
Abstract
Astrocytes are increasingly believed to play an important role in neurovascular coupling. Recent
Keywords
Introduction
Cerebral blood flow dynamics are functionally linked to neural activity in a process termed ‘functional hyperemia’ (Devor et al, 2003; Takano et al, 2006). Although the precise mechanisms underlying functional hyperemia remain uncertain, astrocytes have been implicated as important contributors (Takano et al, 2006; Winship et al, 2007). Astrocytes are ideally positioned to both detect neural activity and transmit signals to nearby blood vessels. They possess stellate or spongiform morphology (Hama et al, 1994), with many cellular processes extending from their soma that form tripartite synapses with neurons and interact with cerebral blood vessels (Koehler et al, 2009). Furthermore, astrocytes control the production and release of vasodilators, such as prostaglandins and epoxyeicosatrienoic acids, and the vasoconstrictor 20-HETE (Koehler et al, 2009; Haydon and Carmignoto, 2006). Recent studies have also shown changes in blood vessel diameter concurrent with Ca2+ signaling in astrocytes (Girouard et al, 2010; Filosa et al, 2006; Takano et al, 2006). In addition to producing vasoactive arachidonic acid metabolites, release of Ca2+ within astrocytes also triggers changes in smooth muscle cell K+ dynamics and vascular tone through the actions of Ca2+-sensitive (BK) and inward-rectifying (Kir) K+ channels (Filosa et al, 2006; Girouard et al, 2010). Together, these characteristics of astrocytes support their potential to contribute significantly to functional hyperemia.
Functional hyperemia manifests as a regional increase in blood flow that localizes to specific responding regions of the cortex. In the rodent somatosensory cortex, regions of the capillary bed exhibiting hyperemia can extend over distances exceeding 1 mm. Arterioles feeding the responding regions can dilate up to 2 mm away from the center of the responding region within less than a second after stimulation begins. After 2 to 4 seconds, or later cessation of stimulation, surface arterioles begin to constrict and blood flow reduces to baseline, sometimes with a short undershoot (Chen et al, 2010; Hillman et al, 2007). Despite evidence linking signaling in individual astrocytes to changes in adjacent blood vessels (Takano et al, 2006), there is as yet no model that conclusively explains how these discrete observations can account for the level of coordinated and rapid actuation required to orchestrate the complete hemodynamic response. A precise understanding of the ways in which astrocyte networks physically interact with cerebral blood vessels should provide an important foundation upon which to build models of functional hyperemia.
Previous studies examining the physical relationship between astrocytes and cerebral blood vessels have been predominantly carried out
This study quantitatively examines the interrelations between astrocytes and blood vessels in living, intact rodent brains using
Materials and methods
Animal Preparation
All procedures were approved by the Columbia University Institutional Animal Care and Use Committee. Five male C57BL/6J mice (JAX, Bar Harbor, ME, USA), two GFAP-GFP mice (from local colonies), and one endothelial-specific receptor tyrosine kinase (TIE2)-GFP mouse (JAX) weighing between 20 and 30 g were anesthetized with either 2%±0.5% isoflurane in a 3:1 air to oxygen mix or 100 mg/kg ketamine and 10 mg/kg xylazine and secured in a custom stereotaxic apparatus. A section of the skull over the somatosensory cortex measuring ∼3 × 4 mm2 was then removed, taking care to preserve the underlying dura mater. The meninges covering the IVth ventricle were punctured to relieve cerebrospinal fluid pressure. Sulforhodamine 101 (SR101; S-359, Invitrogen, Carlsbad, CA, USA) was prepared at a 0.2 mmol/L concentration in artificial cerebrospinal fluid (ACSF), and its fixable analog Texas red hydrazide (T-6256; Invitrogen) (Nimmerjahn et al, 2004) was prepared by first adding 80 μL/mg methanol to the dry hydrazide stock and then diluting to 0.2 mmol/L in ACSF. A reduced concentration of 0.04 mmol/L SR101 was used for GFP mice. These solutions were applied to the surface of the cortex just below the dura using a glass micropipette connected to a Picospritzer III microinjection system (Parker Instrumentation, Cleveland, OH, USA). Approximately 10 μL of the dye solution was also placed onto the dura and allowed to incubate for 30 minutes before washing with ACSF. Bulk loading of SR101 into the cortex was unnecessary because it distributes readily throughout astrocytes within the cortex through their gap junctions (Nimmerjahn et al, 2004). A glass coverslip with a drop of 0.5% agarose in artificial cerebrospinal fluid was then sealed in place over the craniotomy using dental acrylic (Henry Schein, Melville, NY, USA), thereby reducing brain motion and contamination. In wild-type mice, dextran-conjugated fluorescein (FD2000S, Sigma-Aldrich, St Louis, MO, USA, 5 mg in 0.1 mL saline) was administered by a tail vein injection to provide blood vessel contrast.
In Vivo Imaging
After craniotomy and dye application, mice were maintained on intraperitoneal ketamine anesthesia at a dose of 30 to 50 mg/kg per hour before imaging. A custom-built two-photon laser-scanning microscope (Radosevich et al, 2008) equipped with a Ti:Sapphire laser (Mai Tai XF, Spectra Physics, Mountain View, CA, USA), three emission channels (350 to 505 nm, 505 to 560 nm, and 560 to 650 nm), and a × 20 objective lens (XLUMPLanFl 20X 0.95W; Olympus, Tokyo, Japan) was then used to image the somatosensory cortex with excitation wavelengths between 800 and 850 nm at depths up to and exceeding 500 μm below the cortical surface.
Vessels were identified as arteries, veins, or capillaries based on morphology, flow speed, and the direction of blood flow. Arteries typically have fewer branches, more linear morphology, and higher flow speeds relative to veins. Serial stacks of 250 × 250 μm2 images (400 × 400 pixels at 400,000 Hz pixel rate) separated by 1 to 2 μm increments and running from the cortical surface to depths of ∼500 μm were acquired by vertically translating the system's objective (using a M-112.1DG stage; Physik Instrumente, Karlsruhe, Germany). These image stacks recorded the interactions between astrocytes and cerebral blood vessels at multiple cortical depths through layer IV, which terminates at ∼450 μm below the surface of the mouse somatosensory cortex (Altamura et al, 2007).
Immunohistochemistry
After
Data Analysis
Custom analysis software was developed in MATLAB (Mathworks, Sherborn, MA, USA) to allow us to precisely determine each astrocyte's 3D position, dimensions, depth, and vessel proximity, as well as blood vessel density as described below.
To calculate the locations of astrocytes within the cortex, a single point that lay midway between the upper and lower boundaries of the soma was chosen for each cell. The depth dependence of astrocyte density was determined by mapping the total number of these points representing astrocyte somata contained within consecutive, nonoverlapping 20-μm depth increments. Although astrocyte domains may occupy areas much larger than 20 μm, across the use of a single representative voxel for each soma allowed clean grouping of astrocyte cell bodies into nonoverlapping 20-μm segments without the danger of double counting any cells. The glia limitans superficialis, which spans from the surface of the brain to a depth of ∼10 to 15 μm, was excluded from our analyses because of high cell volume and poor contrast within this region.
Masks of blood vessels were generated using a thresholding system and growing algorithm to mark pixels of sufficient brightness along a manually selected seed vessel in the 3D image stack. Capillary density was then calculated by excluding large diving arterioles and venules (identified by their connectivity, diameter, and tortuosity), and then by calculating the total number of capillary mask pixels at 2-μm depth increments below the surface of the cortex. Every voxel in the 3D image stack was considered, and the depth of each vessel pixel was taken as the
The separation between astrocytes and their nearest capillary was calculated by sampling the pixels in expanding spheres around each cell. When a capillary pixel was detected, the vector linking this pixel to the astrocyte soma marker was obtained, and all pixels between these two points were examined. The distance between the capillary pixel and the most peripheral point of the astrocyte cell body closest to the capillary was used as our measure of the separation between the two.
The diameter of blood vessels and their perivascular astrocyte sheaths were manually measured for arteries, capillaries, and veins using the average of numerous sample points along the length of each vessel. Locations at which astrocyte processes connected with vessels and locations of perivascular astrocyte somata in direct contact with vessels were also manually recorded for each type of vessel. These data were then used to calculate the density of astrocyte process contacts along each type of vessel and the spacing of perivascular astrocytes. The density of contacts between astrocyte processes and blood vessels was calculated per unit of vessel length (total visible processes per 100 μm of vessel length) and as a function of vessel surface area (processes per 100 μm2 of blood vessel).
Trends in the data were quantified using a combination of Pearson's correlation and one-way ANOVA (analysis of variance) implemented in PASW (Predictive Analytics SoftWare, Somers, NY, USA). Error in all data presented was calculated as s.e.m.
Spectral Unmixing
For analysis of images obtained from transgenic GFP mice, it was necessary to exclude any potential overlap between the fluorescence emission of SR101 and GFP. Therefore, to generate the images in Figure 3, we used a spectral unmixing strategy as previously used in the studies by Radosevich et al (2008) and Tsai et al (2009). In brief, the pure red—green—blue emission spectral signatures of SR101 and GFP were identified by sampling regions of images that contained only one of these two contrasts (e.g., an astrocyte in a GFAP-GFP mouse before SR101 staining and an astrocyte in a wild-type mouse with only SR101 staining). This yielded a unique set of three emission values for each contrast species (SR101 and GFP) that could be used in a nonnegative least squares fit to all of the tricolor pixels in images acquired in mice in whom both forms of contrast were present. The results of this fit are two images that represent the ‘concentration’ of either SR101 or GFP within each pixel, based on the detected red—green—blue spectrum of that pixel. This approach allows quantitative analysis of any true spatial overlap between these two contrasts without the possible confound of spectral overlap.
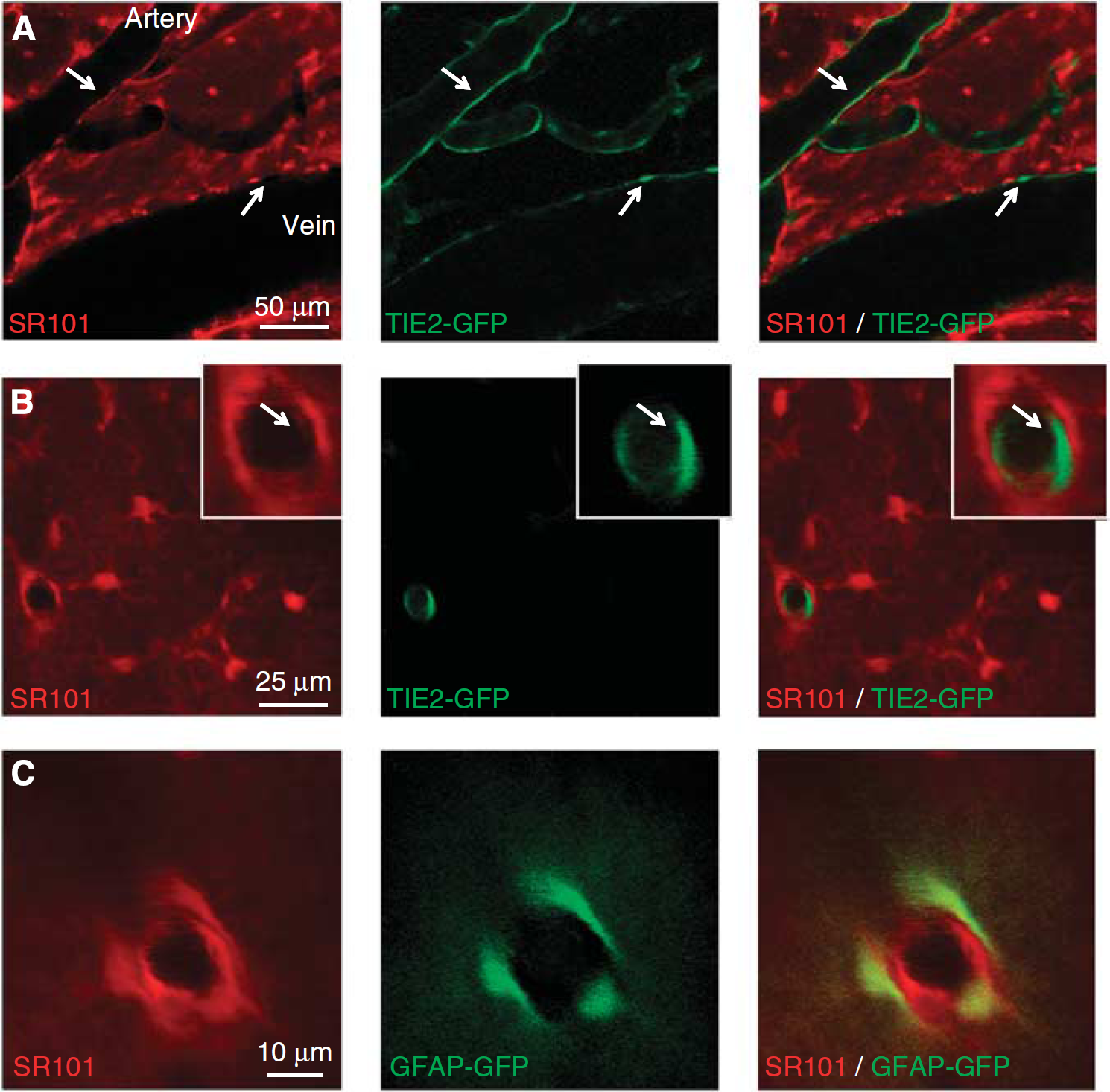
SR101 staining in transgenic TIE2-GFP and GFAP-GFP mice. (
Results
Astrocyte Distribution
On the basis of an examination of 4,167 cortical astrocytes in 8 image stacks (250 × 250 × 500 μm3 field of view) from 5 C57BL/6J mice, astrocyte density was found to vary in a consistent pattern as cortical depth increases (Figure 1A). Although the glia limitans (which was excluded from our automated analysis) has high astrocyte density, the region immediately below it from ∼10 to 40 μm deep contains very few astrocyte cell bodies. However, a dense network of astrocyte processes exists in this region. Astrocyte density (the number of astrocyte soma per unit volume) spikes sharply approaching 40 to 60 μm in depth before falling away by 180 μm. Below 180 μm, astrocyte density increases quite steadily as depth increases (Pearson's correlation between depth and astrocyte density below 180 μm is positive and significant:
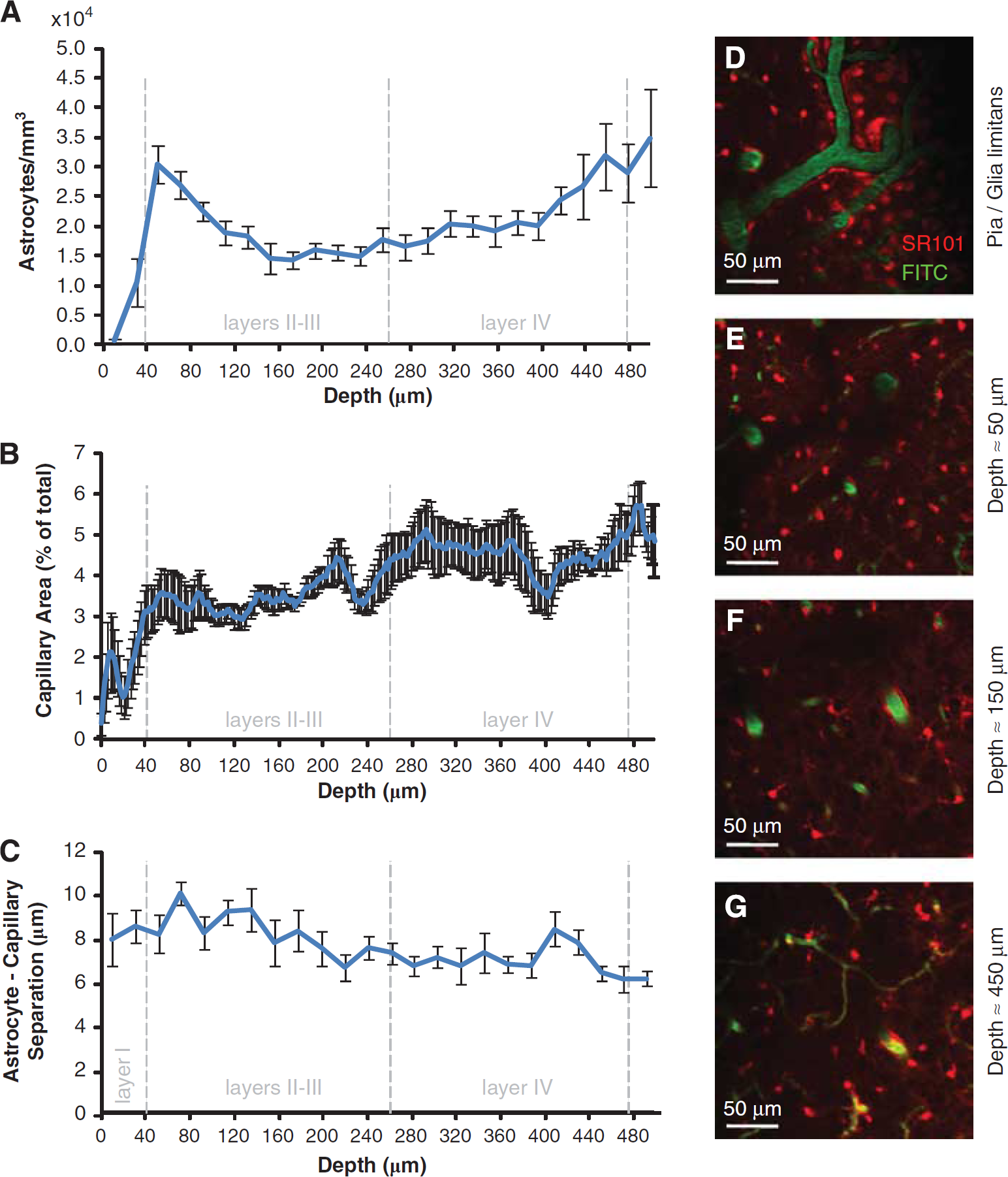
Astrocyte density, capillary density, and astrocyte—vessel separation. (
Capillary Density
Relatively few capillaries were found to occupy the region immediately below the glia limitans, but capillary density increases sharply as depth approaches 30 to 40 μm. Capillary density continues to increase at a reduced rate beyond 40 μm, eventually reaching an apparent maximum near 500 μm below the surface of the somatosensory cortex (Figure 1B) (Pearson's correlation between depth and capillary density is positive and significant:
Astrocyte—Capillary Separation
The separation between astrocytes and capillaries was found to slightly decrease, on average, as cortical depth increases (Pearson's correlation with depth is negative and significant:
Perivascular Sheathing Morphology
All blood vessels below the glia limitans were found to be surrounded by a perivascular sheath, brightly staining with SR101. Arteries on the pial surface, but never surface veins, were also found to exhibit robust perivascular SR101 staining (Figure 2A). For subpial veins, the strong SR101 sheath appears to merge with the glia limitans and disappear as the vein ascends to the pial surface (Figure 2B). A sheath can also be seen around capillaries, forming from an accumulation of processes from the adjacent astrocyte somata (Figure 2C).
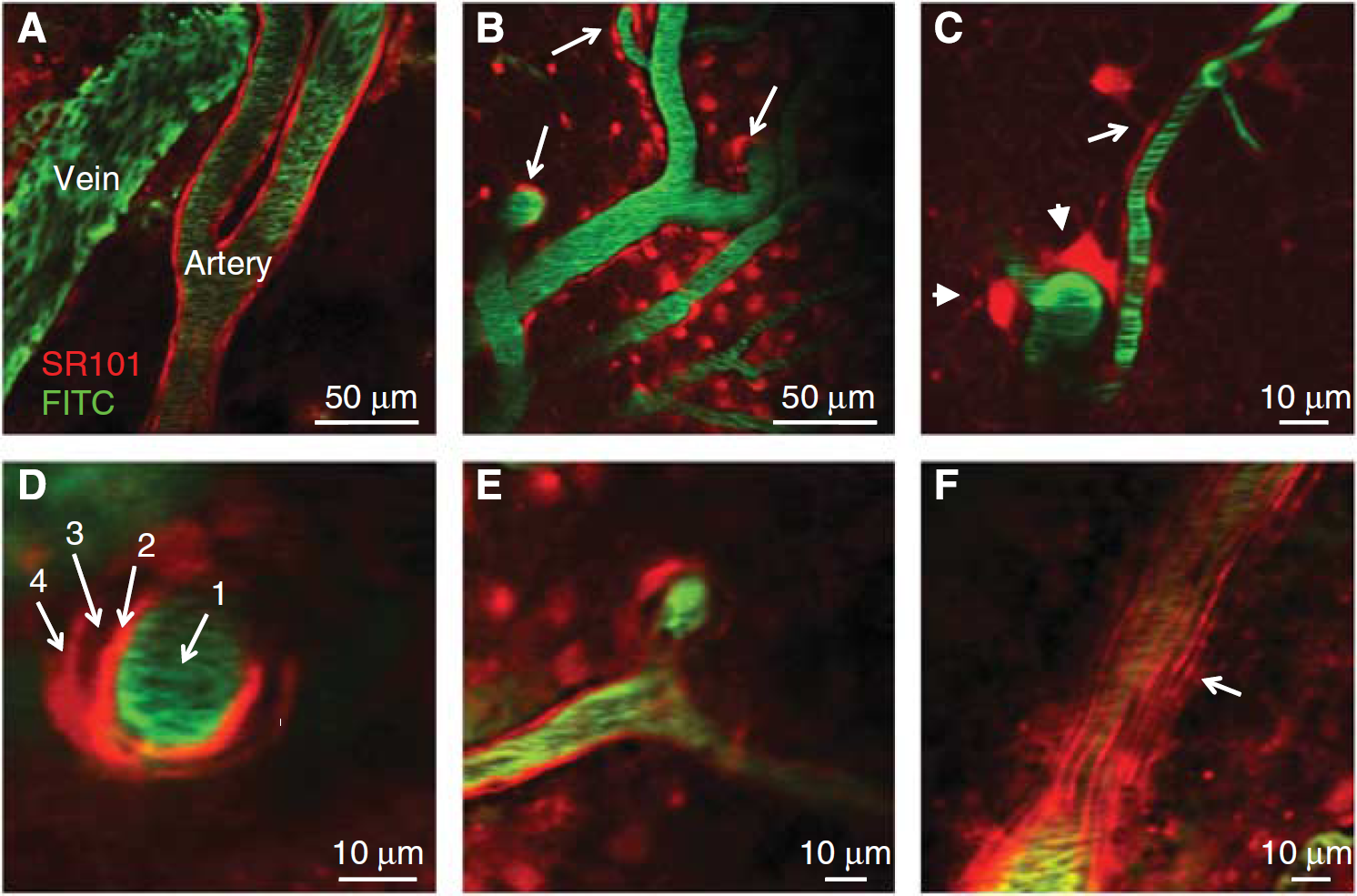
Perivascular sheathing morphology. (
A closer inspection of cortical arterioles and arteries revealed two distinct layers of SR101 sheathing (Figure 2D). Similar to ascending veins, diving arterioles possess an outer perivascular sheath that joins with the cells of the glia limitans and is no longer present above the pial surface (Figure 2E). However, the inner SR101-stained arteriolar sheath extends up through the glia limitans and along all pial arteries and arterioles. This inner arteriolar SR101 sheath exhibits consistent substructure appearing to be composed of elongated striations running parallel to the long axis of the vessel (Figure 2F). These inner structures were not found to be labeled in cortical veins either above or below the glia limitans.
Data acquired
Perivascular Sheath Thickness
Arteries were found to have the thickest astrocytic perivascular sheaths with an average of 2.6±0.30 μm. This number represents the thickness of the outer SR101 sheath around diving arterioles. Veins were found to have the second thickest sheath (2.1±0.05 μm), and capillaries had the thinnest astrocytic perivascular SR101 sheaths (1.2±0.04 μm). These differences are all significant (F(2, 24)=40.859;
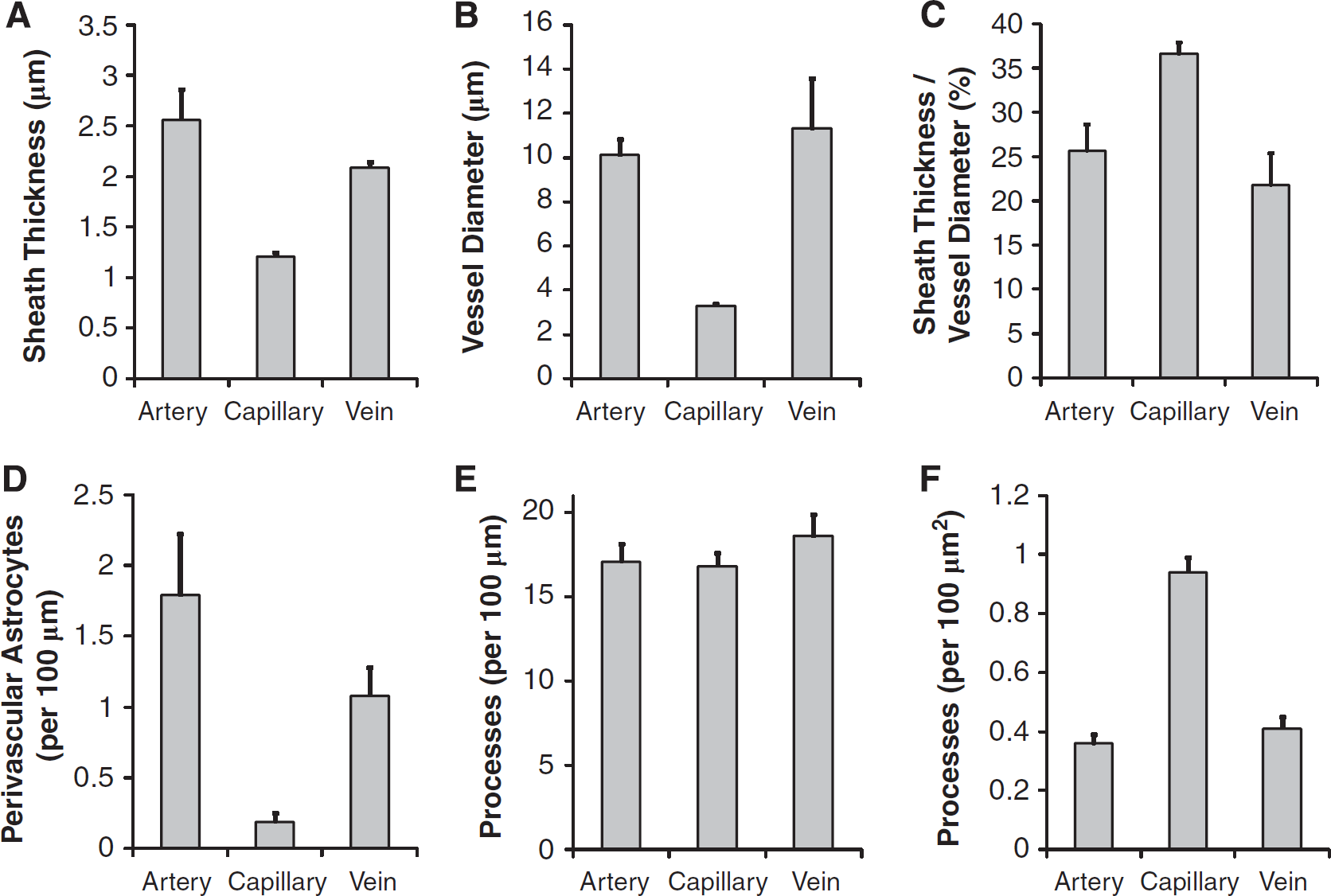
Measured astrocyte—vascular parameters. (
Perivascular astrocytes (astrocytes with somata physically contacting and arcing around a vessel wall) were more commonly found on arteries and veins compared with capillaries (F(2, 24)=20.091;
Astrocyte Process Density
On average, 17.07±1.05 astrocyte processes were found to contact a 100 μm segment of arteriole, 16.80±0.77 processes contact 100 μm of capillary, and 18.59±1.28 processes contact an equal length of venule. These differences are not statistically significant (F(2, 24)=0.848;
Immunohistochemistry Validation
In addition to using transgenic mice as controls, we also used IHC to verify the identity of structures stained by SR101. We found that although IHC GFAP labeling revealed astrocyte processes interacting with diving arterioles, staining did not persist above the glia limitans (Figure 5A), supporting findings from GFAP-GFP mice that the inner SR101 sheath of pial arteries is likely not due to the presence of astrocytes or their processes. Similarly, aquaporin-4 expression was observed around every vessel below the glia limitans, and robust staining was observed within the glia limitans itself (Figure 5B). However, perivascular aquaporin-4 staining was not observed on pial vessels and did not colocalize with the inner SR101 sheath of pial or diving arterioles.
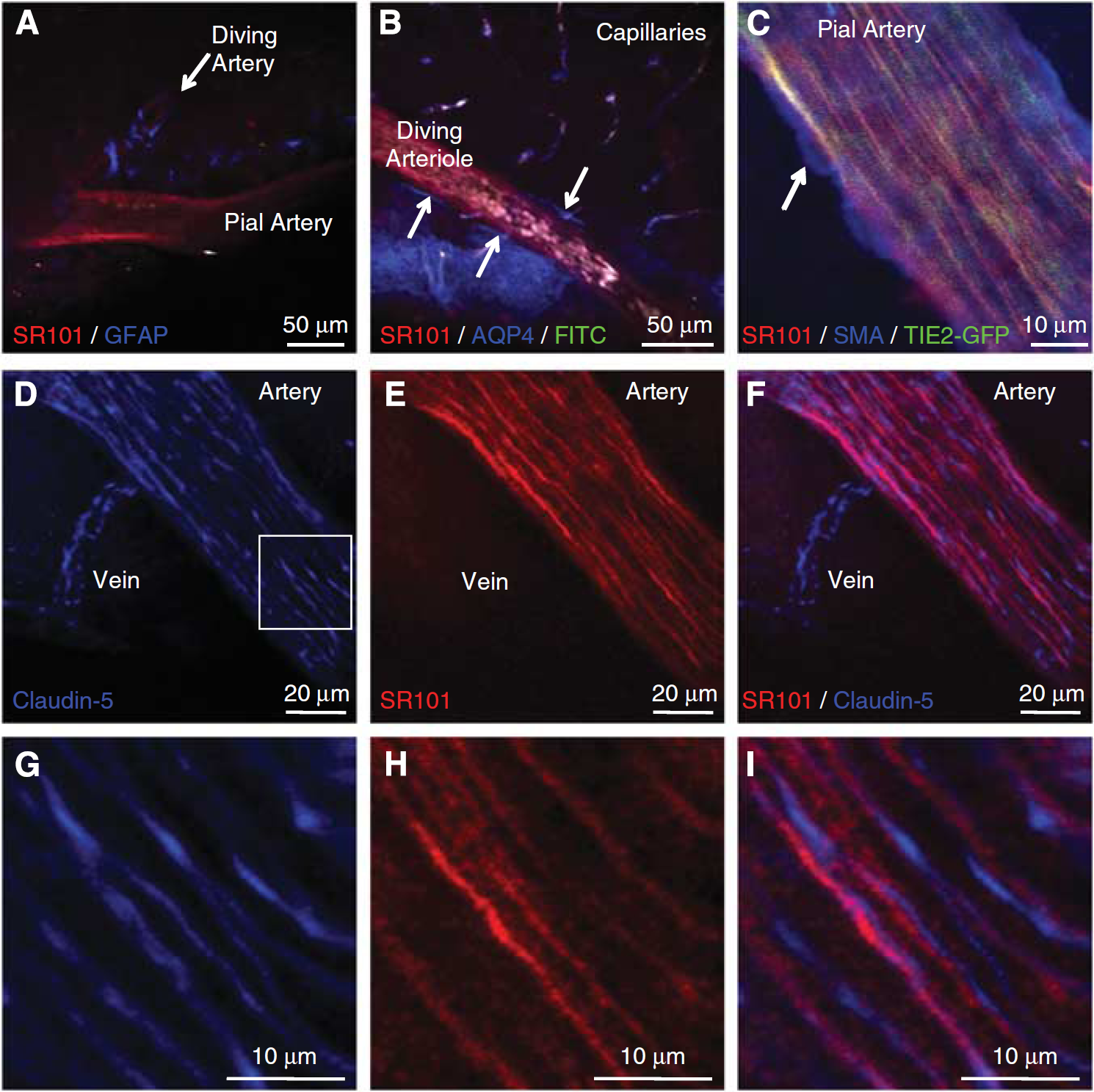
GFAP, AQP-4, SMA, and claudin-5
Conversely, IHC for the endothelial cell tight junction marker claudin-5 strongly costained with SR101 in the inner sheath of pial arteries. Furthermore, claudin-5 staining mimicked the fine structure of this SR101 sheath by staining elongated, parallel structures running along the long axis of the vessel (Figures 5D to 5I). Claudin-5 staining was also observed along pial veins, although the morphology of the stained structures was quite different from that seen on pial arterioles, being much less unidirectional (Figures 5D to 5F). No corresponding SR101 staining was seen on these surface veins. Immunohistochemistry for smooth muscle actin in TIE2-GFP mice further confirmed the location of the inner arteriolar SR101 sheath, showing that the arterial smooth muscle is present outside the colocalized inner SR101 sheath and endothelial GFP in pial arterioles (Figure 5C).
Discussion
The purpose of this study was to document the
Neurovascular Signaling: Beyond the Diving Arteriole
Our Results Suggest that Astrocytic Neurovascular Control May Not be Restricted to Diving Arterioles
It has been shown that astrocyte processes contacting diving arterioles exhibit increases in intracellular calcium [Ca2+] in response to glutamate and/or inositol triphosphate and may release vasoactive metabolites or alter perivascular K+ dynamics to induce changes in vascular tone (Girouard et al, 2010; Koehler et al, 2009; Iadecola and Nedergaard, 2007; Jakovcevic and Harder, 2007; Filosa et al, 2006). Neurovascular coupling literature to date has targeted diving arterioles as the likely point of control of functional hyperemia, assuming that changes in their diameter will have the highest impact on changing vascular resistance and therefore blood flow (Iadecola and Nedergaard, 2007; Nishimura et al, 2007; Takano et al, 2006). This, combined with the presence of smooth muscle, makes arterioles an obvious place to look when examining regulation of blood flow by astrocytes.
However, our findings show that every capillary and vein below the pia exhibits the same general morphology of interaction with astrocyte processes as diving arterioles. Given this lack of selectivity, we posit that neurovascular interactions involving astrocytes might equally occur at other locations, beyond diving arterioles. For example, if astrocytes are assumed to only sense and act discretely within their immediate vicinity, then these interactions would most likely occur in the capillary beds, since this is where most nutrient exchange occurs, and is therefore where increased neuronal activity or energy demands could be most rapidly sensed (Borowsky and Collins, 1989).
Studies focusing on diving arterioles also do not address the fact that the hemodynamic response manifests as a sequence of coordinated, rapidly propagated vasodilations and vasoconstrictions across several millimeters of the cortex (Hillman et al, 2007, Chen et al, 2010). If astrocytes do not simply act locally, but are exclusively responsible for modulating vascular tone, then they would need to be able to communicate and act over long distances in a coordinated manner. We discuss these two possibilities with regard to our findings below.
An Astrocyte Network for Neurovascular Signaling?
The Ubiquitous Presence of an Astrocytic Sheath Around All Subpial Vessels May Indicate that Astrocytes Can Indeed Signal Over Long Distances
Previous experiments using dye filling and transgenic mice have shown that processes from astrocyte cell bodies occupy discrete regions in space that only minimally overlap with adjacent astrocytes (Halassa et al, 2007; Wilhelmsson et al, 2006; Bushong et al, 2002). This discretization of astrocyte domains implies that astrocytes are unlikely to individually exert influences over distances of >100 to 200 μm. However, a recent study showed that gap junctions are highly expressed within perivascular astrocyte sheaths (Giaume et al, 2010), connecting the cytosols of adjacent astrocyte processes. Furthermore, astrocytes have been shown to be capable of communicating through Ca2+ signaling, and increases in astrocytic Ca2+ levels have in turn been linked to modulations in vascular tone (Takano et al, 2006; Zonta et al, 2003). Therefore, the comprehensive sheathing that we have observed along all subpial vessels could feasibly form a system of interconnected conduits capable of transmitting signals along the vasculature over distances that far exceed the discrete range of a single astrocyte.
We also observed that SR101 consistently labels the walls of pial surface arteries but not pial surface veins (Figures 2A and 3A), and that this staining did not correspond to the presence of astrocytes (Figures 3C, 5A, and 5B). Instead, our results suggest that SR101 is staining structures within the arterial endothelium (Figures 3A, 3B, and 5D to 5I). No corresponding SR101 labeling was seen on pial veins, suggesting that it is not simply the presence of endothelial cells or tight junctions that is causing the SR101 staining, but rather some difference between arterial and venous endothelial cells that causes the uptake of SR101 in arteries but not veins.
Nimmerjahn et al (2004) showed that SR101 is efficiently distributed to astrocytes deep within the cortex through their gap junctions (because this distribution was inhibited by gap junction/hemichannel blocker carbenoxolone, Cbx). Therefore, one explanation for the specific staining of the inner arteriolar SR101 sheath may be that astrocytes share gap junctions with the arteriolar endothelium. The possibility of a connection between the subpial astrocyte network and the endothelium of far-reaching pial arterioles is particularly compelling because it might suggest that vasomodulatory signals of astrocyte origin could be directly relayed over the long distances necessary for orchestration of the hemodynamic response to functional stimulus (consistent with the findings of Xu et al (2008)). If SR101 staining of the inner arteriolar SR101 sheath is not an indication of a functional connection between astrocytes and the arteriolar endothelium, then it is still evidence that the arteriolar endothelium possesses unique properties compared with capillary or venous endothelium, allowing it to uptake a substance that otherwise appears to be highly specific only to astrocytes.
Previously proposed mechanisms for the propagation of vasodilation have included possible contributions of endothelial cells (Andresen et al, 2006; Girouard and Iadecola, 2006), vasoactive interneurons (Cauli et al, 2004; Fahrenkrug et al, 2000), and smooth muscle cells (Girouard and Iadecola, 2006). If the network of astrocyte connectivity that we have observed has a role in this propagation, the next question that must be addressed is whether the speed of Ca2+ propagation between interconnected astrocytes could be sufficient to account for the rapid evolution of the hemodynamic response observed
Do Astrocytes Interact Preferentially with Capillaries?
Capillary—Astrocyte Proximity Suggests a Preferential Relationship Between Them that Overrides Spatial Variations in Microvascular and Astrocyte Density
Whether astrocytes propagate vasomodulatory signals directly or act via other intermediate cell types, the potential role of astrocytes as local sensors and even actuators remains. However, particularly if astrocytes are only able to act locally, their proximity to vessels and their relative densities might be important indicators of their role in neurovascular coupling.
Our
If astrocytes are predominantly acting locally, this also means that spatial variations in their density are noteworthy. For example, the peak in astrocyte density at ∼60 μm (Figure 1A) may indicate that this region has particular significance for neurovascular control. Several studies have attempted to explore the depth dependence of blood flow changes in the cortex (Nielsen and Lauritzen, 2001, Hillman et al, 2007); however, none to date have had sufficient resolution to determine whether this astrocyte-dense layer might be of particular functional importance.
What Might the Role of Astrocytes at the Capillary Level Be?
Are Astrocytes Involved in Hemodynamic Modulation of Capillaries, or is Their Presence Primarily for Nutrient Exchange and/or Neurotransmitter Scavenge?
Astrocytes have been hypothesized to serve many different roles in the brain. Therefore, the physical linkage between astrocytes and capillaries can be interpreted in a number of ways. For example, the majority of oxygen, nutrient, and waste exchange occurs at the capillary level, and therefore astrocytes might simply be positioned to ensure that they have adequate access to this supply (Borowsky and Collins, 1989). As neurons do not appear to exhibit similar perivascular morphology to astrocytes (Cauli et al, 2004), close connectivity between astrocytes and capillaries may support hypotheses that astrocytes have vital roles as intermediaries in providing nutrients, such as lactate to active neurons (Giaume et al, 2010; Rouach et al, 2008; Pellerin et al, 1998).
However, if astrocytes' roles extend beyond nutrient and waste exchange, their physical interrelation with capillaries may suggest that they sense local increases in activity and metabolic demand (Pellerin et al, 1998) to mediate changes in vascular tone (either by propagation through the perivascular astrocyte sheath network described above or as intermediaries with other cell types). A further possibility is that the physical connectivity between astrocytes and capillaries may actually form part of a mechanism capable of actively mediating changes in capillary tone or even permeability.
Recent studies have suggested that initiation of the hemodynamic response manifests as a rapid increase in tissue hemoglobin concentration in the capillary bed, with subsequent upstream propagation of dilation along the arteriolar tree (Chen et al, 2010; Sirotin et al, 2009). To account for these observations, an active mechanism around the level of the capillary bed (including precapillary arterioles or postcapillary venules) must be capable of inducing a rapid increase in the number of red blood cells in the capillary bed before upstream arteriolar dilation. Mechanisms by which astrocytes might modulate capillary hemoglobin concentrations in the absence of smooth muscle cells are uncertain (Hutchinson et al, 2006). One possibility is through interactions with pericytes, which are contractile cells located along central nervous system capillaries (Peppiatt et al, 2006). An alternative possibility might be that Ca2+-sensitive K+ (BK) channels on astrocyte endfeet (Filosa et al, 2006; Girouard et al, 2010) could vary intravascular hematocrit levels by altering the osmotic properties of the capillary walls.
Methodological Conclusions
We have shown that two-photon microscopy can be used to perform ‘
Summary
In summary, although control of functional hyperemia most likely involves the interactions of many different cell and vessel types, our findings have provided greater insights into the potential role of astrocytes in neurovascular coupling. Where astrocyte domains were previously believed to be small and discrete, we have found compelling evidence that astrocyte processes form conduits of connectivity that trace along all vessels within the cortex, potentially forming an ‘astrocytic superhighway’ capable of rapidly distributing vasomodulatory signals. We did not find strong evidence that astrocytes have preferential associations with diving arterioles, In fact, our results suggest that the capillary beds may be a more important site for astrocyte—vascular interactions. Our studies also present many questions, including whether the presence of a sheath on ascending veins obviates the significance of sheathing on arterioles and capillaries, and whether astrocyte signal propagation could be fast enough to account for observations of the hemodynamic response.
Footnotes
Acknowledgements
The authors thank James Goldman, Peter Canoll, and Edith Hamel for guidance and useful discussions, and Fiona Doetsch for generous donation of GFAP-GFP mice.
The authors declare no conflict of interest.