
Abstract
The exact etiology of delayed cerebral vasospasm following cerebral hemorrhage is not clear, but a family of compounds termed ‘bilirubin oxidation end products (BOXes)’ derived from heme has been implicated. As proper regulation of vascular smooth muscle tone involves large-conductance Ca2+- and voltage-dependent Slo1 K+ (BK, maxiK, KCa1.1) channels, we examined whether BOXes altered functional properties of the channel. Electrophysiological measurements of Slo1 channels heterologously expressed in a human cell line and of native mouse BK channels in isolated cerebral myocytes showed that BOXes markedly diminished open probability. Biophysically, BOXes specifically stabilized the conformations of the channel with its ion conduction gate closed. The results of chemical amino-acid modifications and molecular mutagenesis together suggest that two specific lysine residues in the structural element linking the transmembrane ion-permeation domain to the carboxyl cytosolic domain of the Slo1 channel are critical in determining the sensitivity of the channel to BOXes. Inhibition of Slo1 BK channels by BOXes may contribute to the development of delayed cerebral vasospasm following brain hemorrhage.
Introduction
Large-conductance Ca2+- and voltage-dependent Slo1 K+ (BK, maxi-K, KCa1.1) channels open in response to an increase in intracellular Ca2+ concentration and/or depolarization, generally providing a negative feedback influence on cellular excitability (Salkoff et al, 2006). Physiologic roles of Slo1 BK channels are well documented in many phenomena, including regulation of smooth muscle tone, determination of action potential duration, action potential frequency adaptation, neurotransmitter release, and neurovascular coupling (reviewed in Salkoff et al (2006)). Mice with the gene for the Slo1 channel pore-forming subunit (
Some human diseases may be associated with the dysregulation of Slo1 BK channels. For instance, a mutation in one of the putative Ca2+-sensor regions of the Slo1 channel is linked to epilepsy and paroxysmal movement disorder (Du et al, 2005). A form of hypertension has been reported to involve downregulation of the vascular Slo1 BK channel complex (Amberg and Santana, 2003), and a gain-of-function mutation in the auxiliary subunit gene β1 may offer protection against diastolic hypertension (Fernández-Fernández et al, 2004). Furthermore, pharmacological openers of Slo1 BK channels have been suggested to have a neuroprotective role following stroke (Gribkoff et al, 2001).
Subarachnoid hemorrhage (SAH) is a serious condition that claims many lives, and even those patients who survive the initial incidence often face delayed high mortality and exhibit delayed neurologic deterioration (Macdonald et al, 2007). Subarachnoid hemorrhage is often followed by delayed cerebral vasospasm, long-lasting narrowing of cerebral blood vessels, typically occurring 1 week after the initial hemorrhage (Macdonald et al, 2007). Although a definitive single etiology has not been established, heme and heme breakdown products released from hematomas may have critical roles in the vasospasm (Macdonald and Weir, 1991). In particular, a class of heme breakdown products known as bilirubin oxidation end products (BOXes; Figure 1A), small pyrroline derivatives, has been implicated (Clark et al, 2002; Clark and Sharp, 2006; Clark et al, 2008; Kranc et al, 2000; Pyne-Geithman et al, 2005). Bilirubin oxidation end products are present in cerebral spinal fluids (CSFs) of cerebral hemorrhage patients with vasospasm at concentrations of up to 1 μmol/L in the extracellular compartment (Pyne-Geithman et al, 2005) and potentiate contraction of isolated porcine carotid arteries (Clark et al, 2002; Kranc et al, 2000; Pyne-Geithman et al, 2008). Therefore, the evidence supports the idea that BOXes are important in the development of delayed cerebral vasospasm; however, the effectors or molecular targets of BOXes are only beginning to be revealed. Thus far, protein kinase C and the small GTPase Rho A have been suggested as direct or indirect effectors of BOXes (Pyne-Geithman et al, 2008). It is not known whether BOXes affect other cell constituents.
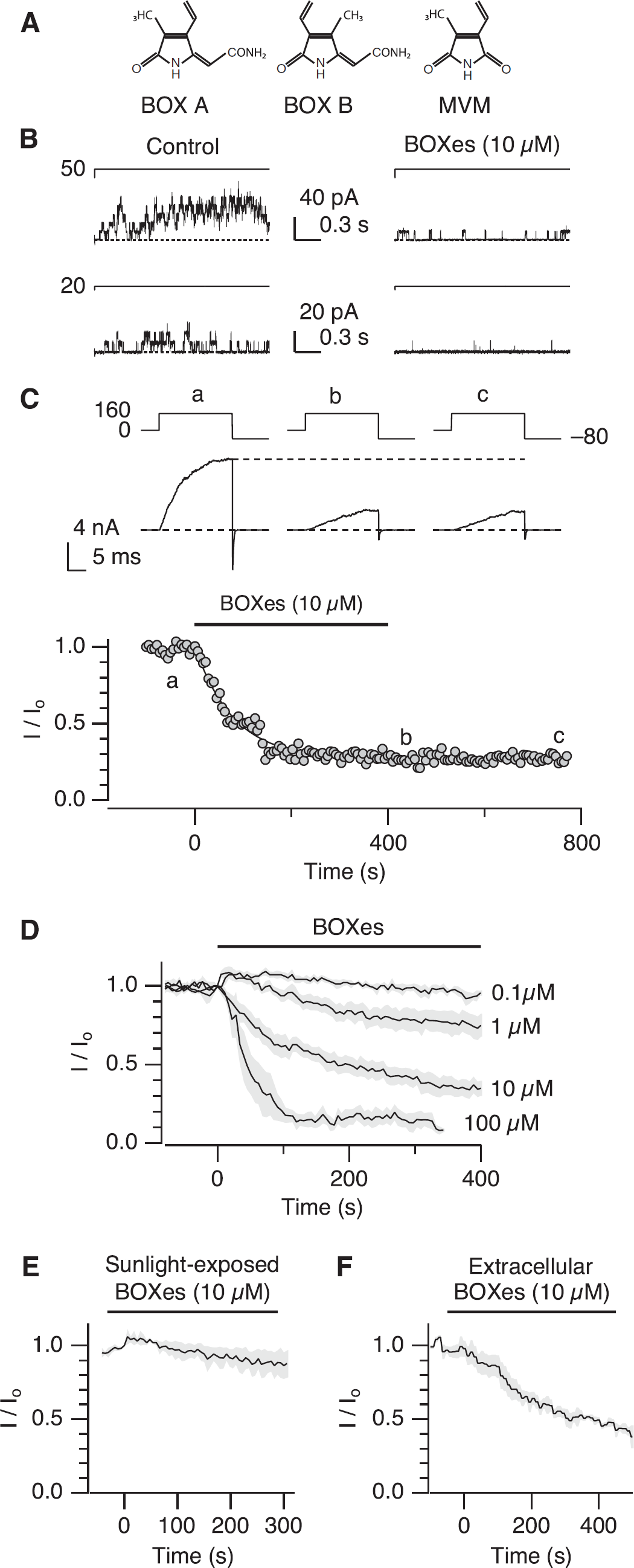
BOXes progressively inhibit Slo1 currents in a poorly reversible manner. (
The importance of Slo1 BK channels in the regulation of vascular smooth muscle tone (Nelson et al, 1995) and the vasoconstrictive ability of BOXes (Clark et al, 2002; Kranc et al, 2000; Pyne-Geithman et al, 2008) together raise the possibility that BOXes may regulate functional properties of Slo1 BK channels. If BOXes inhibit the activity of Slo1 BK channels in vascular smooth muscle cells, the inhibition could promote vascular constriction. In this study, we therefore examined whether BOXes alter functional properties of Slo1 BK channels using electrophysiological and molecular biological methods.
Materials and methods
Human large-conductance Ca2+- and voltage-gated Slo1 K+ channels (U11058/AAB65837) were heterologously expressed in human embryonic kidney (HEK) cells using FuGene 6 (Roche, Indianapolis, IN, USA) (Horrigan et al, 2005). In some experiments, Slo1 was cotransfected with mouse β1 (AAH13338) or human β1 (AAS20193) (1:1 weight ratio). Other constructs used are noted in the figure legends. Mutations were introduced to Slo1 using a standard PCR-based mutagenesis method (Agilent, Santa Clara, CA, USA) and the sequences were verified (University of Pennsylvania DNA Sequencing Facility, Philadelphia, PA, USA).
Basilar arteries and their main branches were collected from ∼2-month-old C57BL/6 mice using the protocol of Jaggar (2001), except that dithiothreitol replaced dithioerythritol. The animals were killed according to a protocol approved by the University of Pennsylvania Animal Care and Use Committee. Smooth muscle cells were identified by their elongated appearance.
Electrophysiological experiments were performed and the results were analyzed as described previously (Horrigan et al, 2005). The inside-out configuration of the patch-clamp method was used primarily to record ionic currents at room temperature using an Axopatch 200A or 200B amplifier (Molecular Devices, Sunnyvale, CA, USA). The output of the amplifier was digitized (ITC16, HEKA, Bellmore, NY, USA) and stored for later analysis. For macroscopic current measurements, the recording pipettes made of borosilicate glass (Warner, Hamden, CT, USA) had a typical resistance of 0.8 to 1 Mohms when filled with the solution described below and ≥ 50% of the initial pipette resistance was electronically compensated. Native BK channel currents were recorded with the extracellular solution containing (in mmol/L): 134 NaCl, 6 KCl, 1 MgCl2, 2 CaCl2, 10 glucose, 10 HEPES, pH 7.4 with NMG (
The electrophysiological results were analyzed using custom routines implemented in IgorPro (WaveMetrics, Lake Oswego, OR, USA). To generate macroscopic conductance-voltage (GV) curves, tail currents, excluding the initial ∼180 microseconds to avoid capacitative transients, at −80 mV following prepulses to different voltages were fitted with a single exponential function, and the extrapolated amplitudes at
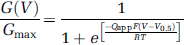
where
Single-channel open probability at negative voltages was estimated from all-point amplitude histograms using the half-amplitude criterion and corrected for the number of channels present in each patch. The number of channels was determined from the peak macroscopic current size recorded in the same patch at ≥160 mV before application of BOXes. The single-channel current–voltage curve (
The statistical results are presented as mean ± s.e.m. (
Bilirubin oxidation end products were synthesized and purified according to the protocol of Kranc et al (2000). Three members are present in the samples of BOXes used, namely BOX A, BOX B, and MVM (4-methyl-3-vinylmaleimide) (Kranc et al, 2000). Separation of BOXes to the individual components is difficult and drastically decreases the yield to <5% (Kranc et al, 2000), and the mixture containing all three components was used in the experiments presented herein because this is the ratio (2:2:1) found in CSF (BOX A, BOX B, and MVM, respectively). However, the preparations of BOXes were essentially free of bilirubin and other bilirubin derivatives. Bilirubin oxidation end products were applied with minimal illumination, typically to the cytoplasmic side and washed out. The effects of BOXes were slow to reverse (see below) and most of the results shown are comparisons of those obtained before and after treatment with BOXes followed by a washout. When the kinetics of the inhibitory effect of BOXes was examined, repeated pulses to a single voltage, at which the open probability was ∼0.5, were applied.
Simulations using the Horrigan and Aldrich (HA) model of gating of the Slo1 channel were performed as described previously (Horrigan et al, 2005). To simulate the results in the absence of Ca2+, the kinetic model (Figure 5A, bottom; Horrigan et al, 1999) was used to generate ionic currents with the parameter values shown in Table 1. The simulated ionic currents were then analyzed as described above to generate GV curves and the voltage dependence of the macroscopic time constant. The GV curves at different concentrations of Ca2+ were analytically simulated (Figure 5A, top) using a custom routine implemented in IgorPro (Horrigan et al, 2005).
HA model parameter values
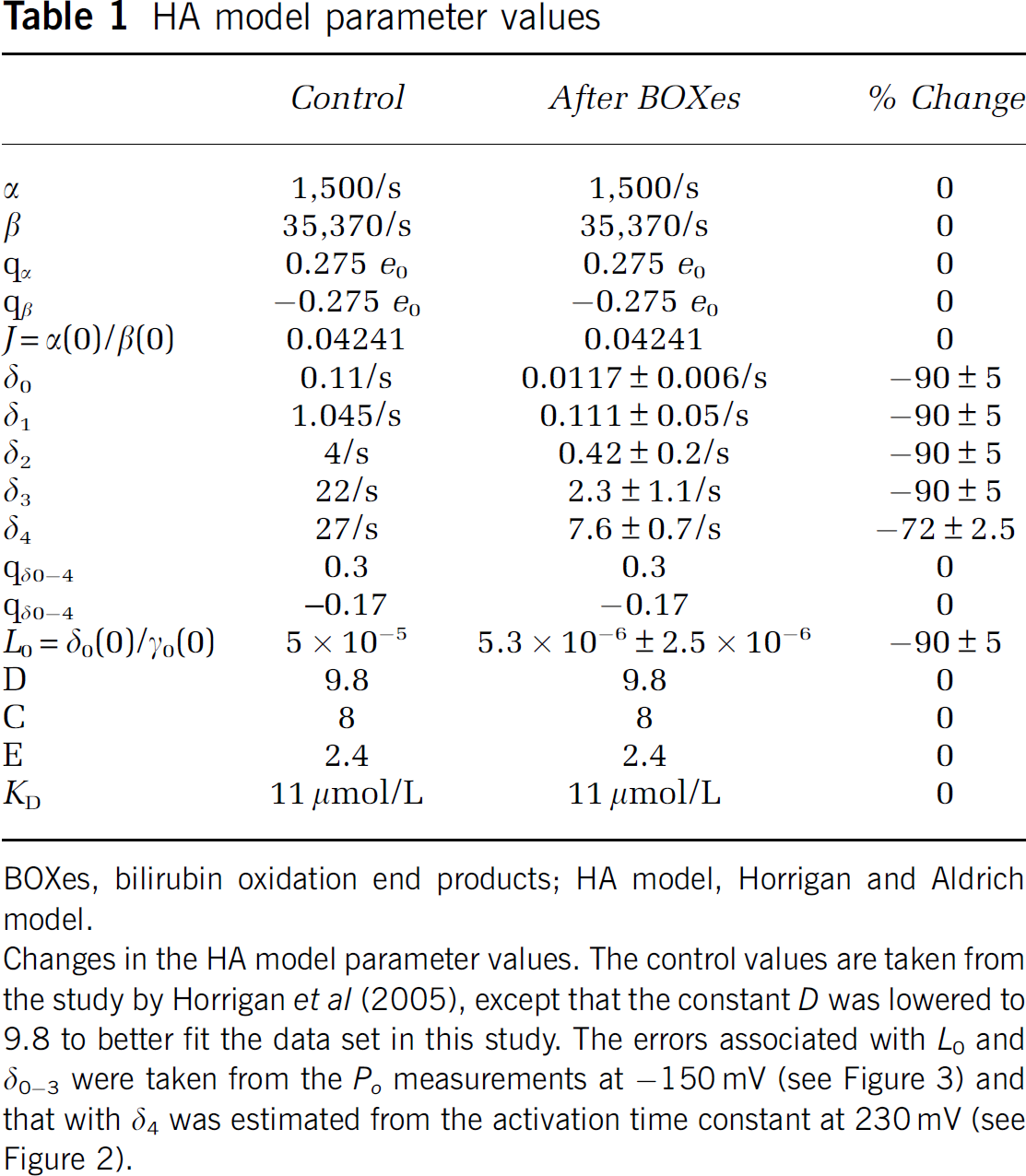
BOXes, bilirubin oxidation end products; HA model, Horrigan and Aldrich model.
Changes in the HA model parameter values. The control values are taken from the study by Horrigan et al (2005), except that the constant
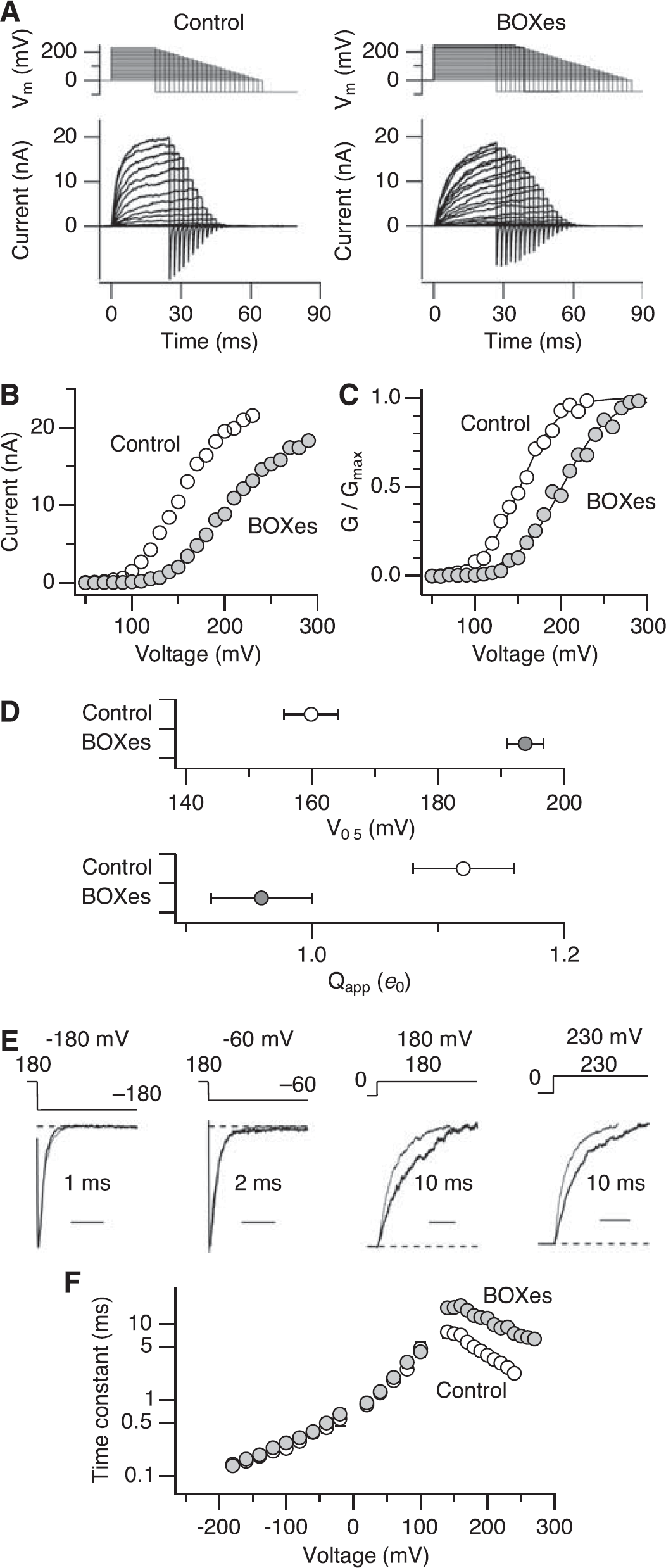
BOXes alter gating properties of Slo1 in the absence of Ca2+. (
BOXes decrease
Slo1 channels treated with BOXes remain Ca2+ sensitive. (
Simulations using the HA model. (
Chloramine-T, DEPC (diethyl pyrocarbonate), PGO (phenylglyoxal), dimethyl adipimidate (DMA), and iodoacetamide were obtained from Sigma (St Louis, MO, USA). Both MTSEA (2-aminoethyl methanethiosulfonate) and MTSET (2-(trimethylammonium) ethyl methanethiosulfonate) were acquired from Biotium (Hayward, CA, USA). The reagents were prepared immediately before use and applied to the intracellular side. The reagents were washed out of the chamber and the sensitivity to BOXes was examined.
Results
Application of BOXes (Figure 1A; 10 μmol/L) to the cytoplasmic side of a membrane patch excised from a mouse cerebral artery myocyte markedly decreased the frequency of Slo1 BK channel openings (Figure 1B). To elucidate the biophysical and molecular mechanisms of the inhibitory action of BOXes in detail, we heterologously expressed human Slo1 channels in HEK cells. The opening and the closing of the ion conduction gate of the Slo1 channel are allosterically controlled by its transmembrane voltage and cytoplasmic Ca2+ sensors (Figure 5A, top), and a full description of the voltage- and Ca2+-dependent gating of the channel requires a large number of conformational states (Horrigan and Aldrich, 2002). To simplify interpretations of the electrophysiological results, our initial experiments were performed in the virtual absence of Ca2+, such that the channel opening is allosterically controlled only by the voltage sensors (Horrigan et al, 1999; Figure 5A). As found with the native myocyte BK channels (Figure 1B), application of BOXes (10 μmol/L) to the cytoplasmic side of an excised cell-free membrane patch containing Slo1 channels progressively decreased the current elicited by depolarization roughly following an exponential time course with a time constant value of ∼80 seconds (Figure 1C). Extensive wash, up to ∼10 minutes, did not fully reverse the inhibition (Figure 1C), indicating a stable interaction between BOXes and the channel protein. Similar measurements in multiple experiments showed that the kinetics of inhibition became faster with greater concentrations of BOXes (Figure 1D). For ≥10 μmol/L, the steady-state fractional inhibitions observed within a typical lifetime of a patch (∼15 minutes) were similar. Lower concentrations of BOXes (0.1 and 1 μmol/L) appeared less effective (Figure 1D), but it was not clear whether the inhibition reached steady state within the limited measurement duration. The inhibition time courses with BOXes were indistinguishable whether the holding voltage was 0 or −100 mV (data not shown).
A previous study has identified that one of the notable characteristics of BOXes is that their vasoconstrictive ability is diminished by exposure to sunlight (Kranc et al, 2000). Consistent with this report, application of sunlight-exposed BOXes was much less effective in altering the current size (Figure 1E).
Application of BOXes to the extracellular side was also effective (Figure 1F), but the inhibition kinetics was often slower than that observed with application of BOXes to the intracellular side. The faster action of BOXes applied to the cytoplasmic side may signify that the sites on the channel required for the interaction with BOXes are more readily accessed from the intracellular side. The results presented in the remainder of this article were obtained using application of BOXes to the intracellular side using cell-free inside-out membrane patches. Bilirubin oxidation end products were typically applied at 10 μmol/L, because the inhibition kinetics with this concentration was rapid enough to obtain sufficient results using the patch-clamp method.
Single-channel current–voltage curves before and after treatment with BOXes (10 μmol/L for 10 minutes) were indistinguishable, showing that BOXes had no effect on the open-channel ion-conduction properties (Supplementary Figure 1). In contrast, voltage-dependent steady-state activation of the Slo1 channel was markedly altered by BOXes (Figure 2A). The Slo1 currents were inhibited more noticeably at less depolarized voltages (e.g., 120 versus 200mV in Figure 2B). The inhibition was accompanied by a ∼35 mV shift in half-activation voltage (V0.5) of macroscopic conductance (GV) to the positive direction (Figures 2C and 2D; 160 ± 4 mV versus 194 ± 3mV; P< 0.0001). The slope of the GV curve was also shallower; the apparent charge movement estimated from Boltzmann fits decreased by ∼15% from ∼1.10 ± 0.04 to 0.96 ± 0.04
Treatment with BOXes also altered the kinetics of the channel. Bilirubin oxidation end products noticeably slowed the activation kinetics at positive voltages at which the opening transitions dominate and open probability (
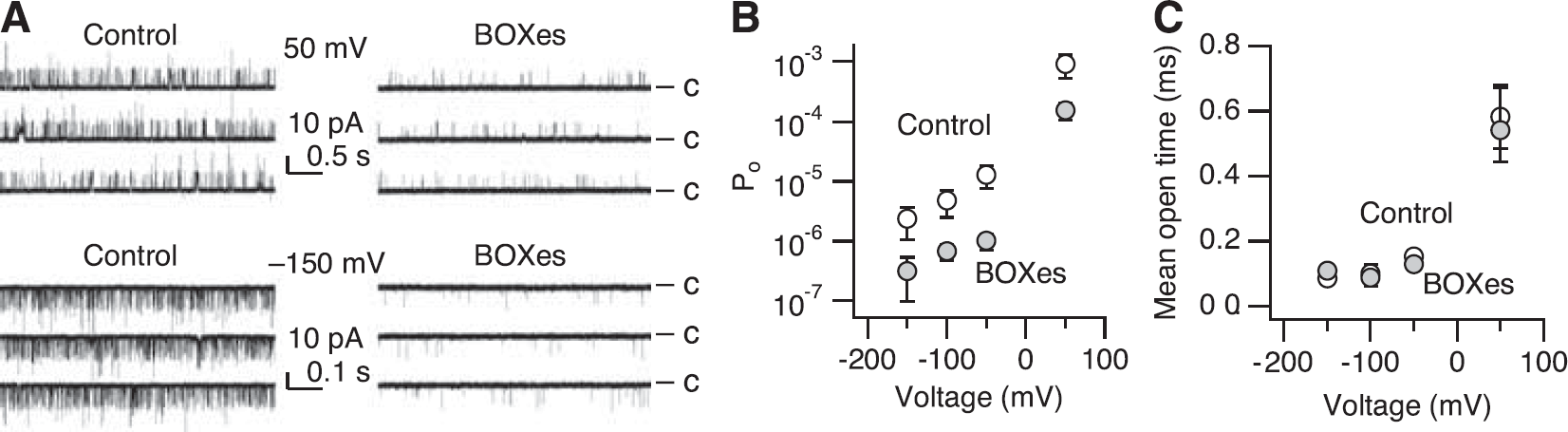
At extreme negative voltages (≤ −100 mV), the primary voltage sensors of the Slo1 channel largely assume the resting conformation (Figure 5A) (Horrigan et al, 1999). As voltage-sensor activation is not strictly required for the ion conduction gate of Slo1 to open, single-channel currents can be observed at such extreme negative voltages, although infrequently (Figures 3A and 5A) (Horrigan et al, 1999). Treatment with BOXes decreased
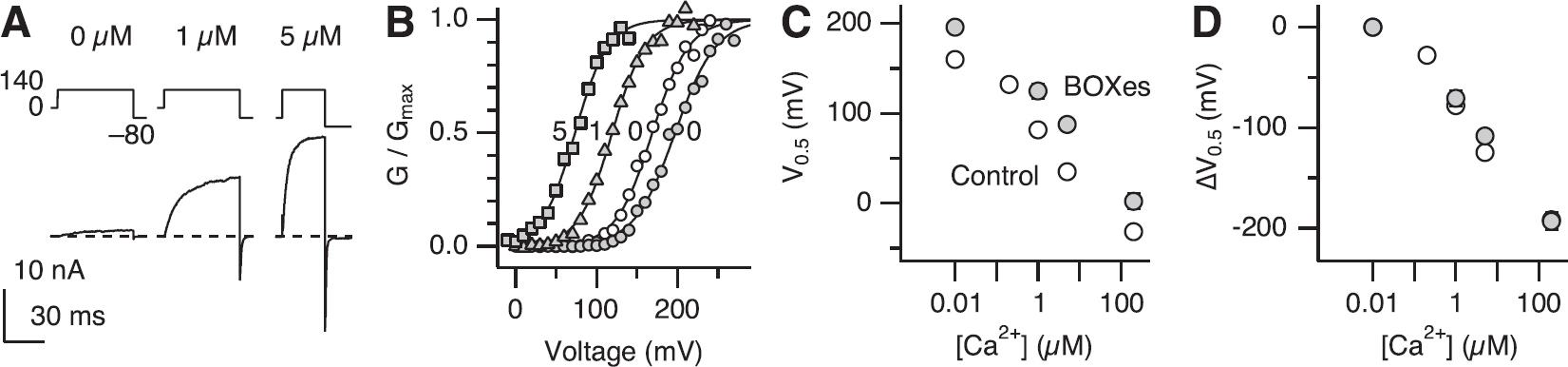
The results presented thus far show that treatment with BOXes leads to marked alterations in the functional properties of the Slo1 BK channels in the absence of Ca2+, reflecting the reciprocal and allosteric interactions between the gate and voltage sensors of the channel (Figure 5A). As physiologic activation of the Slo1 BK channel often involves intracellular Ca2+ (Nelson et al, 1995), we examined whether BOXes altered the Ca2+ sensitivity of the channel. After treatment with BOXes (10 μmol/L), Slo1 currents remained dependent on [Ca2+]i, increasing in size with higher concentrations (Figures 4A and 4B). At a given Ca2+ concentration, BOXes shifted the voltage dependence of activation as measured by V0.5 to the positive direction by 30 to 50 mV (Figure 4C). However, when normalized to the respective V0.5 values in the absence of Ca2+ (Figure 4D), the control channels and the channels treated with BOXes had similar Ca2+ dependence of activation. For example, in both groups, 1 μmol/L Ca2+ caused a −70 mV shift of
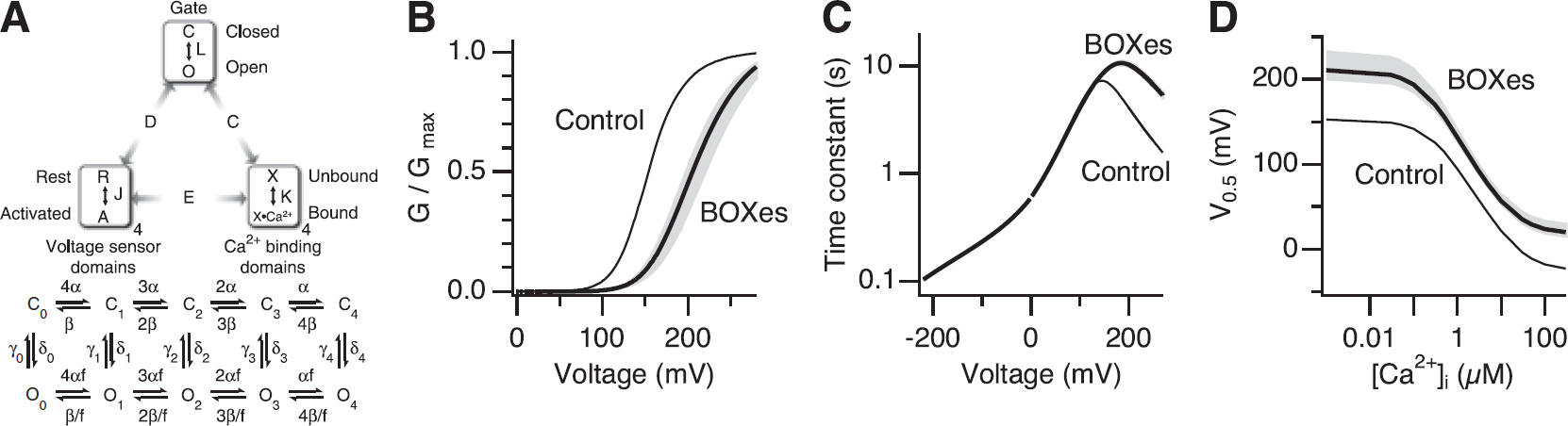
Steady-state activation properties of the Slo1 BK channel are described by the model of Horrigan and Aldrich (2002) (HA model), and the kinetic characteristics in the absence of Ca2+ are well approximated by a subset of the HA model (HCA model; Horrigan et al, 1999) as depicted in Figure 5A. To gain biophysical insight, the HA model was used to simulate the action of BOXes on the Slo1 channel gating by varying the model parameter values in the following manner. Without Ca2+, at extreme negative voltages at which voltage-sensor activation is negligible,
To explore which amino-acid residues of the Slo1 protein are required for the inhibitory effect of BOXes, we pretreated the channels with different amino-acid-modifying reagents and then, after washout, assessed the sensitivity of the modifier-treated channels to BOXes. Pretreatment with the methionine-preferring modifier chloramine-T, the imidazole modifier DEPC, the arginine modifier PGO, the cysteine modifiers iodoacetamide, MTSEA, and MTSET failed to alter the sensitivity of the channel to BOXes (data not shown). In contrast, pretreatment with the lysine modifier DMA (Duszyk et al, 1998) noticeably decreased the sensitivity to BOXes (Figures 6A–6D). The DMA pretreatment itself often decreased currents; however, the DMA-treated channels were clearly less sensitive to BOXes (Figures 6A versus 6B). Bilirubin oxidation end products (10 μmol/L for 400 seconds) decreased the current size only by 25% in DMA-treated channels compared with the ∼75% reduction observed in nontreated channels (Figures 6B and 6D).
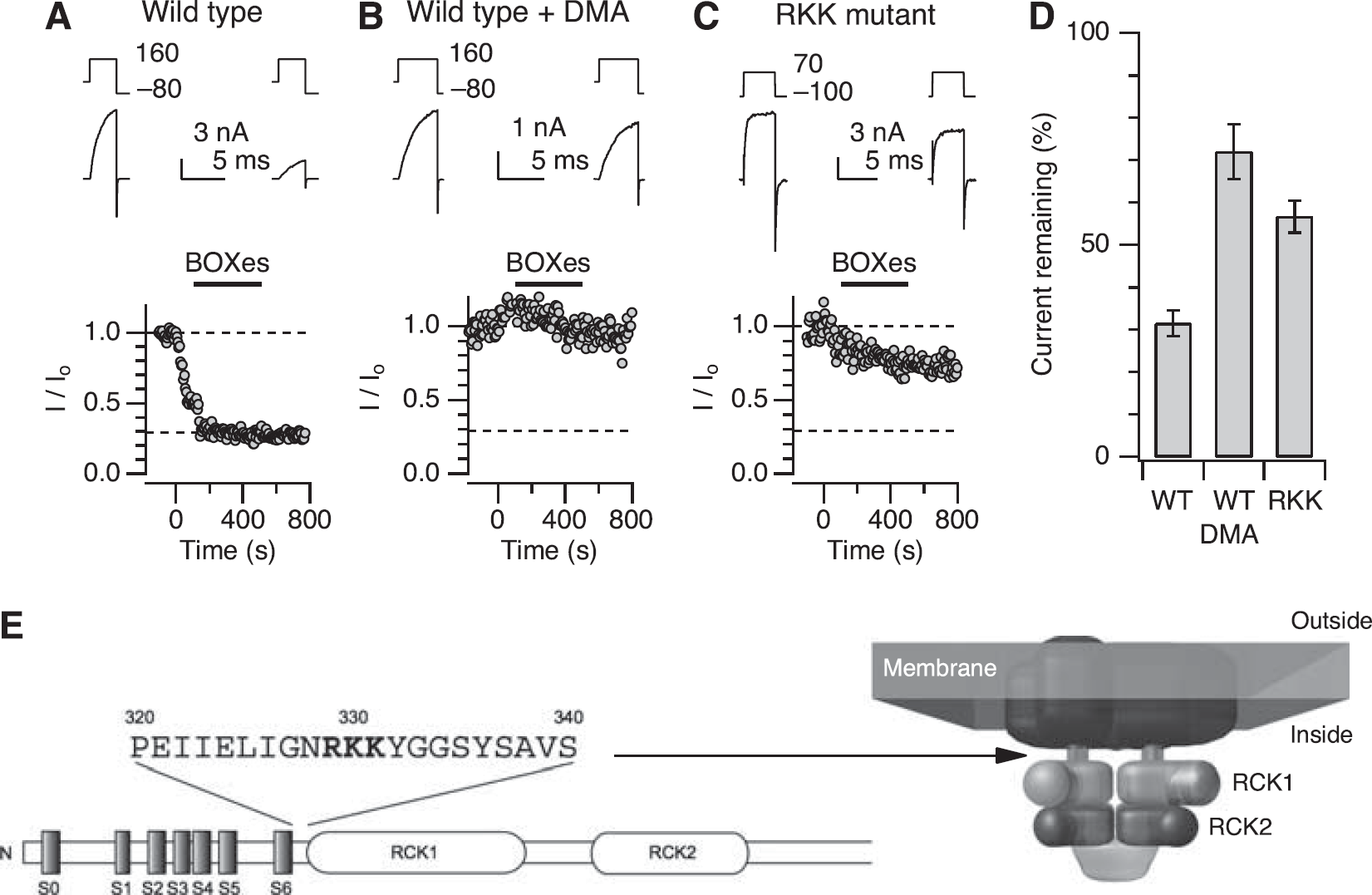
Lysine residues may be important in the action of BOXes. (
The ability of the lysine-modifying agent DMA to antagonize the inhibitory effect of BOXes suggests an involvement of Lys residues. Each Slo1 subunit (U11058/AAB65837) possesses 59 Lys residues and the majority of them are located in the large cytoplasmic C-terminal area. Of the numerous Lys residues, we focused on those in the sequence 329RKK331 in the linker segment between the transmembrane segment S6 and the cytoplasmic RCK1 domain (Figure 6E). This sequence has been implicated in the direct action of phosphatidylinositol 4,5-bisphosphate (PIP2); the triple mutation R329A:K330A:K331A (referred to as the RKK mutation hereafter) has been reported to diminish the stimulatory effect of PIP2 by 50% (Vaithianathan et al, 2008). Prompted by the purported importance of the 329RKK331 sequence, we tested whether the sensitivity of the channel to BOXes was also altered by the mutation. Indeed, the RKK mutation significantly diminished the sensitivity of the channel to BOXes (Figures 6C and 6D). After treatment with BOXes (10 μmol/L), ∼60% of the current remained in the RKK mutant compared with ∼30% remaining in the wild-type nontreated channel (
The transmembrane segments and the cytoplasmic RCK1 domains of the Slo1 channel together coordinate Mg2+ ions, which also promote the opening of the channel (Yang et al, 2008). As the sequence 329RKK331 is located in the S6-RCK1 linker segment (Figure 6E), we examined whether treatment with BOXes altered the Mg2+ sensitivity of the channel. Intracellular Mg2+ (10 μmol/L) produced a ∼100mV shift in V0.5 to the negative direction in channels treated with BOXes (Supplementary Figure 3), which was indistinguishable from that in the control group.
Native vascular BK channel complexes include both the pore-forming Slo1 and auxiliary β1 subunits (Salkoff et al, 2006). Consistent with the observation that BOXes inhibited native myocyte BK channels (Figure 1B), BOXes remained effective when Slo1 and β1 were heterologously coexpressed together (Supplementary Figure 3). In addition to Slo1 BK channels, vascular smooth muscle cells contain other K+ channels, including Kv1.5 (Brevnova et al, 2004). Bilirubin oxidation end products (10 μmol/L) did not alter currents through human Kv1.5 channels expressed in HEK cells (Supplementary Figure 4).
Discussion
Bilirubin oxidation end products, heme breakdown products, are found in the CSF of SAH patients and have been implicated in the development of delayed cerebral vasospasm following SAH (Clark and Sharp, 2006; Clark et al, 2008; Kranc et al, 2000). Although the molecular and cellular targets of BOXes are yet to be fully revealed, a recent study has shown that BOXes exert their action in part by regulating protein kinase C and GTPase Rho A (Pyne-Geithman et al, 2008). Our results here show that BOXes profoundly inhibit the Slo1 BK channel activity in a lysine-dependent manner and suggest that the inhibitory effect of BOXes on the channel could contribute to the vasoconstrictive effect of BOXes.
Gating of the Slo1 channel involves reciprocal and allosteric interactions among the ion conduction gate of the channel, transmembrane voltage sensors, and cytoplasmic ligand-binding domains (Horrigan et al, 1999; Horrigan and Aldrich, 2002). After treatment with BOXes, the voltage dependence of activation of the channel is shallower and shifted to the positive direction, and the activation kinetics is also slower. These changes in gating can be readily accounted for by postulating that BOXes specifically slow the opening process of the ion conduction gate of the channel. A 70% to 90% decrease in each of the four opening rate constants (δ0–4) in the HA model of Slo1 gating provides a good approximation of the channel's gating after exposure to BOXes.
The inhibitory effect of BOXes is somewhat reminiscent of that of heme, which also alters the Slo1 channel activity (Tang et al, 2003). Heme and its oxidized form hemin inhibit the channel at positive voltages in part by shifting the voltage dependence of activation to the positive direction (Tang et al, 2003). In addition, the activation kinetics is noticeably slower in the presence of heme/hemin (Horrigan et al, 2005). Despite the superficial resemblances, the underlying mechanisms of BOXes and hemin are markedly different. Bilirubin oxidation end products decrease the opening rate constant of the channel's gate (δ0) as shown in this study, whereas hemin increases the same rate constant (δ0) and weakens the allosteric coupling between the voltage sensors and the gate (D) (Horrigan et al, 2005) (see Figure 5A). The differential effects of hemin and BOXes are particularly evident at negative voltages at which the primary voltage sensors of the channel are at rest; hemin increases but BOXes decrease
The different biophysical mechanisms of the actions of hemin and BOXes on the Slo1 channel are reflected in the distinct amino-acid residues of Slo1 important for the actions of these related molecules. A short sequence containing two Cys and a His residue located in the cytoplasmic linker segment between the RCK1 and RCK2 domains of the Slo1 channel has an important role in the sensitivity of the channel to hemin (Jaggar et al, 2005; Tang et al, 2003). In contrast, the full sensitivity of the channel to BOXes requires the sequence 329RKK331 located downstream of the S6 transmembrane segment in the primary sequence.
The sequence 329RKK331 is also important in the direct action of the membrane-bound signaling molecule PIP2 on the channel; contemporaneous mutation of the three residues to alanine has been reported to diminish the effect of PIP2 by 50% (Vaithianathan et al, 2008). Given the putative location of the sequence 329RKK331 downstream of the S6 segment, it may be speculated that BOXes position themselves between the transmembrane helices and the cytoplasmic RCK1 domain, and stabilize the closed conformation of the ion conduction gate. The interaction of BOXes with the channel may be further stabilized by the probable ability of hydrophobic and planar BOXes to partition well in membranes and/or to intercalate nonpolar areas of the channel protein, as suggested by the finding that BOXes from either side of the membrane are effective. The relatively hydrophobic characteristic of BOXes should also contribute to the poor reversibility by simple washing. These membrane-permeable and light-sensitive characteristics of BOXes somewhat resemble those of the voltage-gated Ca2+ channel modulators dihydropyridines (Meyer et al, 1984). The diminished inhibitory effect of the BOXes following sunlight exposure is consistent with an earlier report (Kranc et al, 2000). Such light exposure has been postulated to photooxidize BOX A and BOX B to MVM (Clark and Sharp, 2006). Thus, the amide group, present in BOX A and BOX B but absent in MVM (Figure 1A), may be required to inhibit the Slo1 BK channel.
In addition to the sequence 329RKK331, there may be other determinants of the inhibitory effect of BOXes because the RKK mutation leaves ∼35% to 40% of the inhibition intact. Mechanistically, the sequence 329RKK331 in the S6-RCK1 linker could function as a binding site component. Alternatively, the RKK mutation may alter the accessibility of BOXes to the binding sites located elsewhere. Our results do not directly distinguish these two possibilities.
The exact structural location of the ion conduction gate in the Slo1 channel is yet to be revealed, and consequently, elucidation of the physicochemical basis of the preferential closed-state stabilization by BOXes requires further investigation. The linker connecting the S6 segment and the cytoplasmic RCK1 domain, which includes the sequence 329RKK331, has been envisioned to work like a passive mechanical spring to influence voltage-dependent gating of the Slo1 channel (Niu et al, 2004). However, it may be noted that our results do not show that BOXes alter the strongly voltage-dependent transitions mediated by the transmembrane voltage sensors.
Multiple types of K+ channels are expressed in vascular smooth muscle cells and alterations in any one of the channels could contribute to vascular dysfunction, including delayed cerebral vasospasm (Wellman, 2006). A transcript profiling study using a canine model of SAH involving autologous blood injection documented the downregulation of Kv2.2 and upregulation of Kir2.1 but no change in the expression of Slo1 or Cav1.2 (Aihara et al, 2003). Using the same canine model of SAH, Jahromi et al (2008) showed that the BK channel activity in acutely dissociated basilar artery smooth muscle cells is unaltered; however, the concentrations of BOXes were not measured in the study. How these findings using native cells harvested from the experimental SAH animals relate to our finding that BOXes regulate Slo1 is not clear. It is possible that the inhibitory effect of BOXes was somehow removed during the cell dissociation process, or that native BK channels possess or are exposed to cellular factors that protect them from BOXes or facilitate their unbinding.
SAH leads to the generation of multiple hemoglobin breakdown products, the concentrations of which in the CSF change with time. One of the early breakdown products is oxyhemoglobin, which has been suggested to have a role in SAH-induced cerebral vasospasm. The role played by oxyhemoglobin, however, may, some argue, be limited to early vasoconstrictive effects owing to its relatively short time in the hemolysate and also in part to its ability to scavenge the vasodilator nitric oxide (Windmuller et al, 2005). Thus oxyhemoglobin is an antidilatory agent as opposed to a procontractile agent (Aoki et al, 1994; Windmuller et al, 2005). The role of oxyhemoglobin in vasoconstriction during delayed vasospasm at 3 to 7 days is therefore unclear compared with other chemical species resulting from the hemorrhage. Bilirubin oxidation end products, in contrast, may serve as a procontractile agent (Pyne-Geithman et al, 2008), and the inhibitory effect of BOXes on the Slo1 channel may contribute to the development of delayed cerebral vasospasm following SAH.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.