
Abstract
Disruption of the blood-brain barrier (BBB) caused by cerebral ischemia can initiate the development and progression of brain injuries, which may lead to irreversible dysfunction of the central nervous system. It is likely that tyrosine phosphorylation of a membrane-associated tight junctional protein, occludin, is important for the interaction of occludin with intracellular proteins, ZO-1 to ZO-3, and it regulates vascular permeability. Little is known about the pathophysiological alterations of tight junctional proteins after transient focal cerebral ischemia. In this study, we examined the tyrosine phosphorylation of occludin in isolated brain capillaries after transient focal cerebral ischemia. We further examined the effects of the Src-family tyrosine kinase inhibitor, PP2, on the tyrosine phosphorylation of occludin and on vascular permeability and infarct volume. Transient focal ischemia increased the tyrosine phosphorylation of occludin in the isolated brain capillaries. The administration of PP2 attenuated this phosphorylation, which was coincident with an inhibition of BBB leakage and a decrease in infarct volume. These results suggest that the increase in the tyrosine phosphorylation of occludin in the brain capillaries may be linked to the disruption of tight junctions, whose disruption can cause dysfunction of the BBB and the consequent increase in infarct volume.
Introduction
The blood-brain barrier (BBB) is composed of a continuous layer of brain capillary endothelial cells that are connected at a junctional complex by tight junctions and adherens junctions (Hawkins and Davis, 2005). Tight junctions form a metabolic and physical barrier that restricts the transport of macro-molecules between the blood and the brain for maintenance of cerebral homeostasis (Mitic and Anderson, 1998). The components of the tight junctions at the BBB include membrane-associated accessory proteins (such as occludin and claudin) and intracellular tight junctional accessory proteins (such as zonula occludens (ZO)-1 to ZO-3) (Hawkins and Davis, 2005).
The intracellular C-terminal domain of occludin is associated with the cytoskeleton through accessory proteins ZO-1, ZO-2, and ZO-3 (Furuse et al, 1994), indicating that this association is critical for maintenance of the BBB function. It has been shown that occludin has multiple sites for phosphorylation of its serine and threonine residues, and that the phosphorylation of these residues is involved in the regulation of its association with the cell membrane (Hawkins and Davis, 2005). In addition, the assembly of tight junctions can be regulated by nonreceptor tyrosine kinases in epithelial monolayers (Meyer et al, 2001; Rao et al, 1999, 2002). In this sense, it was shown that occludin appears to be phosphorylated on its tyrosine residues by Src-family tyrosine kinase (Chen et al, 2002). This phosphorylation site at the C terminus is critically important for the interaction of occludin with ZO-1, ZO-2, and ZO-3 molecules (Kale et al, 2003). These findings imply that protein tyrosine phosphorylation at tight junctions plays a crucial role in the regulation of BBB function.
An increase in vascular permeability caused by a variety of brain injuries including cerebral ischemia is a consequence of pathology and can induce brain edema (Rosenberg, 1999). Indeed, it is well recognized that ischemic cerebral edema is a common complication in stroke patients (Katzman et al, 1977; Ng and Nimmannitya, 1970). As disturbance of the BBB can be the initiating factor for the development of cerebral infarction, maintenance of BBB function at the acute stage after cerebral ischemia may have an important influence on the severity of the disease. Although thrombolysis with tissue plasminogen activator is widely accepted to be an effective therapy for acute thromboembolic stroke, symptomatic intracerebral hemorrhage associated with a delayed administration of it remains a major complication. In animal models, tissue plasminogen activator promotes the disruption of the BBB, which leads to edema and cerebral hemorrhage (Aoki et al, 2002; Tsuji et al, 2005; Yepes et al, 2003). Therefore, poor outcomes after tissue plasminogen activator administration may be associated with opening of the BBB.
Although the molecular structure of the tight junction has been identified, pathophysiological alterations in these proteins under an in vivo ischemic insult are not fully understood. Therefore, we sought to determine the role of tyrosine phosphorylation of occludin in the pathophysiological modification of the properties of the BBB and subsequent brain injuries after transient focal cerebral ischemia. Although we showed earlier that cerebral ischemia resulted in tyrosine phosphorylation of occludin and BBB dysfunction (Kago et al, 2006), it remains unclear whether such phosphorylation in the brain capillaries is caused by Src-family tyrosine kinases. In this study, we focused on the in vivo effects of the Src-family tyrosine kinase inhibitor, PP2, on tyrosine phosphorylation of occludin and also investigated the effects of PP2 treatment on vascular permeability and the development of cerebral infarction after transient focal cerebral ischemia.
Materials and methods
Animal Surgical Procedures
Male Sprague-Dawley rats (7 weeks, weighing between 240 and 280 g, Charles River Japan, Atsugi, Japan) were used in this study. The animals had free access to food and water, and were maintained according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals and the Guideline of Experimental Animal Care issued by the Prime Minister's Office of Japan. The study protocol was approved by the Committee of Animal Care and Welfare of Tokyo University of Pharmacy and Life Sciences.
Transient focal ischemia was induced using the method described earlier (Longa et al, 1989). Anesthesia was induced with 3% enflurane and maintained with 1.5% enflurane in a mixture of oxygen/nitrous oxide (30:70%). Rectal temperature was monitored and maintained at 37.0°C ± 0.5°C using a heating pad and a heating lamp throughout surgery. The right common carotid artery, external carotid artery, and internal carotid artery were exposed. A 4–0 nylon surgical suture with a silicon-coated tip was inserted from the external carotid artery to the beginning of the right middle cerebral artery. Surface blood flow was monitored before and during middle cerebral artery occlusion using a laser flowmeter (Omegawave, Tokyo, Japan) to ensure the success of occlusion. At 60 mins after the occlusion, the behavior of the operated rats was confirmed according to the method described earlier (Bederson et al, 1986). The rats with consistent circling toward their contralateral side and showing a reduced resistance to a lateral push toward the contralateral side were used in this study. After 90 mins of the occlusion, the suture was removed to allow reperfusion. Sham-operated animals received exactly the same surgical procedure without middle cerebral artery occlusion, whereas those did not undergo any surgical procedure were indicated as naive control animals. We used the non-operated naive animals as a control when the time course of changes in the amount of proteins and phosphorylation levels was examined. Time-matched sham-operated animals were used as a control when the effect of tyrosine kinase inhibitor on proteins of the isolated brain capillaries and on infarct volume after transient focal cerebral ischemia was determined at 6 h and/or 24 h of reperfusion. The amounts of protein and phosphorylation levels in the sham-operated animals were similar to those in the naive control animals. A total number of 92 rats were used in this study.
Administration of Src-Family Tyrosine Kinase Inhibitor
Src-family tyrosine kinase inhibitor 4-amino-5-(4-chlorophenyl)-7-(t-butyl) pyrazolo [3,4-d]pyrimidine (PP2; Biomol International L.P., Philadelphia, PA, USA) was dissolved in 1% dimethyl sulfoxide. The intravenous infusion of PP2 (30 μg/kg) or vehicle (1% dimethyl sulfoxide) through the left femoral vein was begun 10 mins before reperfusion.
The dose used in this study was based on the data obtained in our preliminary study showing that treatment with 3 μg/kg PP2 decreased the infarct size to a lesser degree than that with 30 μg/kg PP2. The average infarct volume of vehicle-treated, 3 μg/kg PP2-treated, and 30 μg/kg PP2-treated ischemic animals were 326.7 ± 31.0, 304.4 ± 13.7, and 111.1 ± 56.3 mm3 (n = 4 each). There was a significant difference in the infarct volume by groups (F2,9 = 9.8; P < 0.01). Fisher's protected least significant difference (PLSD) test revealed that the infarct volume of 30 μg/kg PP2-treated animals was significantly lower than that of vehicle-treated animals (P<0.05).
2,3,5-Triphenyltetrazolium Chloride Staining
To determine the infarct size after transient focal cerebral ischemia, we carried out TTC (2,3,5-triphenyltetrazolium chloride) staining of brain slices obtained from animals untreated or treated with the Src-family tyrosine kinase inhibitor. In brief, coronal sections with a 2-mm thickness were prepared at 24 h of reperfusion, and the slices were incubated with 2% TTC in physiologic saline. TTC-unstained areas were analyzed using an image analyzer (NIH image 1.63, NIH, Bethesda, MD, USA).
In Situ Vascular Permeability
We measured the area of fluorescein isothiocyanate-conjugated albumin (FITC-albumin, Sigma, Tokyo, Japan) leakage to assess the disruption of the BBB after ischemia. FITC-albumin was dissolved in 0.1 mol/L phosphate-buffered saline (pH 7.4) and injected into both carotid arteries by a modification of the method of Cavaglia et al (2001). Rats at 1, 6, 12, and 24 h of reperfusion were anesthetized with an intraperitoneal injection of 40 mg/kg sodium pentobarbital (Abbott Laboratories, Abbott Park, IL, USA). The bilateral carotid arteries were exposed and cannulated with a polyethylene tube (SP 31; Natume, Tokyo, Japan). FITC-albumin (10 mg/mL) was perfused through the carotid arteries at a rate of 1 mL/min (10 mL/kg) using a syringe pump (CFV-2100; NMON KOHDEN, Tokyo, Japan). The same amount of blood was withdrawn from the inferior vena cava to avoid increase of the systemic blood pressure. The brains were removed within 2 mins after perfusion of FITC-albumin, fixed for 24 h in 4% paraformaldehyde, and then cryoprotected by sequential immersion in 10, 20, and 30% sucrose solutions for 4h for each concentration. Then, they were embedded in Tissue-Tek O.C.T. compound (O.C.T. compound, SAKURA, Tokyo, Japan) and frozen in liquid nitrogen. Coronal brain sections (40 μm thick) were cut and thaw-mounted on poly-l-lysine-coated slide glasses (Muto Pure Chemicals, Tokyo, Japan), soaked in 0.1 mol/L phosphate-buffered saline for 30mins, and mounted with DAKO fluorescent mounting medium (DAKO Corporation, Carpinteria, CA, USA). Fluorescence was detected using a fluorescence microscope (BX52, Olympus, Tokyo, Japan). The area of FITC-albumin leakage regions (FITC-stained areas out of the total area occupied by cerebral vessels) was measured using an image analysis software (NIH image 1.63, NIH). The microscopic observations were carried out by a person blinded to the study group.
Isolation of Rat Brain Capillaries
Isolation of brain capillaries was carried out using a modified version of the method described earlier (Dallaire et al, 1991). Rats at 1, 6, 12, and 24 h of reperfusion were killed by decapitation, and their heads were quickly near-frozen in liquid nitrogen. The cortex was homogenized in ice-cold 15 mmol/L HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid), pH 7.4, containing 147 mmol/L NaCl, 4 mmol/L KCl, 3mmol/L CaCl2, 1.2 mmol/L MgCl2 (physiologic buffer). The homogenate was centrifuged at 3,500, g for 10 mins at 4°C. The pellets were resuspended in physiologic buffer containing 20% Ficoll T-400 (Sigma) and homogenized. After centrifugation at 25,000,g for 10 mins at 4°C, the pellets were resuspended in 15% dextran T-500 (Sigma). The suspension was layered onto 20% dextran T-500 and centrifuged at 25,000, g for 10 mins at 4°C. The pellets were resuspended in physiologic buffer as the brain capillaries.
Immunohistochemistry
Brain capillaries placed on Cell Tak (BD Bioscience, Franklin Lakes, NJ, USA)-coated slides were incubated for 2 h at 25°C. The attached brain capillaries were fixed for 1 h with 4% paraformaldehyde. For antigen retrieval, samples were incubated in 0.1 mol/L phosphate-buffered saline containing 0.25% Triton X-100 for 30 mins. After having been blocked with phosphate-buffered saline containing 10% bovine serum albumin, the brain capillaries were incubated overnight with a mouse monoclonal anti-rat endothelial cell antigen antibody (Serotec, Oxford, UK) at 4°C and then for 1 h with a Cy3-conjugated goat antimouse IgG antibody at 37°C. After having been washed with phosphate-buffered saline, they were mounted with DAKO fluorescent mounting medium (DAKO Corporation). Fluorescent images of capillaries were observed under an Olympus fluorescence microscope (BX-52, Olympus) equipped with a mercury arc lamp and incorporated into a personal computer using Viewfinder Lite and Studio Lite software (Pixcera Corporation, Los Gatos, CA, USA). Diameters of the isolated brain capillaries were measured in 50 randomly chosen areas from three separate experiments and expressed as the mean ± s.e.
Immunoblotting
Isolated brain capillaries were homogenized in ice-cold 12.5 mmol/L Tris-HCl, pH 7.4, containing 0.32mol/L sucrose, 2mmol/L sodium orthovanadate, 0.1 mmol/L phenylmethylsulfonyl fluoride, 20 mmol/L sodium diphosphate decahydrate, 20 mmol/L β-glycerophosphate, and 5 μg/mL each of leupeptin, aprotinin, and antipain. Homogenates were solubilized by heating at 100°C for 5 mins in the sample buffer (62.5 mmol/L Tris-HCl, pH 6.8, containing 2% sodium dodecyl sulfate, 5% β-mercaptoethanol, 5% glycerol), separated on polyacrylamide gels, and transferred to a polyvinylidene difluoride membrane. Protein blots were incubated with the appropriate antibodies, and the bound antibody was detected by the enhanced chemiluminescence method (Amersham Biosciences Inc., Piscataway, NJ, USA). Immunoblots were quantified using computerized densitometry and an image analyzer. Care was taken to ensure that bands to be semiquantified were in the linear range of response. To minimize between-blot variability, we applied an aliquot of pooled ‘control’ proteins obtained from naive control animals to one lane of every gel and calculated the immunolabeling of samples relative to this standard. Antibodies used for immunoblotting were anti-NeuN (Chemicon, Temecula, CA, USA), anti-occludin (Zymed Laboratories Inc., South San Francisco, CA, USA), anti-Tie2 (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), anti-Src, anti-phosphotyrosine (Upstate Biotechnology, Lake Placid, NY, USA), and anti-phospho-c-Src (Biosource International Inc., Camarillo, CA, USA) antibodies.
Immunoprecipitation
For immunoprecipitation of occludin, brain capillaries were solubilized by heating at 100°C for 5mins in 1% sodium dodecyl sulfate containing 1% β-mercaptoethanol; and then the solution was diluted 10-fold with 50mmol/L HEPES, pH 7.1, containing 1.5% Triton X-100, 150 mmol/L NaCl, 1.5 mmol/L MgCl2, 10% glycerol, 1 mmol/L concentration of each of NaF, sodium orthovanadate, phenylmethylsulfonyl fluoride, ZnCl2, and EGTA, and 5 μg/mL each of antipain, leupeptin, and aprotinin. The lysates were preincubated for 1 h at 4°C with protein G-agarose beads to remove any proteins that would bind nonspecifically to the beads. Anti-occludin antibody was added to the supernatant, and immunoprecipitation was allowed to proceed overnight at 4°C. Protein G-agarose beads were then added, and the immune complexes were isolated by centrifugation. The bound proteins were eluted by heating the beads at 100°C in the sample buffer.
Statistics
The results were expressed as the means ± s.e.m. Statistical comparison among multiple groups was evaluated by analysis of variance followed by post hoc Fisher's protected least significant difference test. Differences with a probability of 5% or less were considered significant (P<0.05).
Results
Effects of Transient Focal Cerebral Ischemia on Blood-Brain Barrier Leakage
At first, we examined BBB leakage at 1, 6, 12, and 24 h after the start of reperfusion after transient focal cerebral ischemia by injecting FITC-albumin into the carotid arteries. FITC-albumin leakage started at 6 h in the cerebral temporal cortex (Figure 1) and in the striatum, and lasted at least up to 24 h of reperfusion (Figure 1). There was no FITC-albumin leakage in the cerebral cortex or in the striatum in the contralateral hemisphere (data not shown).
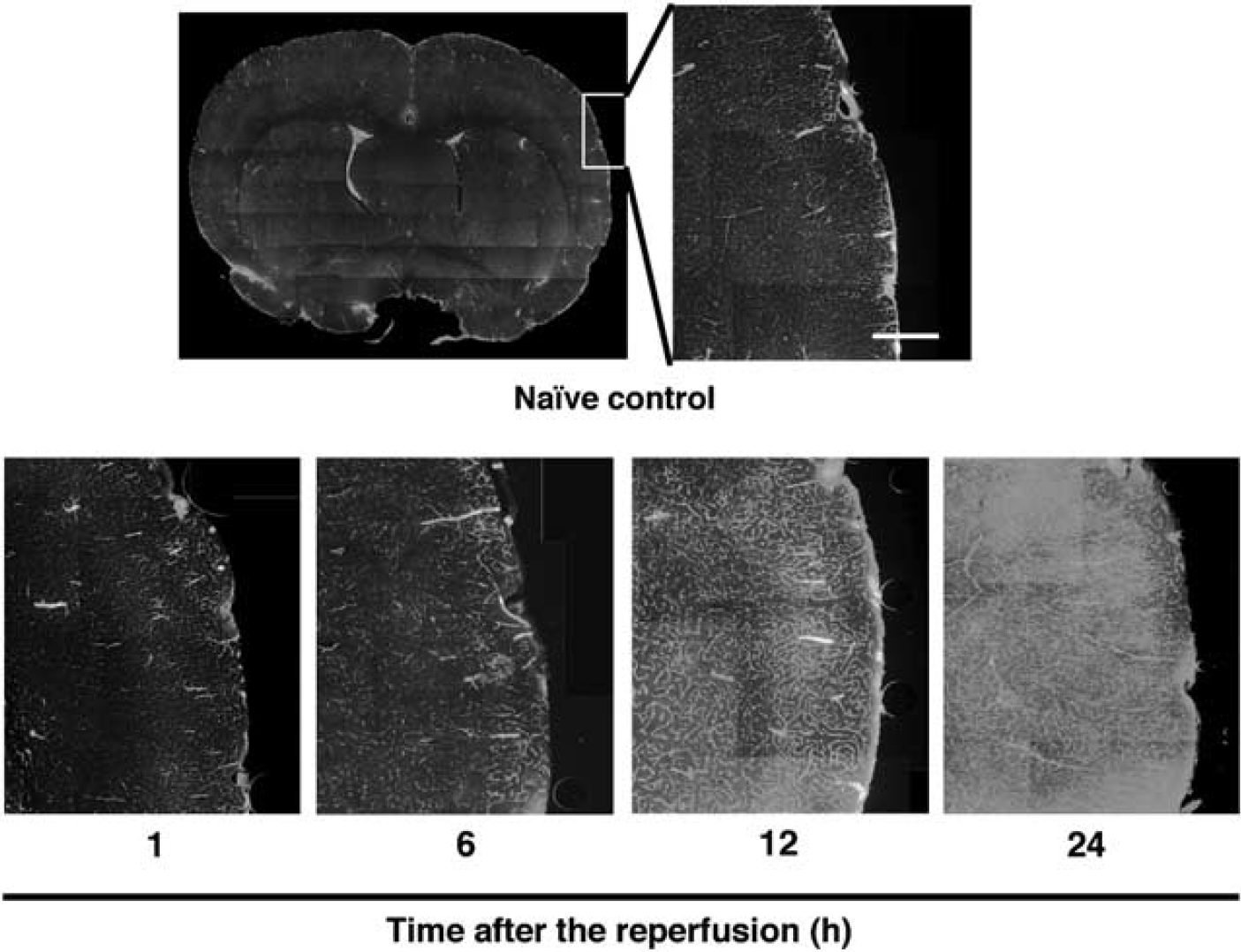
Time course of changes in the FITC-albumin leakage of middle cerebral artery-occluded rats. Photomicrographs of FITC-albumin-perfused vessels in the ipsilateral cortex from naive control and middle cerebral artery-occluded rats at 1,6, 12, and 24 h of reperfusion (n = 3 each). Scale bar = 500 μm.
Effects of Transient Focal Cerebral Ischemia on the Amount of Tight Junction Proteins in Brain Capillaries
To determine the pathophysiological changes in the level of the tight junctional protein occludin, we isolated brain capillaries from naive control and ischemic rats. Isolated brain capillaries were identified by immunoblotting for the expression of Tie2, which is a marker of endothelial cells. As shown in Figure 2A, Tie2 proteins were highly expressed in the isolated brain capillaries, whereas NeuN protein, a marker of neurons, was not detected (Figure 2A). We further labeled isolated brain capillaries with antibodies against rat endothelial cell antigen, another marker of endothelial markers (Figure 2B). The average diameter of the brain capillaries isolated from naive rats was 6.4 ± 0.4 μm, comparable with that from ischemic rats (6.4 ± 0.3 μm; Figure 2B). We also showed that there were no significant changes in the protein yield of isolated brain capillaries of ischemic rats compared with that of the naive rats up to 24 h of reperfusion (Figure 2C).
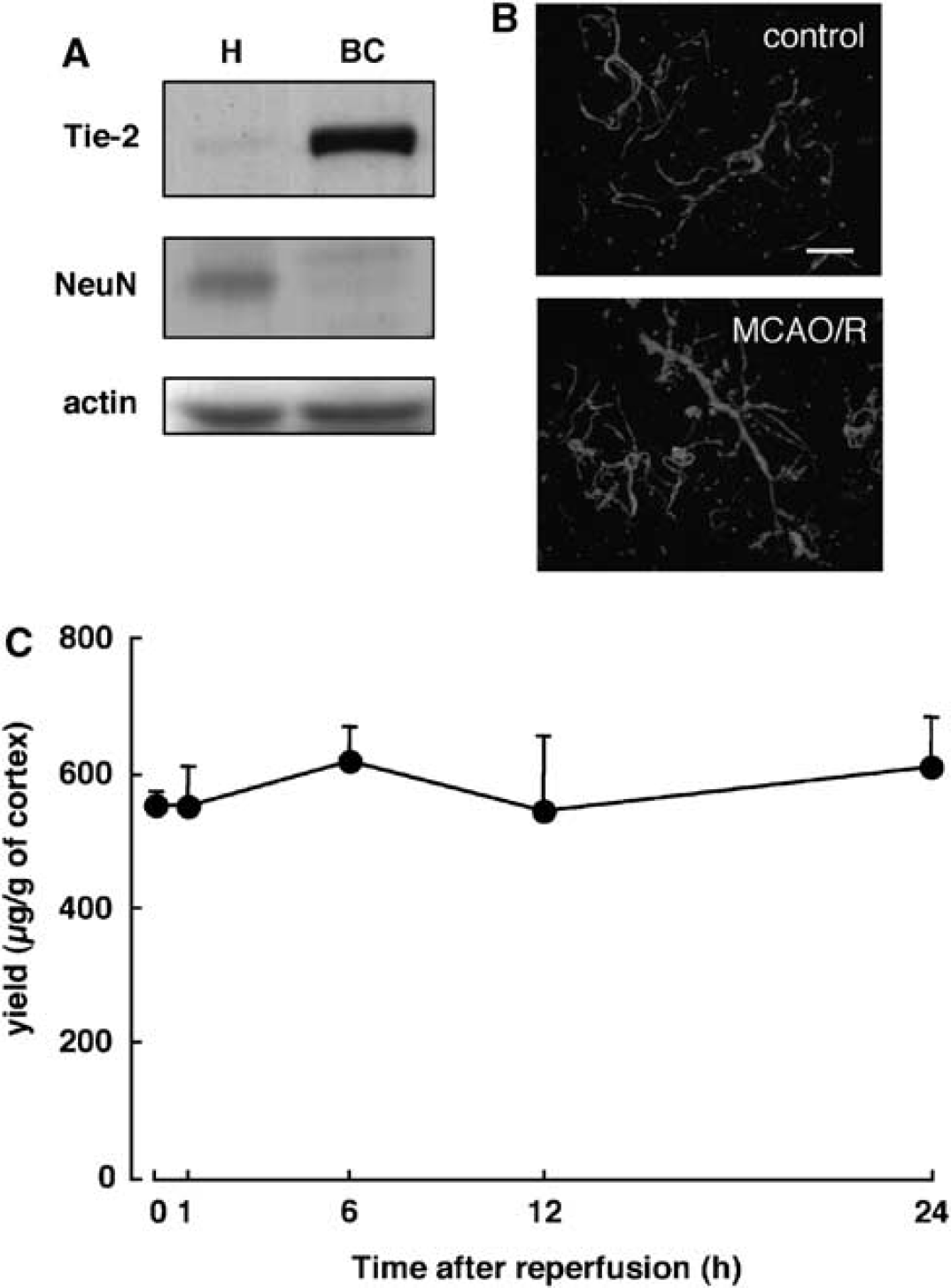
Characteristics of isolated brain capillaries of naive control and middle cerebral artery-occluded rats. (
Time Course of Changes in the Activity of Src in the Isolated Brain Capillaries after Transient Focal Ischemia
It was earlier reported that Src-mediated tyrosine phosphorylation of glutathione S-transferase (GST)-fused occludin protein appeared to reduce the ability of occludin to interact with ZO-1 in vitro (Kale et al, 2003), whose reduction could result in disruption of tight junctions. To investigate the possibility that Src-mediated tyrosine phosphorylation of occludin is related to disruption of the BBB, we determined the time course of changes in the level of activated Src by detecting the phosphorylation level at tyrosine 418 of Src (Salter and Kalia, 2004) in the isolated brain capillaries after transient focal cerebral ischemia. This level was increased at 6 h after the start of reperfusion after transient focal cerebral ischemia (Figure 3A). The average tyrosine phosphorylation of Src at 6 h of reperfusion was 247.7 ± 37.1% of that of the naive control rats (Figure 3B).
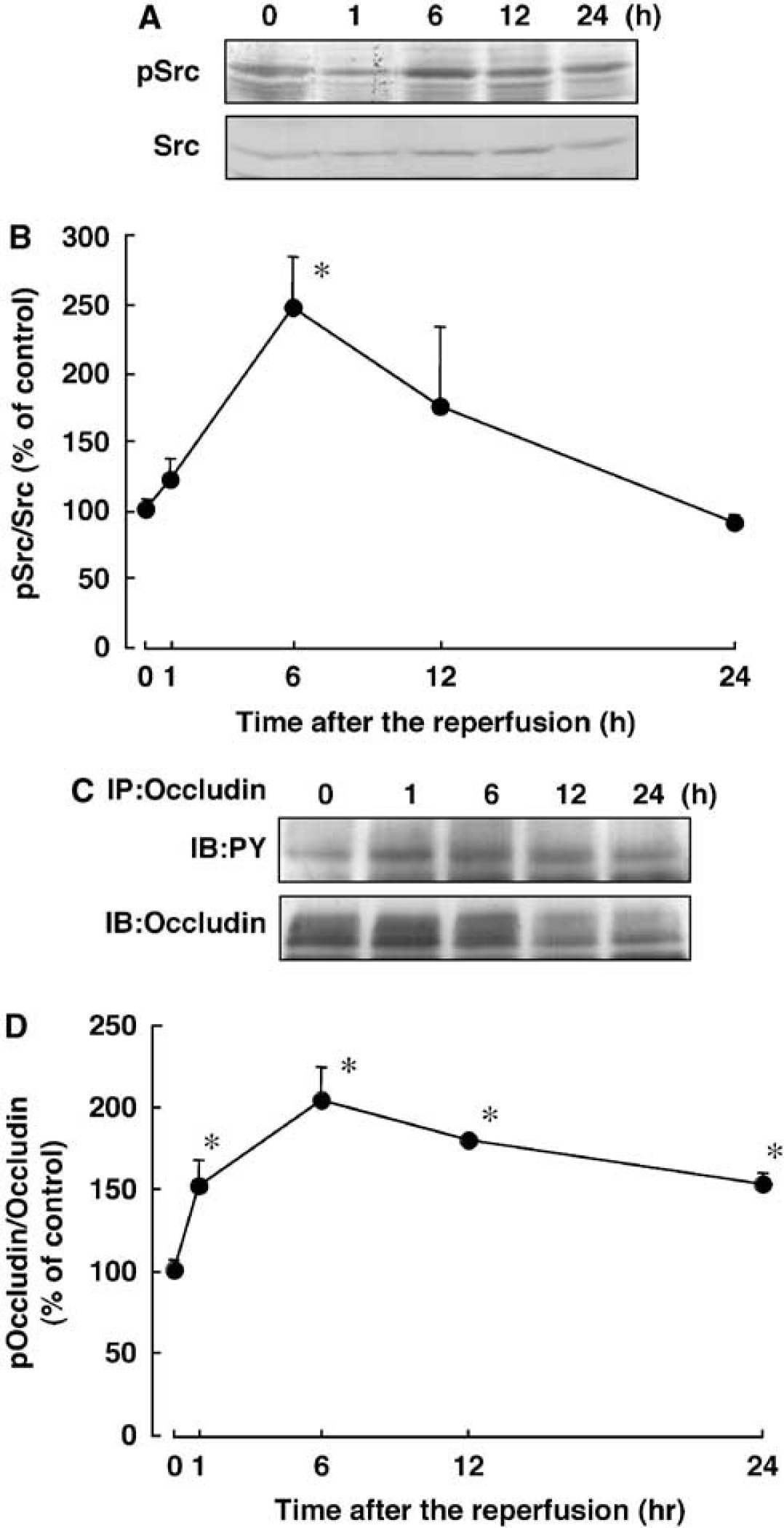
Time course of changes in the tyrosine phosphorylation of Src and occludin from isolated rain capillaries after transient focal cerebral ischemia. (
Time Course of Changes in Tyrosine Phosphorylation of Occludin in the Isolated Brain Capillaries after Transient Focal Ischemia
We next determined the time course of changes in the tyrosine phosphorylation of occludin in the isolated brain capillaries after transient focal cerebral ischemia. Using immunoprecipitation with occludin-specific antibodies followed by immunoblotting with anti-phosphotyrosine antibodies, a marked increase in the tyrosine phosphorylation of occludin was detected as early as 1 h of reperfusion, and this increased level was maintained for at least 24 h (Figures 3C and 3D). The maximum increase in it was detected at 6h of reperfusion (Figure 3D).
Effect of the Src-Family Tyrosine Kinase Inhibitor, PP2, on Tyrosine Phosphorylation of Occludin after Transient Focal Ischemia
As tyrosine phosphorylation of Src and occludin was prominent at 6 h of reperfusion (Figure 3), we next examined the effect of the Src-family tyrosine kinase inhibitor, PP2, on the tyrosine phosphorylation of Src and occludin at 6 h of reperfusion. As shown in Figures 4A and 4B, the ischemia-induced increase in the amount of tyrosine phosphorylated-Src in the isolated brain capillaries was reduced by treatment with PP2 (P<0.05).
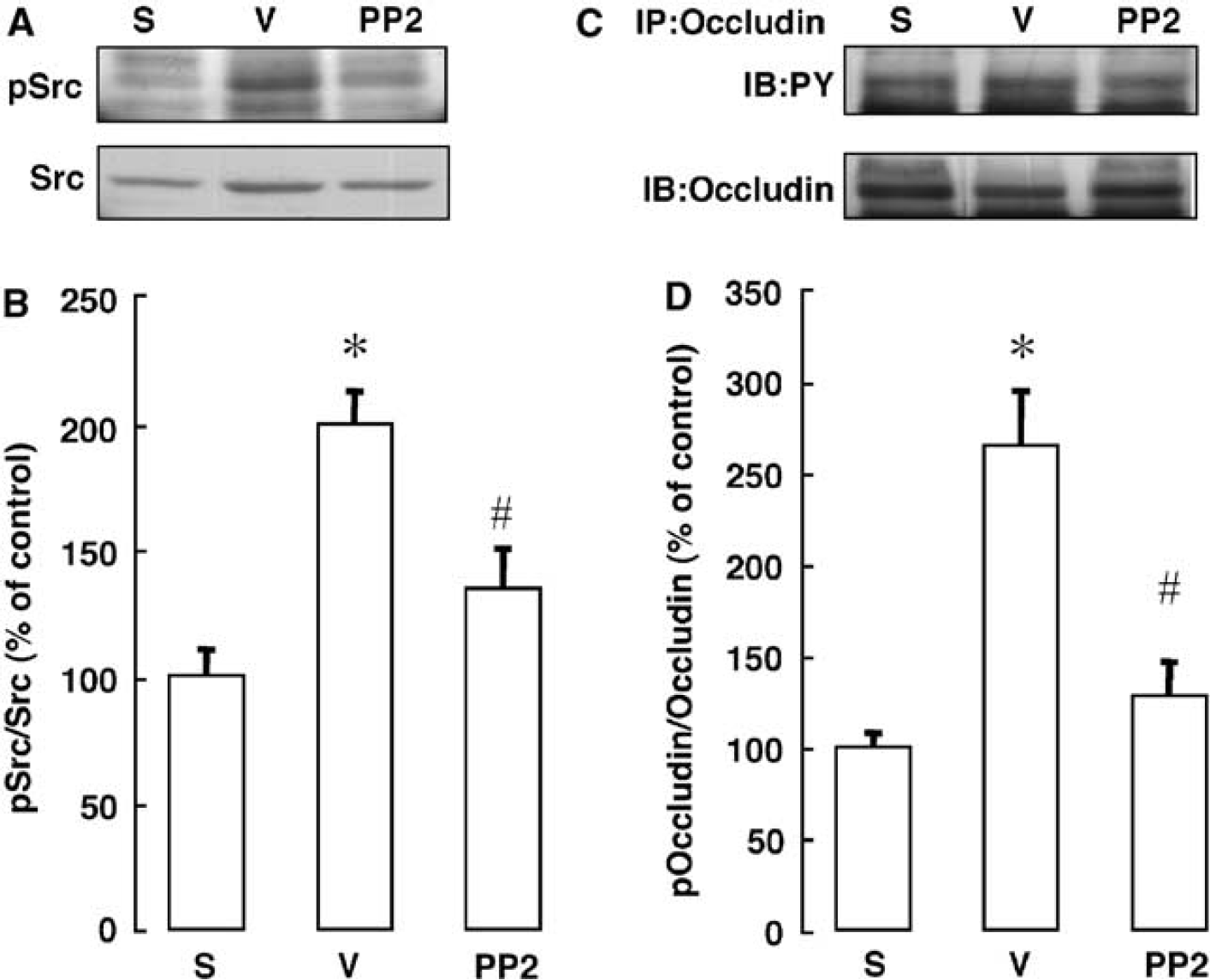
Effect of PP2 on the tyrosine phosphorylation of Src and occludin after transient focal cerebral ischemia. (
Furthermore, as shown in Figures 4C and 4D, the marked ischemia-induced increase in the tyrosine phosphorylation of occludin was reduced by treatment with PP2 (P<0.05).
Time Course of Changes in the Amount of Occludin in the Isolated Brain Capillaries after Transient Focal Ischemia
We next examined the time course of changes in the amount of occludin in the isolated brain capillaries after transient focal ischemia. This amount started to decrease at 1 h and remained significantly lower than that for the naive control for up to 24 h after the start of reperfusion after transient focal cerebral ischemia (Figures 5A and 5B).
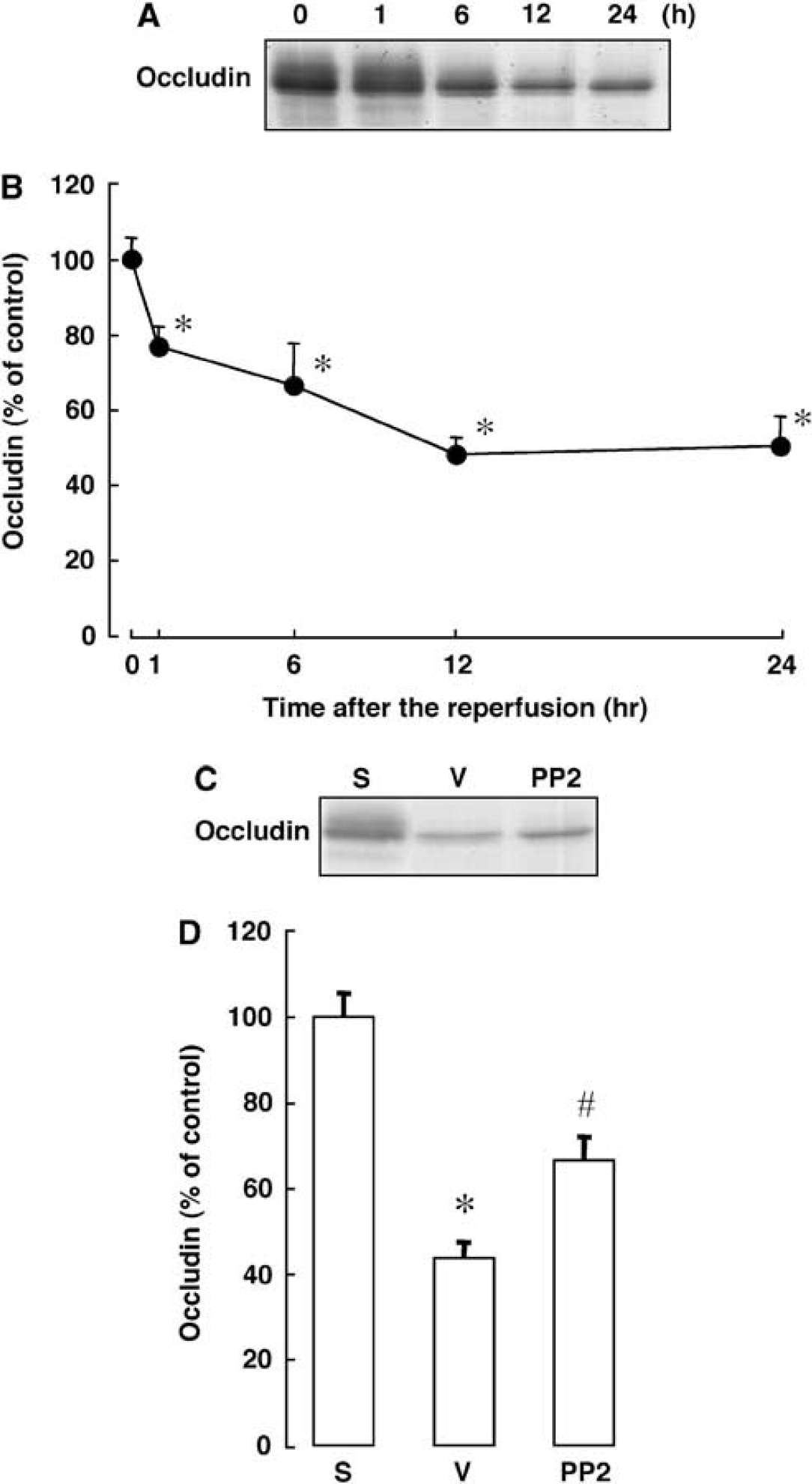
Time course of changes in the level of occludin and effect of PP2 on the level of occludin after transient focal cerebral ischemia. (
Effect of PP2 on the Amount of Occludin in the Isolated Brain Capillaries after Transient Focal Ischemia
As the ischemia-induced decease in the amount of occludin in the isolated brain capillaries was prominent at 12 and 24 h of reperfusion, we next examined the effect of PP2 on the amount of occludin in the isolated brain capillaries at 24 h of reperfusion. As shown in Figures 5C and 5D, this decrease in the amount of occludin was attenuated by treatment with PP2 (P < 0.05).
Effect of PP2 on FITC-Albumin Leakage and Infarct Volume after Transient Focal Ischemia
We further examined the effect of PP2 on FITC-albumin leakage and infarct volume at 24 h of reperfusion. As shown in Figures 6A and 6B, FITC-albumin leakage was detected in ∼ 75% of the ipsilateral hemisphere, and PP2 reduced this leakage (Figure 6B). PP2 also reduced infarct volume at 24 h of reperfusion (Figures 6C and 6D).
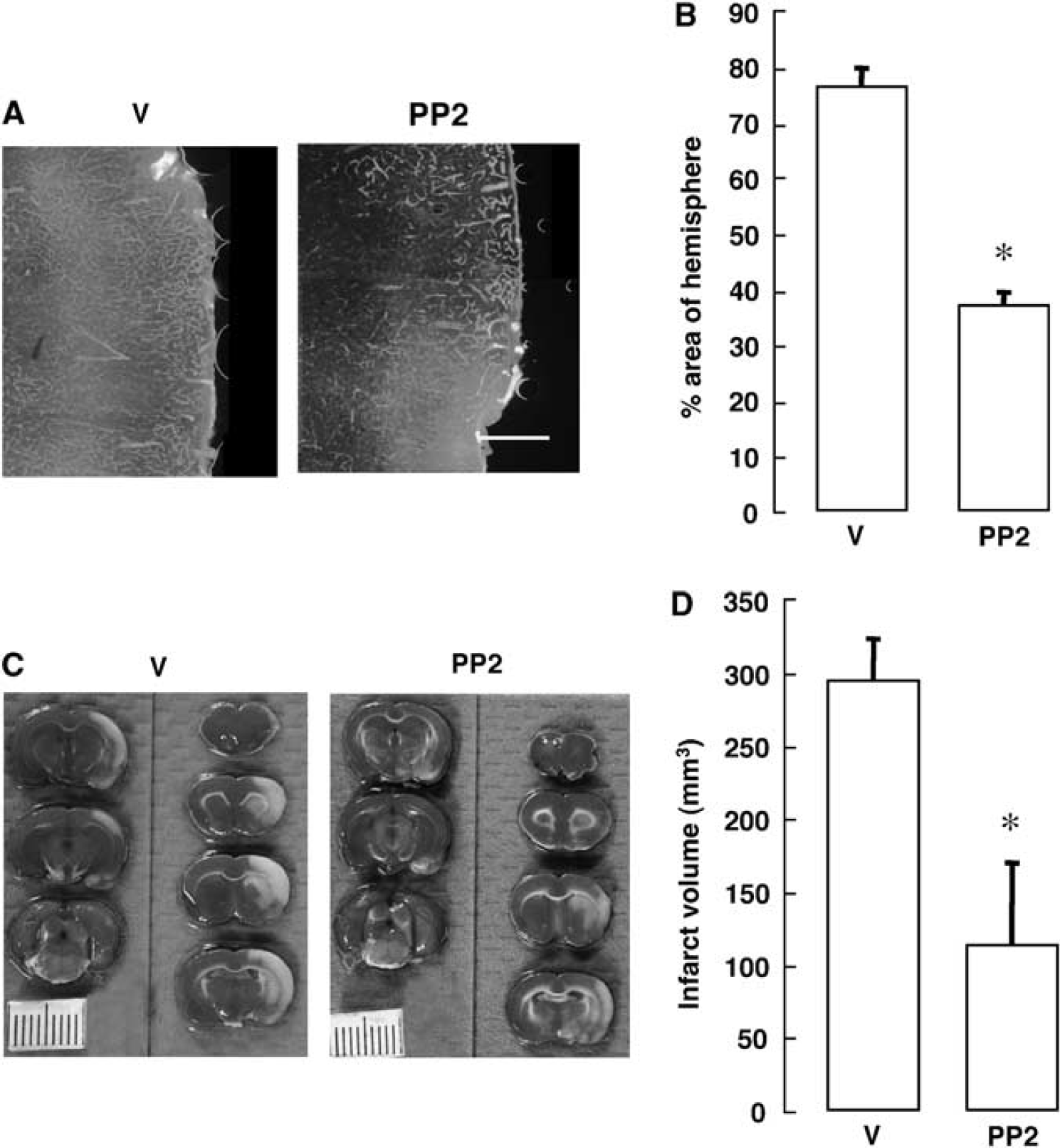
Effect of PP2 on the FITC-albumin leakage and on the infarct volume after transient focal cerebral ischemia. (
Discussion
In this study, we showed a marked increase in vascular permeability in the brain after transient focal cerebral ischemia, which was comparable with earlier findings that showed dysfunction of the BBB after cerebral ischemia (Belayev et al, 1996; Gotoh et al, 1985; Kago et al, 2006). It has been suggested that cerebral ischemia-induced pathophysiological events, which may lead to infarction and tissue degeneration, are associated with disruption of the BBB (Nag, 2002; Petty and Wettstein, 2001). Therefore, it is important to clarify the mechanism of the BBB disruption after cerebral ischemia to assess the appropriateness of therapeutic strategies. Accumulating evidence indicates that tight junctional proteins play a pivotal role in the maintenance of the physical barrier function of the BBB. Although phosphorylation of occludin, a tight junctional protein, affects paracellular permeability in microvessel endothelial cells, the contribution of tyrosine phosphorylation of occludin to in vivo ischemia-induced vascular permeability and the development of infarction remains to be determined. In this study, we sought to determine the role of tyrosine phosphorylation of occludin after ischemia by brain capillaries isolated from ischemic rats.
At first, we characterized brain capillaries isolated from naive control rats by staining them with an anti-rat endothelial cell antigen antibody. These capillaries had a diameter of 6.4 ± 0.4 μm, which is considered to be the general diameter of capillaries (Zhang et al, 2002). In addition, using markers for the endothelial cells and neurons, we showed that brain capillaries were concentrated in the brain capillary fraction used in this study. Furthermore, there was no significant different in the protein yield of isolated brain capillaries between naive control and ischemic rats. Therefore, the alterations in amounts of tight junctional proteins after ischemia are considered not to have been because of a variation in the amount of isolated brain capillaries.
Treatment with vanadate/H2O2 results in an increase in paracellular permeability in Madin-Darby canine kidney cells along with tyrosine phosphorylation of proteins located at intracellular junctions (Collares-Buzato et al, 1998). The finding raises the possibility that the increase in the activity of tyrosine kinase in the brain capillaries after transient ischemia may cause tyrosine phosphorylation of occludin and contribute to subsequent disruption of the BBB. In this sense, we showed that transient focal cerebral ischemia resulted in a marked increase in the tyrosine phosphorylation of occludin in the brain capillaries concomitant with the induction of FITC-albumin leakage. Treatment with the tyrosine kinase inhibitor inhibited not only Src activation but also tyrosine phosphorylation of occludin in the isolated brain capillaries. These findings identify the occludin molecule in brain capillaries as a target for tyrosine kinases after transient focal cerebral ischemia and suggest that tyrosine phosphorylation of occludin may play an important role in vascular permeability after cerebral ischemia.
It was shown earlier that peripheral inflammation decreases the expression of occludin in brain microvasculature samples and leads to increased BBB permeability (Huber et al, 2001). This decrease is comparable with that in the amount of occludin in brain capillaries after transient focal cerebral ischemia found in this study. The tyrosine phosphatase inhibitor, phenylarsine oxide, induces severe disruption of cell-cell contacts, which is accompanied by a decrease in the amount of occludin (Lohmann et al, 2004). Furthermore, c-Src-mediated tyrosine phosphorylation of GST-fused occludin protein appears to reduce the ability of occludin to interact with ZO-1 (Kale et al, 2003). These findings support our results that cerebral ischemia induced an increase in the tyrosine phosphorylation of occludin leading to the disruption of the BBB.
In this study, tyrosine phosphorylation of Src was increased only at 6 h of reperfusion compared with that of the naive control, whereas occludin was significantly phosphorylated throughout the experiment. Therefore, other tyrosine kinases or protein tyrosine phosphatases might also be involved in regulating the tyrosine phosphorylation of occludin in the brain capillaries after transient focal cerebral ischemia.
We showed that treatment with PP2 also reduced FITC-albumin leakage and infarct volume after transient focal cerebral ischemia. It is noteworthy that mice lacking c-Src or treated with a Src inhibitor show decreased infarct volumes after permanent middle cerebral artery occlusion (Paul et al, 2001). Cerebral ischemia results in a marked increase in the tyrosine phosphorylation of the N-methyl-D-aspartate receptor, a glutamate receptor, which is likely to enhance ion channel activities of the receptor and contribute to ischemic pathogenesis (Hisatsune et al, 1999; Murotomi et al, 2008; Salter and Kalia, 2004; Takagi et al, 1997, 1999, 2003). Although we cannot fully rule out roles of Src in BBB-independent signaling pathways that are important for neuronal cell function and tissue survival, our data point to a role of Src in tyrosine phosphorylation of occludinmediated disruption of the BBB leading to brain damage after cerebral ischemia.
In addition to phosphorylation at its tyrosine residues, occludin can also be phosphorylated on its threonine and serine residues by Rho kinase (Yamamoto et al, 2008), protein kinase C (Andreeva et al, 2001), by and casein kinase 2 (Cordenonsi et al, 1999). It appears that changes in phosphorylation by Rho kinase are accompanied by functional impairment of the barrier (Yamamoto et al, 2008).
In summary, we showed that inhibition of tyrosine phosphorylation of occludin by treatment with PP2 was accompanied by attenuation of functional impairment of the BBB and by decreased development of cerebral infarction. These results suggest that tyrosine phosphorylation of occludin would be a possible mechanism for disruption of the BBB after transient focal cerebral ischemia. Thus, the tyrosine phosphorylation of occludin in the brain capillaries may be a primary and/or an appropriate target to prevent disruption of the BBB permeability that leads to secondary ischemic cell damage.
Footnotes
The authors declare no conflict of interest.