. Shyu et al (2006) conducted a trial involving 10 patients with acute cerebral infarction using subcutaneous G-CSF injections (15
Current clinical trials of G-CSF for ischemic stroke
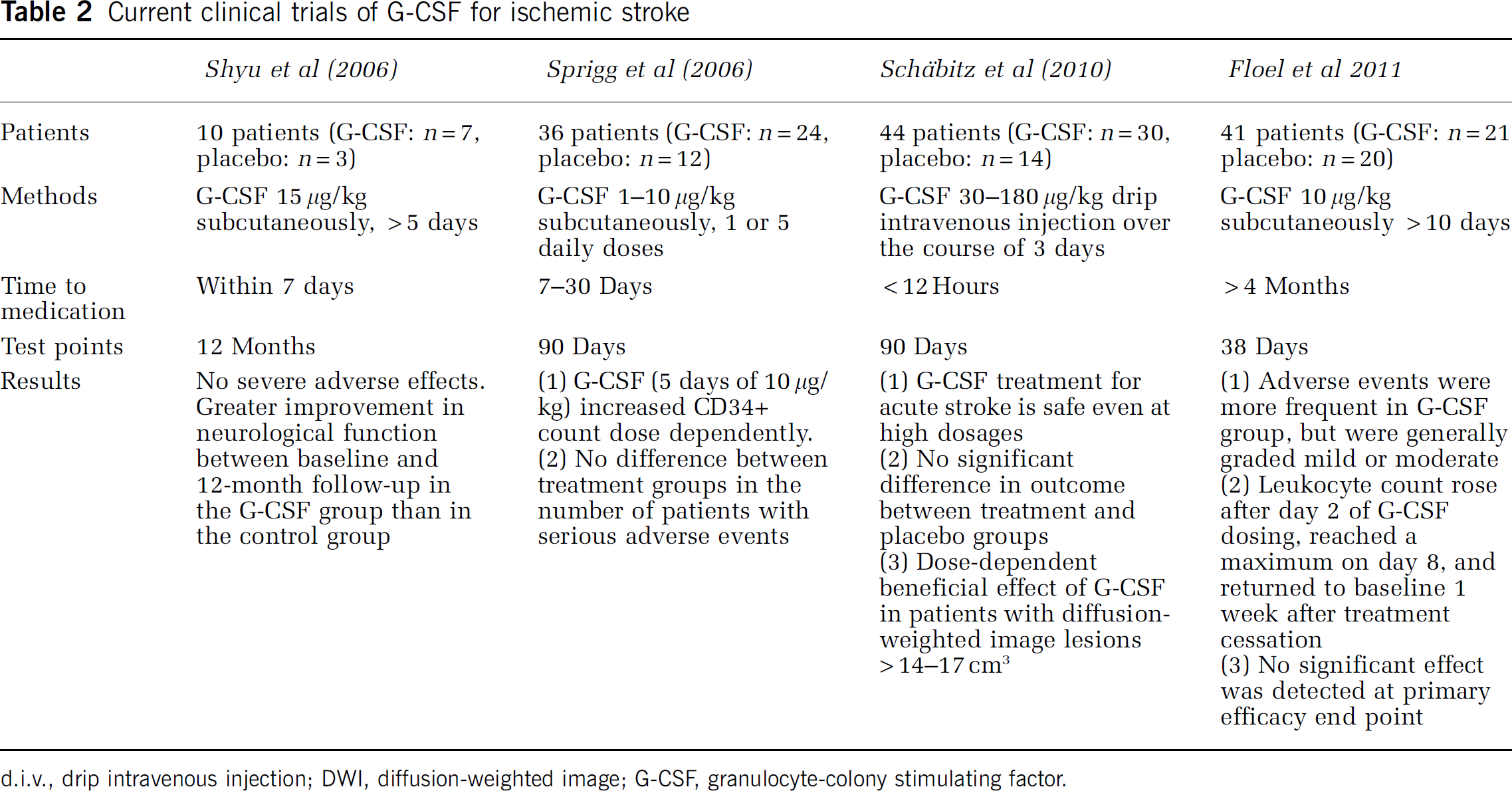
d.i.v., drip intravenous injection; DWI, diffusion-weighted image; G-CSF, granulocyte-colony stimulating factor.
In conclusion, it is unclear whether G-CSF has a neuroprotective or neuroregenerative effect in stroke patients. To clarify the optimal time and dose of G-CSF administration, further clinical studies with a larger number of patients enrolled, uniform infarct size, similar stroke subtype and background factors, lower extent of leukocytosis, among other factors, will be needed.
Part 3. Basic and translational aspects of bone marrow stromal cell transplantation for ischemic stroke
Basic Aspects of Bone Marrow Stromal Cell Transplantation
Although a huge number of preclinical and clinical tests were performed over these past 50 years, there are few drugs that are effective to protect or repair the damaged CNS due to ischemic stroke (Savitz and Fisher, 2007). However, recent decade studies have strongly suggested that cell transplantation therapy may potentially promote functional recovery after various kinds of CNS disorders, including cerebral infarct. A variety of cell types have been studied as cell source of transplantation into animal models of CNS disorders, including ES cells, NSCs, iPS cells, and BMSCs. Of these, BMSCs may have the most promising potential because they can be harvested from patients without posing ethical or immunological difficulties (Bliss et al, 2007; Parr et al, 2007). Bone marrow stromal cells are known to support the homing and proliferation of hematopoietic cells in the BM (Kortesidis et al, 2005; Uccelli et al, 2011). They differentiate into fat, bone, and cartilage, but can also transdifferentiate into embryological unrelated tissues, including neural cells (Sanchez-Ramos et al, 2000; Uccelli et al, 2011; Woodbury et al, 2000).
There is increasing evidence that transplanted BMSCs aggressively migrate toward the damaged CNS tissue and promote the recovery of motor function after cerebral infarct. Recent studies have shown that BMSCs also improve cognitive function due to chronic cerebral ischemia (Shichinohe et al, 2010). Now, they are considered to provoke these beneficial effects through their differentiation into neural cells and production of various kinds of cytokines or growth factors that can rescue the host neurons (Kuroda et al, 2011; Prockop et al, 2003). Thus, BMSCs
Recent
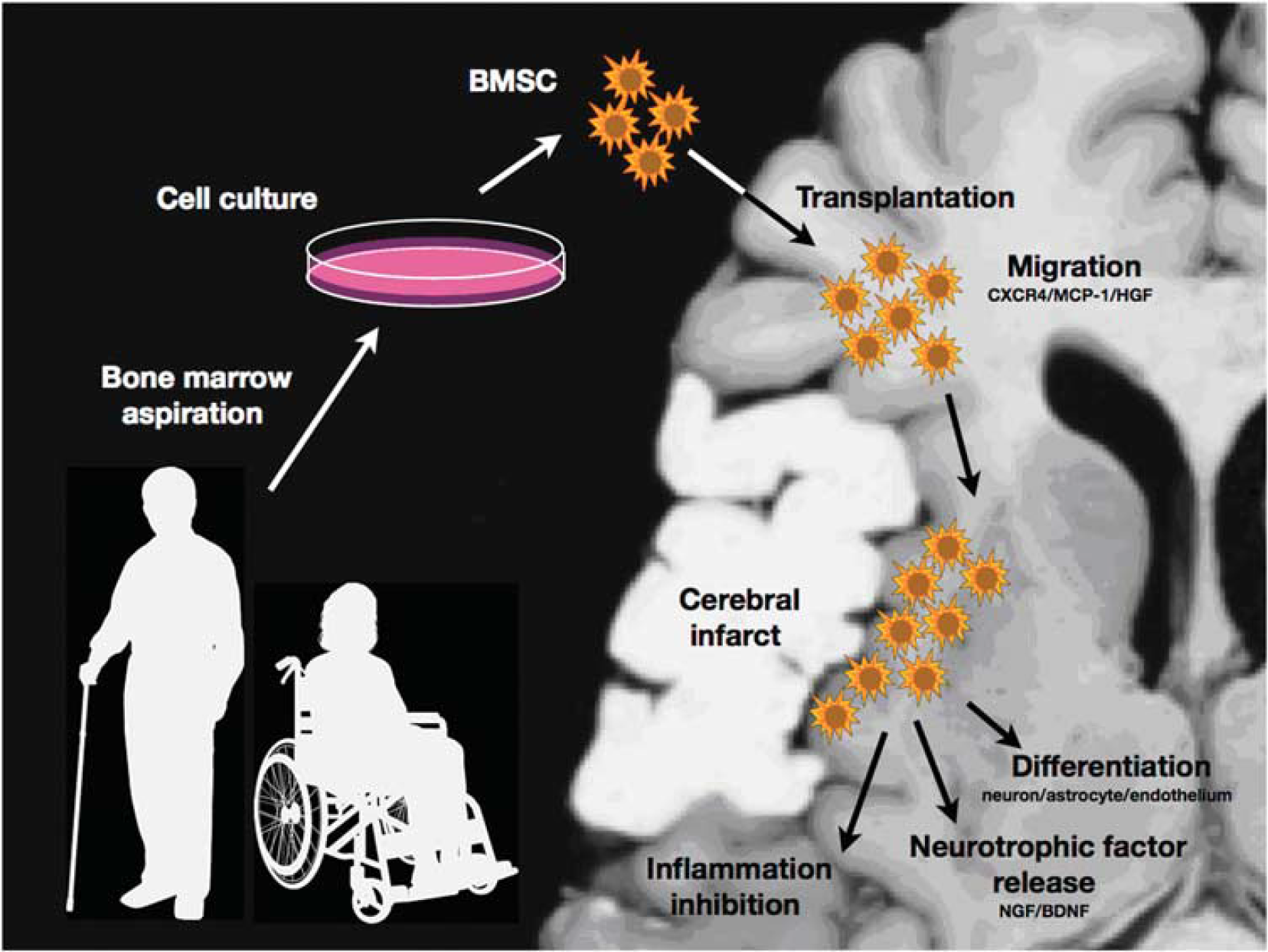
Scheme of bone marrow stromal cell (BMSC) transplantation for ischemic stroke. The engrafted cells migrate toward the peri-infarct area via chemokine interaction. They may rescue and repair the damaged central nervous system (CNS) tissue through the differentiation into the neural cells, the release of neurotrophic or neuroprotective factors, and the inhibition of inflammatory reactions.
Translational Aspects of Bone Marrow Stromal Cell Transplantation
As described above, the observations in basic experiments are encouraging. Some clinical trials of BMSC transplantation have already been initiated for patients with ischemic stroke. Bang et al (2005) intravenously injected autologous BMSCs into five patients with severe neurologic deficits due to ischemic stroke at 5 to 9 weeks after the onset, and concluded that autologous BMSC infusion is a feasible and safe therapy that may improve functional recovery. Honmou et al (2011) intravenously transplanted BMSCs into 12 patients with ischemic stroke 36 to 133 days after stroke. Lee et al (2010) performed an open-label, observer-blinded clinical trial of 52 patients with ischemic stroke, and followed them for up to 5 years. They concluded that intravenous transplantation of autologous BMSCs could be a safe and effective strategy for ischemic stroke. These studies indicate that BMSC transplantation may at least be safe and feasible for patients with ischemic stroke. However, we should clearly remind that there are no clinical trials that prove the clinical significance of cell-based therapy, including BMSC transplantation.
There are many variables that may affect the efficacy of BMSC transplantation in a clinical setting. As shown in Figure 4, these include donor cell factors (safety, autologous or allogeneic,
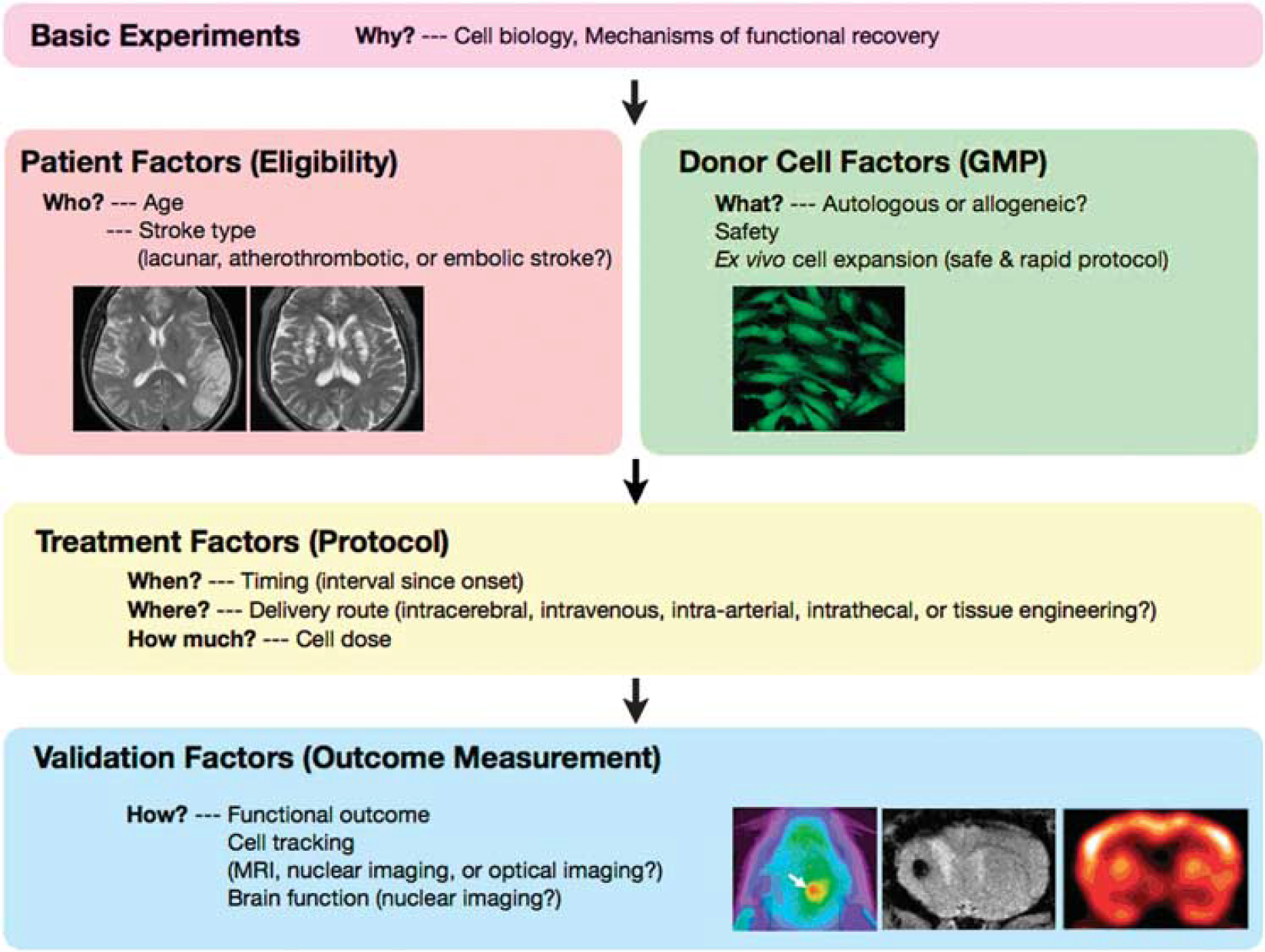
‘Five Ws and two Hs (5W2H)’ of cell therapy—the issues to be answered in preclinical studies and early-stage clinical trial of bone marrow stromal cell (BMSC) transplantation for ischemic stroke. GMP, good manufacturing practice; MRI, magnetic resonance imaging.
First, allogeneic cells would permit ‘off the shelf’ use even within 24 hours after the onset, but force a long-term medication of immunosuppressant. Therefore, the use of autologous donor cells would be ideal for patients if the following issues were to be resolved. Autologous BMSCs from patients themselves would be ideal as donor cells for restorative medicine, but they would require several weeks for
Second, BMSCs are transplanted within 24 hours or 7 days after the insults in the majority of animal studies, whereas they are usually transplanted several weeks (or even several months) after the onset in previous clinical trials (Bang et al, 2005; Honmou et al, 2011; Lee et al, 2010). Therefore, a considerable gap of treatment protocol exists between animal experiments and clinical trials, which may correspond to ‘inadequate preclinical testing’ (Savitz and Fisher, 2007). Granulocyte-colony stimulating factor may be useful to speed up BMSC expansion and shorten the interval between onset and transplantation therapy. Thus, a certain concentration of G-CSFs significantly enhances their proliferation by modulating their cell cycle and also upregulates their production of nerve growth factor, hepatocyte growth factor, and stromal cell-derived factor-1
Third, BMSCs can be transplanted directly, intravenously, intraarterially, or intrathecally. Although direct (intracerebral) injection permits most efficient delivery of the donor cells to the damaged tissue, a less invasive procedure would be optimal. Intravenous or intrathecal transplantation is attractive because it is a noninvasive, safe technique for the host CNS, but has been reported to result in less pronounced cell migration and functional recovery than direct cell transplantation (Vaquero et al, 2006). Therefore, an optimal transplantation technique should be developed to serve maximally safe and efficient results. Alternatively, the intraarterial injection of BMSCs may be valuable to noninvasively deliver them to the damaged CNS (Osanai et al, 2011; Shen et al, 2006). There are a limited number of studies that directly compare the therapeutic effects of these delivery routes under the same conditions. It is urgent to test the effects of each delivery route on functional recovery after cerebral infarct (Kuroda et al, 2011). More interestingly, tissue-engineering technology may also provide an alternative route for cell delivery. Degradable biomaterials have been accepted as a valuable ‘scaffold’ to fix and stabilize the transplanted cells in other organs such as bone, cartilage, heart, and skin. Until recently, however, there have only been a small number of studies that denote effective scaffolds for cell transplantation for CNS disorders (Lu et al, 2007). A recent study has shown that a fibrin matrix may improve the survival and migration of BMSCs, being a useful material for injured CNS tissue (Yasuda et al, 2010). A thermoreversible gelation polymer hydrogel may also be one candidate scaffold to provide a suitable environment for BMSCs (Osanai et al, 2010). However, no clinical trials have been performed to examine the efficacy of such technology in cell therapy for ischemic stroke.
Finally, it would be essential to develop techniques to serially and noninvasively track the fate of the transplanted cells in the host CNS. A cell tracking technique would also be important as a ‘biologically relevant end point’ (Savitz and Fisher, 2007). Magnetic resonance imaging, nuclear imaging, and optical imaging are candidate modalities. Donor cells can be identified on magnetic resonance imaging by labeling with a superparamagnetic iron oxide agent (Hoehn et al, 2002; Ito et al, 2011; Jendelova et al, 2003). Magnetic resonance imaging can visualize intact, opaque organisms in three dimensions with good spatial resolution, but requires long imaging times and consequently slows data acquisition because of low sensitivity. Nuclear imaging can also detect the transplanted cells by labeling them with radioactive tracers. Correa et al (2007) recently labeled BM mononuclear cells with 99mTc-hexamethylpropylene (HMPAO), and injected them into a patient with ischemic stroke through a catheter. The transplanted cells were visualized on single photon emission tomography. Nuclear imaging can detect the target with high sensitivity, but has the difficulty to monitor donor cells for several weeks because of the relatively short half-life of clinically available tracers. Separately, optical imaging techniques may also serve future technology to visualize BMSCs engrafted in the damaged CNS. Quantum dot emits near-infrared fluorescence with a longer wavelength (800 nm) that can easily penetrate living tissue. A very recent study has shown that the quantum dot-labeled BMSCs can be clearly visualized under
In conclusion, recent studies have strongly suggested the therapeutic potential of BMSC transplantation for ischemic stroke. However, further translational studies would be warranted to establish it as a scientifically proven strategy in a clinical setting. In addition, a cell-based therapy combined with other procedures such as recanalization strategies would provide additional benefits for patients.
In this review, we summarize the current progress of basic stem cell science and its early clinical applications for advanced stem cell therapy, with a focus on iPS cells, G-CSF, and BMSCs as current topics. Problems such as tumorigenicity of iPS cells and exaggerated inflammatory response of G-CSF need to be overcome. The mechanisms underlying functional recovery after cell transplantation, including of BMSCs, remain to be clarified. Although it may take time to realize a future therapy for human stroke, the current prospect supported by successful research looks promising.
Footnotes
Disclosure/conflict of interest
The authors declare no conflict of interest.