
Abstract
The proapoptotic function of phosphatase and tensin homolog deleted on chromosome 10 (PTEN) phosphatase has been linked to its capacity to antagonize the phosphatidylinositol-3-kinase–Akt signaling pathway. Previous studies have shown that the Forkhead transcriptional factor (FOXO3a) is a critical effector of the PTEN-mediated tumor suppressor. However, whether the PTEN–Akt–FOXO3a pathway is involved in neuronal apoptosis in developing rat brain after hypoxia–ischemia (HI) is unclear. In this study, we generated an HI model using postnatal day 10 rats. Immunohistochemistry and western blot were used to detect the expression of total and phosphorylated PTEN, Akt, and FOXO3a, as well as its target gene
Introduction
Phosphatase and tensin homolog deleted on chromosome 10 (PTEN) was originally identified as a tumor suppressor because of its high frequency mutations in various types of tumors (Li et al, 1997). Its activity is regulated by the balance of phosphorylation and dephosphorylation at Ser residues at the C-terminal domain of the protein, which is important for the stability and function of the protein (Vazquez et al, 2000). Phosphorylation of the C-terminal domain of PTEN inactivates its phosphatase activity. Thus, dephosphorylated PTEN is an active form (Vazquez et al, 2001). The proapoptotic property of PTEN is mainly dependent on its lipid phosphatase activity, which antagonizes PI3K (phosphatidylinositol-3-kinase)–Akt and inhibits cell survival (Downes et al, 2001).
Activated Akt promotes cell survival by phosphorylating its substrates (Downward, 2004), including the mammalian members of the Forkhead transcription factors (FOXO). The FOXO family consists of three members, namely FOXO1a, FOXO3a, and FOXO4 (Hoekman et al, 2006). Among these members, FOXO3a has been shown to be important in the life span of tumor cells (Arden, 2006). While FOXO3a is phosphorylated by Akt, it will bind to the 14-3-3 protein and be retained in the cytoplasm (Van Der Heide et al, 2004). Conversely, dephosphorylation of FOXO3a causes its translocation from the cytoplasm to the nucleus, inducing its target genes, such as the proapoptotic protein, Bcl-2-interacting mediator of cell death (Bim), and Fas ligand (Gilley et al, 2003; Brunet et al, 1999). The upregulation of Bim can promote cytochrome
Previous studies have shown that tumor cells lack apoptosis in PTEN-mutated cell lines (Stahl et al, 2003). These findings suggest a role for PTEN in regulating neuronal apoptosis. Evidence has shown that a suppression of PTEN expression may provide neuroprotection in neurodegenerative disorders (Griffin et al, 2005; Zhu et al, 2007). In PTEN-mutated cells (characterized by the constitutive activation of Akt and inactivation of FOXO3a), cellular apoptosis was significantly inhibited (Nakamura et al, 2000), which suggests that FOXO3a is a proapoptotic effector of PTEN-mediated tumor suppressor (Nakamura et al, 2000). However, the underlying mechanisms of PTEN in regulating neuronal apoptosis after hypoxia–ischemia (HI) remain unclear. Therefore, we hypothesized that the PTEN–Akt–FOXO3a pathway is involved in neuronal apoptosis in the developing rat brain after HI. To test this hypothesis, we generated neonatal HI brain damage using postnatal day 10 rats to study this pathway in HI-induced neuronal apoptosis.
Materials and methods
Animal Protocols
All animal research was approved by the Sichuan University Committee on Animal Research. Female Sprague–Dawley rats with litters of mixed gender were acquired from the animal center of the Sichuan University (Chengdu, China). The mother was given food and water and was housed in a temperature- and light-controlled facility until the pups were 10-days-old. For the HI model, we used a method described previously (Ferriero, 2004). Briefly, each pup was anesthetized with halothane. With the pup supine, the right common carotid artery was exposed and permanently ligated with a 7-0 silk suture through amid line cervical incision. After ligation of the common carotid artery, the pups were returned to the dam for 1 h to recover from anesthesia. Duration of 2.5 h of hypoxia (8% O2/92% N2) was used to induce HI injury. For the bisperoxovanadium (bpv)-treated group, pups received intraperitoneal injections of bpv (20
Immunohistochemistry
Paraffin-embedded sections were used for immunohistochemistry as described previously (Li et al, 2007). Briefly, the sections were deparaffinized in xylene and dehydrated through 100% to 70% graded ethanol to distilled water, immersed in diluted antigen unmasking solution (Vector Laboratories, Foster City, CA, USA), and boiled for 30 mins in a microwave. After cooling, the sections were washed with phosphate-buffered saline. Endogenous peroxidase was inhibited with 0.3% hydrogen peroxide in methanol at room temperature for 20 mins. The sections were incubated using Clean Vision blocking solution (ImmunoVision Technologies, CA, USA) for 1 to 2 h. The following primary antibodies were used: rabbit anti-PTEN (Cell Signaling, Beverly, MA, USA, 1:100), rabbit anti-p-PTEN (ser 380) (Cell Signaling, 1:150), rabbit anti-p-FOXO3a (ser 253) (Cell Signaling, 1:100), and rabbit anti-Bim (Abcam, Cambridge, UK, 1:200). The sections were treated overnight at 4°C with primary antibodies in blocking solution. After washing in 0.05 mol/L phosphate-buffered saline, the sections were incubated with goat anti-rabbit IgG as a secondary antibody for 30 mins and then placed in a horseradish peroxidase complex solution for 30 mins. Peroxidase activity was shown by dipping the sections in a mixture containing 0.05% DAB (3-3′-diaminobenzidine, Dako, Carpinteria, CA, USA) and 0.03% H2O2 for 5 mins. The sections were then counterstained with hematoxylin, coverslipped, and observed under a microscope (Leica, CM 2000, Nussloch, Germany). Application of a control serum instead of the primary antibody on alternative sections of the same brain provided negative controls.
Western Blot Analysis
Isolated cortices at different time points as described above were homogenized in ice-cold lysis buffer containing cytosol extraction buffer that contained HEPES (pH 7.9; 10 mmol/L), KCL (10 mmol/L), EDTA (ethylenediaminetetraacetic acid; 0.1 mmol/L), EGTA (ethylene glycol tetraacetic acid; 0.1 mmol/L), DTT (dithiothreitol; 1 mmol/L), PMSF (phenylmethanesulfonyl fluoride; 0.5 mmol/L), protease inhibitor aprotinin (5
Reverse-Transcriptase PCR and Real-Time PCR
Rat cortices from sham controls, 0.5, 2, 4, 8 and 24 h after HI, as well as from bpv (pic) and NS-treated rats were removed and frozen for reverse transcriptase-PCR analysis as described previously (Li et al, 2008). Briefly, frozen samples were homogenized and total RNA was isolated using Trizol reagent (Gibco BRL, Gaithersburg, MD, USA). cDNAs were generated from 1
Bim: 5′-CACCCATGAGTTGTGACA-3′ (forward), 5′-TGGAAGCCATTGCACTGAG-3′ (reverse);
GAPDH: 5′-CCTCAAGATTGTCAGCAAT-3′ (forward), 5′-CCATCCACAGTCTTCTGAGT-3′ (reverse).
The reaction mixture for Bim real-time PCR contained 2
TUNEL Staining
Apoptotic cell death was detected using an
Results
Expression and Distribution of PTEN, p-PTEN, p-FOXO3a, and Bim after HI
To determine the expression of PTEN and its phosphorylation status (p-PTEN) in the P10 rat cortex after HI, we performed immunohistochemistry using paraffin-embedded sections from sham controls, as well as from control rats at 0.5, 2, 4, 8, and 24 h after HI (each group
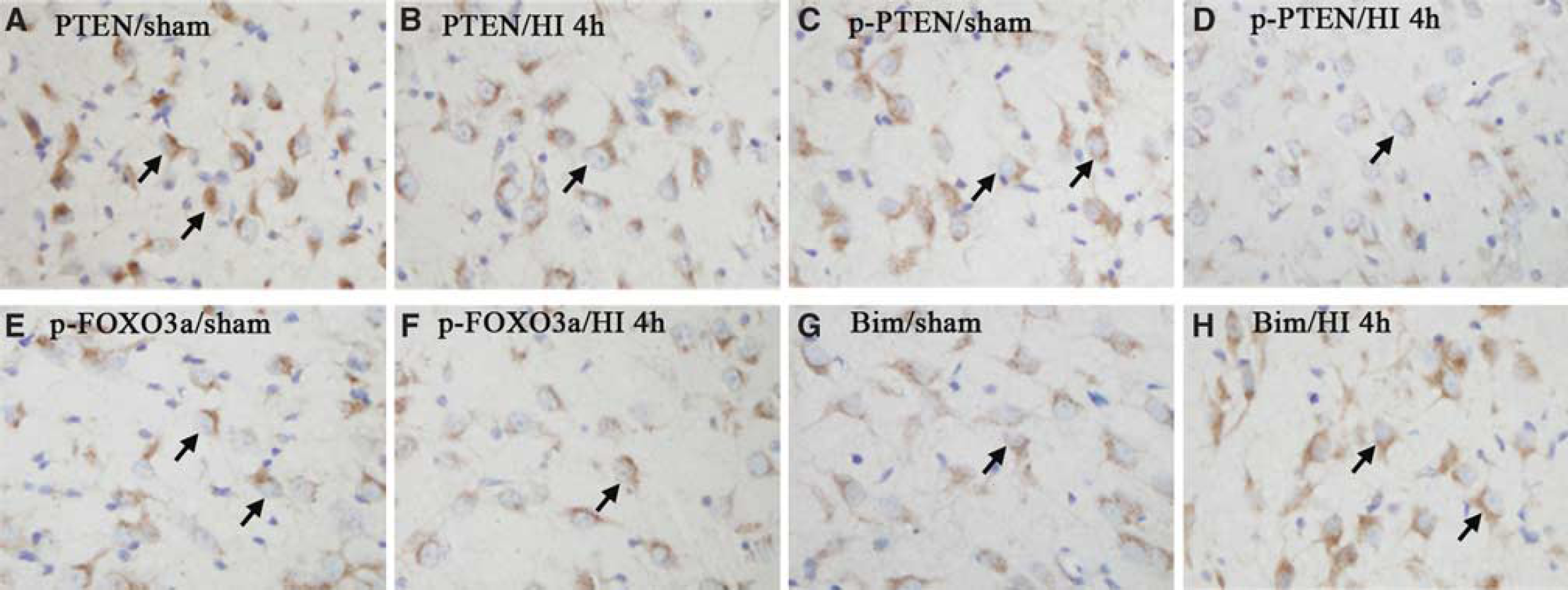
Immunoreactivity of PTEN, p-PTEN, p-FOXO3a, and Bim in P10 rat cortices was detected using immunohistochemistry (
To determine whether the phosphorylated form of FOXO3a (p-FOXO3a), a downstream transcriptional factor of PTEN (Nakamura et al, 2000), is regulated in this model, alternate slides from the brains as described above were used for immunohistochemistry detection. We found that the expression of p-FOXO3a was decreased at 0.5 and 2 h (data not shown), and reached the lowest at 4 h after HI (Figure 1F) compared with that of sham controls (Figure 1E). p-FOXO3a started to recover but still remained at a low level at 8 and 24 h (data not shown).
We further examined the expression of Bim, a target gene of FOXO3a, using immunohistochemistry. We found that the expression of Bim evidently increased from 0.5 h, (data not shown), and maintained at 4 h after HI (Figure 1H) compared with that of sham controls (Figure 1G). However, Bim expression declined fast to baseline at 8 h (data not shown).
Expression and Phosphorylation of PTEN, Akt, and FOXO3a Protein after HI
To quantify p-PTEN and PTEN expression after HI in this model, we measured the p-PTEN and PTEN protein expression using western blot analysis. Total protein was isolated using brain cortices from sham controls, as well as from control rats at 0.5, 2, 4, 8, and 24 h after HI (each group
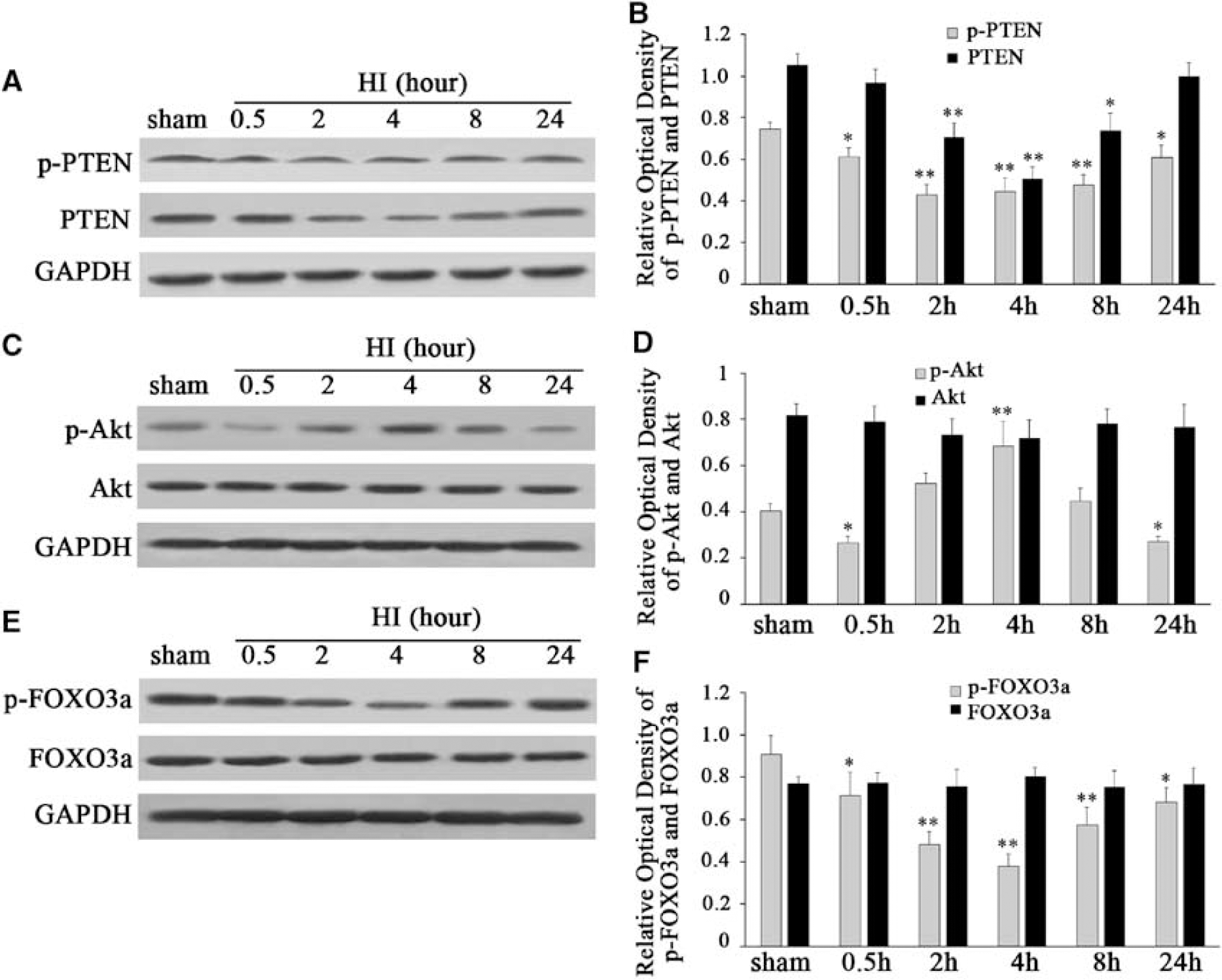
Western blot analysis of the expression and phosphorylation of PTEN, Akt, and FOXO3a in the hypoxic–ischemic cortex of P10 rats after HI (
As Akt is downstream of PTEN (Downes et al, 2001), we speculated whether the regulation of PTEN after HI would influence the activity of Akt. To investigate the changes of Akt activity after HI, we analyzed both phosphorylation of Akt (p-Akt) and total Akt protein expression using western blot analysis. We found that p-Akt significantly decreased at 0.5 h, but transiently increased at 4 h, and then decreased again at 24 h after HI (Figure 2C and 2D). After normalization with GAPDH, there was an ∼35% p-Akt decrease at 0.5 h, and a 33% decrease at 24 h after HI, compared with that in sham controls (F=31.843,
As FOXO3a has been identified as a principal substrate of Akt in neurons (Van Der Heide et al, 2004), we quantified both p-FOXO3a and the total FOXO3a protein expression after HI. We found that p-FOXO3a was significantly decreased at 0.5 and 2 h, and reached the lowest at 4 h (Figures 2E and 2F). p-FOXO3a started to recover but still remained at a low level at 8 and 24 h (Figures 2E and 2F). After normalization with GAPDH, there was an approximately 21% p-FOXO3a decrease at 0.5 h and a 25% decrease at 24 h after HI compared with that in sham controls (F=21.215,
HI Promotes FOXO3a Translocation from the Cytoplasm to the Nucleus and Induction of Bim
Dephosphorylation of FOXO3a has been reported to induce Bim expression and to lead to cell death in cultured cerebellar granule neurons deprived of growth factors (Brunet et al, 1999; Gilley et al, 2003). We speculated whether dephosphorylation of FOXO3a induced by HI would promote FOXO3a translocation into the nucleus and upregulate Bim transcription in this model. To answer this question, we extracted nuclear and cytosolic proteins from cortices and quantified FOXO3a expression separately in the nucleus and cytoplasm using western blot analysis (Figure 3A). We found that the nuclear protein of FOXO3a was obviously increased from 0.5 to 24 h in a time-dependent manner (Figures 3A and 3B). After normalization with GAPDH expression, we found an ∼1.7- and 2.9-fold nuclear protein of FOXO3a increase at 0.5 and 24 h, respectively, after HI compared with sham controls (F=32.071,
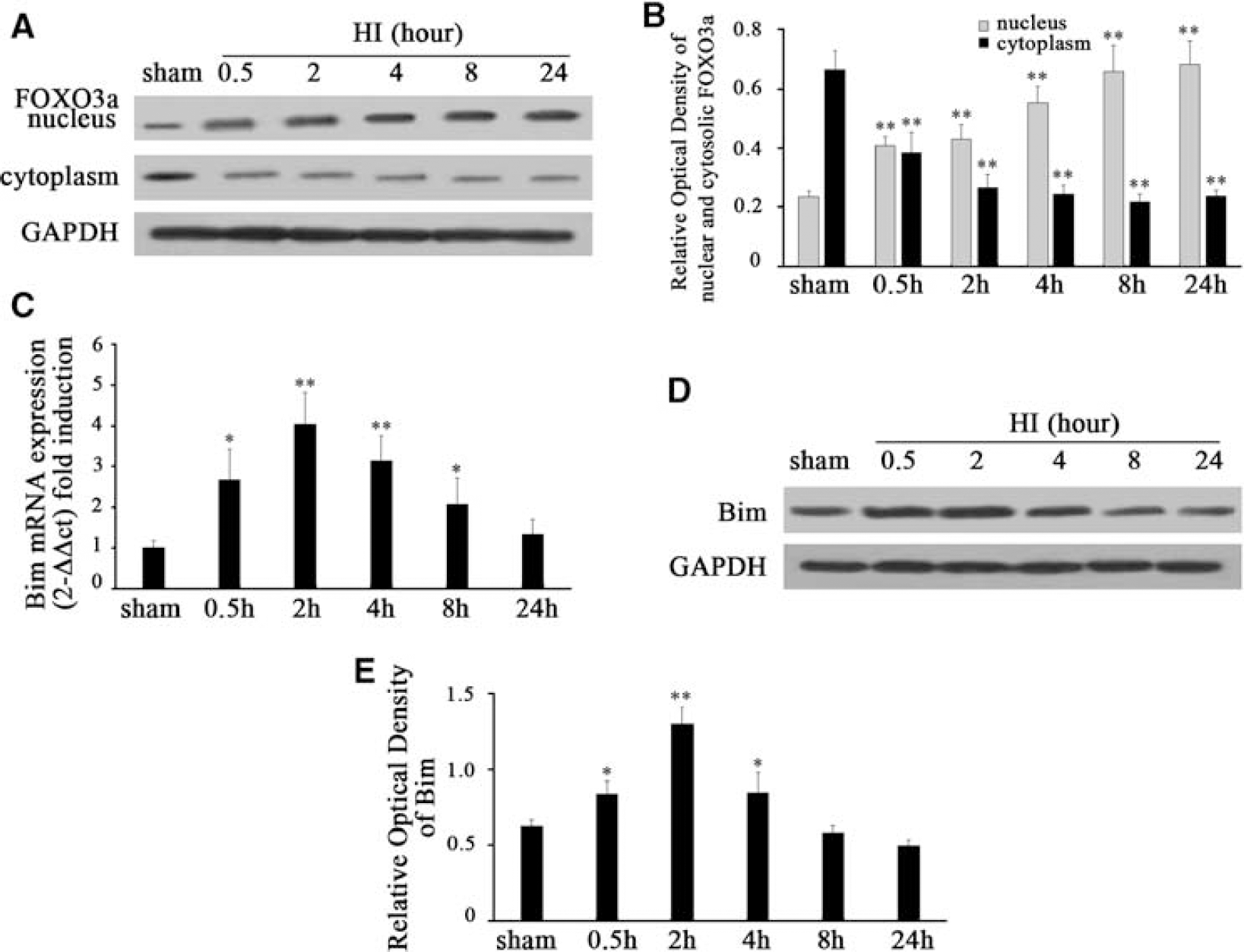
HI promotes FOXO3a translocation from the cytoplasm to the nucleus as well as the induction of Bim (
To determine whether
Similarly, Bim protein was significantly induced after HI (Figure 3D and 3E). After normalization with GAPDH expression, there was an ∼1.3- and 2.1-fold Bim protein increase at 0.5 and 2 h, respectively, after HI compared with sham controls (F=47.992,
Bisperoxovanadium Significantly Enhanced Phosphorylation of PTEN, Akt, and FOXO3a after HI
To further determine whether PTEN was involved in the regulation of Akt and FOXO3a after HI, we used a potent PTEN phosphatase inhibitor, bpv, which preferentially inhibits PTEN phosphatase activity in the nanomolar range without affecting cell viability (Schmid et al, 2004). In this study, 100 nmol/L of bpv was injected intraperitoneally before HI. We found that bpv pretreatment increased the expression of p-PTEN, p-Akt, and p-FOXO3a at 0.5 and 24 h after HI (Figures 4A, 4C, and 4E). After normalization with GAPDH, there was an ∼1.8- and 1.4-fold p-PTEN increase at 0.5 and 24 h (F=20.454,
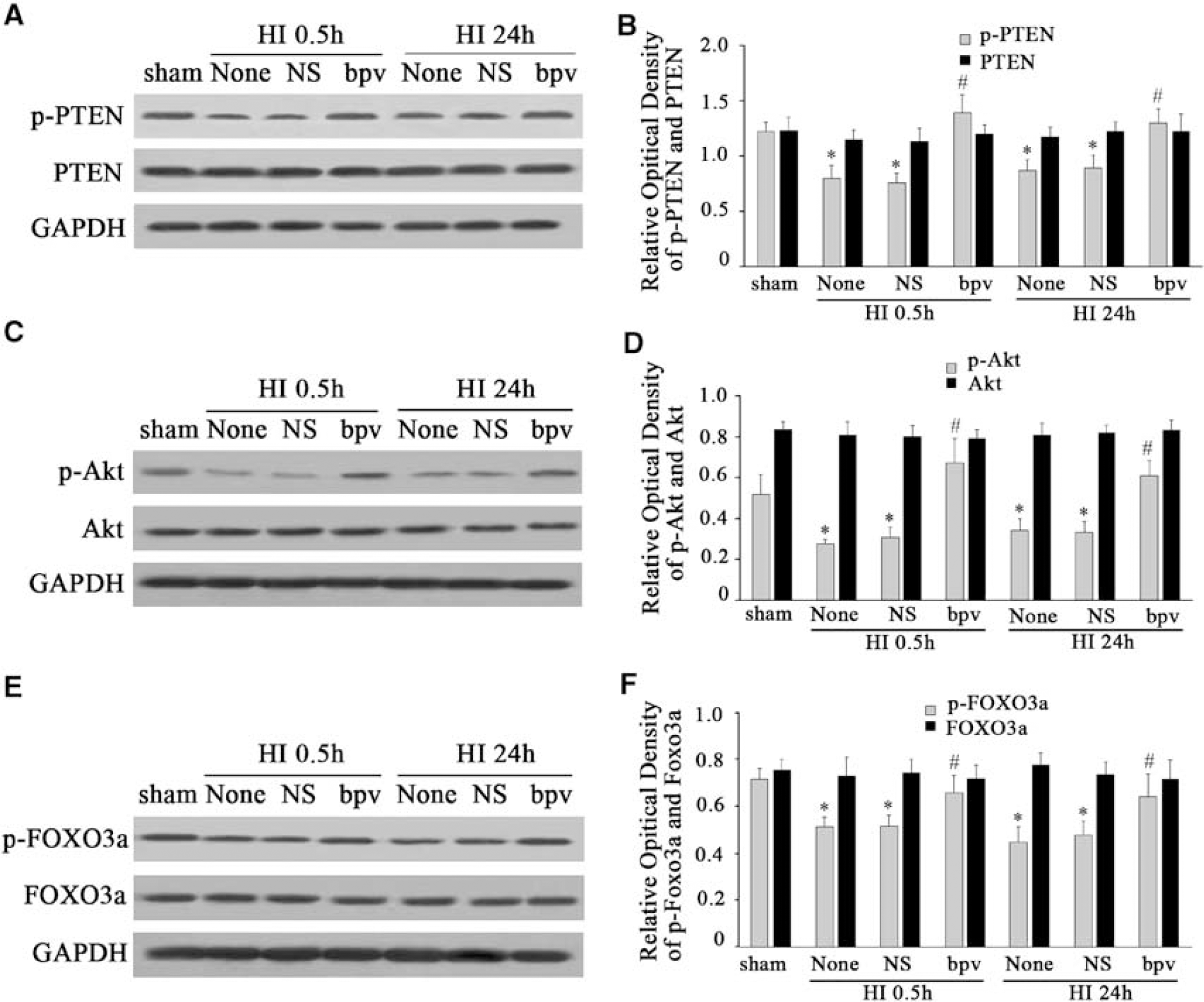
bpv pretreatment significantly enhanced the phosphorylation of PTEN, Akt, and FOXO3a after HI. bpv, a PTEN phosphatase inhibitor, was used to inhibit the activity of PTEN in this study. bpv can inhibit dephosphorylation of PTEN, the active form of PTEN. While the dephosphorylation form of PTEN was inhibited by bpv, the nonactive form of PTEN, the p-PTEN level increased. When p-PTEN increased, it promoted p-Akt expression and then phosphorylated FOXO3a. We found that bpv pretreatment increased the expression of p-PTEN, p-Akt, and p-FOXO3a at 0.5 and 24 h after HI. However, total PTEN, Akt, and FOXO3a protein remained unchanged (
Bisperoxovanadium Attenuated FOXO3a Translocation into the Nucleus and the Induction of Bim after HI
As bpv treatment rescued the expression of p-PTEN, p-Akt, and p-FOXO3a, we further investigated whether PTEN was involved in the regulation of FOXO3a translocation from the cytoplasm to the nucleus after HI. We found that FOXO3a was upregulated in the nucleus but downregulated in the cytoplasm at 0.5 and 24 h after HI (Figures 5A and 5B). However, bpv obviously blocked the upregulation of FOXO3a in the nucleus and the downregulation of FOXO3a in the cytoplasm (Figures 5A and 5B). After normalization with GAPDH expression, we found that bpv blocked ∼31 and 54% of the nuclear FOXO3a protein (F=32.768,
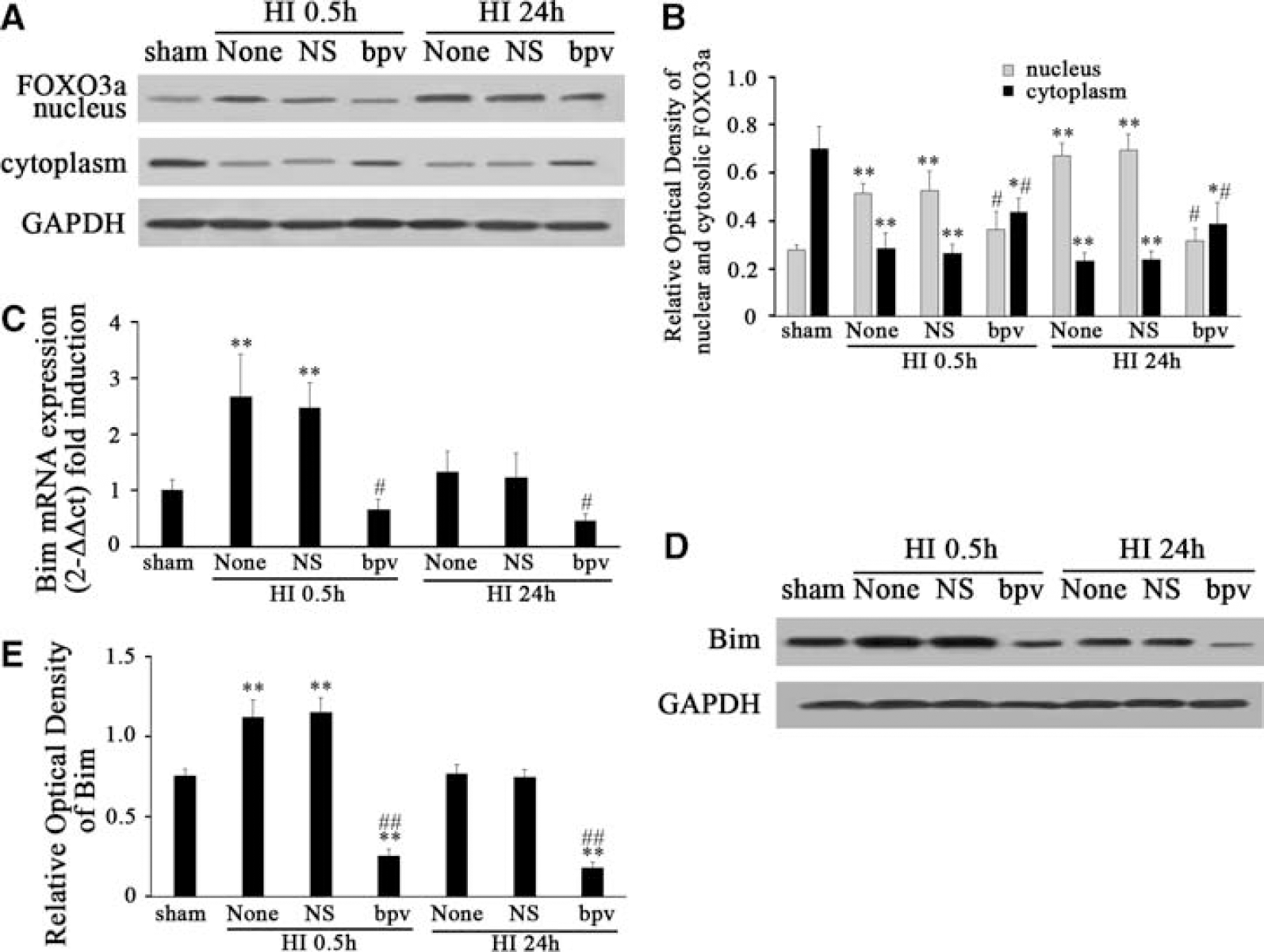
bpv pretreatment significantly attenuated FOXO3a translocation from the cytoplasm to the nucleus and the induction of Bim after HI (
To further understand whether PTEN blockage could regulate a proapoptotic gene,
Bisperoxovanadium Reduced Neuronal Apoptosis in the P10 Rat after HI
As PTEN activity blocked with bpv decreased the expression of the proapoptotic protein, Bim, we investigated whether cellular apoptosis could be blocked with the downregulation of Bim. Brain sections from sham controls and from control rats 0.5, 2, 4, 8, and 24 h after HI, as well as from bpv-treated sections were used to detect cellular apoptosis using TUNEL (terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling; each group
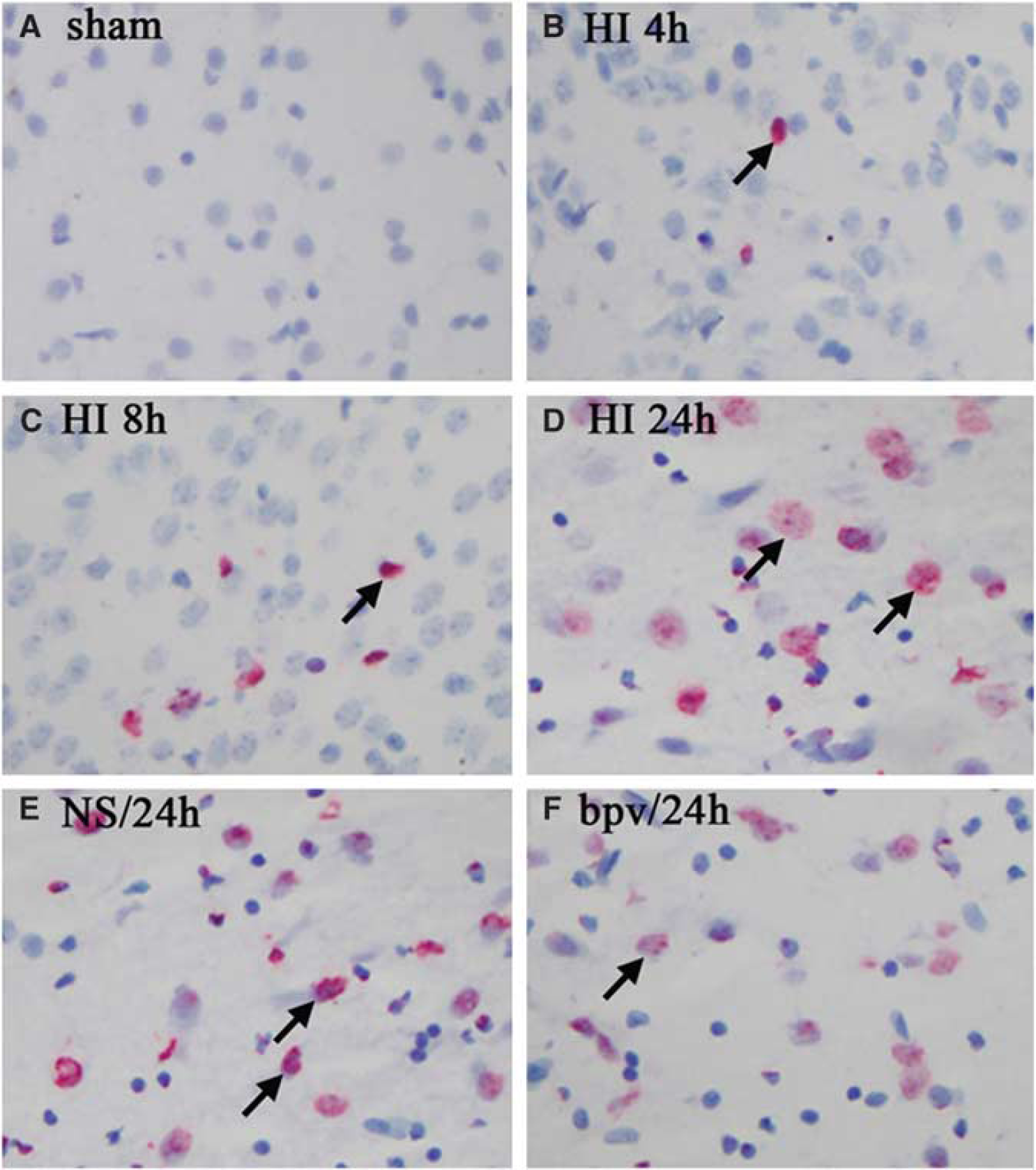
bpv pretreatment reduced neuronal apoptosis in P10 rat brain after HI. TUNEL-positive cells were hardly detected in the sham controls (
Discussion
Despite the progress in understanding the roles of tumor suppressor PTEN in normal brain function (Li et al, 2002), the roles and mechanisms of PTEN in the pathologic processes of neuronal apoptosis are not clear. In this study, we showed for the first time that the PTEN–Akt–FOXO3a pathway is involved in neuronal apoptosis in the developing rat brain after HI.
Evidence supporting the involvement of the PTEN–Akt–FOXO3a pathway in cellular apoptosis after HI is provided in this study. First, the phosphorylation level of PTEN, Akt, and FOXO3a decreased at an early stage of 0.5 h after HI, which was before cellular apoptosis, suggesting that the dephosphorylation of PTEN and its downstream components might be involved in mediating HI brain damage. Second, the HI-induced dephosphorylation of FOXO3a was accompanied by an increase in FOXO3a translocation from the cytoplasm to the nucleus. Nuclear translocation of FOXO3a induced Bim expression, indicating that FOXO3a dephosphorylation might contribute to cellular apoptosis. Third, pretreatment of rats with bpv, which is a potential inhibitor of PTEN activity, improved p-PTEN, p-Akt, and p-FOXO3a, leading to a decrease in FOXO3a translocation from the cytoplasm to the nucleus. Finally, PTEN inhibition with bpv decreased early cellular apoptosis after HI. Therefore, PTEN might have a critical role in regulating the Akt–FOXO3a pathway involved in cellular apoptosis in the developing rat brain after HI.
Phosphatase and tensin homolog deleted on chromosome 10 is a negative regulator of the PI3K–Akt pathway that prevents the recruitment of Akt to the cellular membrane for phosphorylation (Fresno Vara et al, 2004). However, when PTEN was phosphorylated, its interaction with phosphatidylinositolphosphate (PIP), a substance of PTEN, will be decreased and will lead to a decrease in its activity (Vazquez et al, 2001). Therefore, dephosphorylation of PTEN indicates an increase in its activity, which might have a role in cell growth. We found that p-PTEN but not total PTEN decreased early at 0.5 h, which indicates that PTEN was dephosphorylated at 0.5 h after HI (Figure 2). Dephosphorylation of PTEN is a signal for PTEN degradation, as dephosphorylated PTEN is degraded by proteasome (Torres and Pulido, 2001). Our findings that p-PTEN decreased after HI are consistent with previous findings in an adult global ischemic rat model (Zhang et al, 2007). Although there are controversial reports that p-PTEN increased at 24 h after focal ischemia (Omori et al, 2002), we believe that these different findings may be due to the difference in animal age and the model of ischemia.
Akt is a downstream factor of PTEN, which has a critical role in controlling the balance between survival and apoptosis by regulating the phosphorylation of its downstream components (Franke et al, 2003). In this study, p-Akt decreased at an early stage at 0.5 h and at a later stage at 24 h after HI, which is consistent with the changes of p-PTEN. This suggests that the decrease in p-Akt after HI may be related to the decrease in p-PTEN. However, it is not clear whether the decrease in p-Akt will have a role in the pathologic changes of neonatal brain after HI. We detected the regulation of transcriptional factor FOXO3a, a downstream event of p-Akt. FOXO3a is known to have an important role in cellular apoptosis (Huang and Tindall, 2007). We found that p-FOXO3a decreased starting at 0.5 h and maintained at a low level at 24 h after HI, which is consistent with the changes of p-Akt. As p-FOXO3a, the inactive form of FOXO3a, decreased and total FOXO3a was not changed, dephosphorylation of FOXO3a, the active form of FOXO3a, increased. We then investigated whether dephosphorylation of FOXO3a leads to its translocation from the cytoplasm to the nucleus and activates its target gene
Next, we tried to investigate whether the PTEN signaling pathway was involved in cellular apoptosis in this model. The PTEN phosphatase inhibitor, bpv, was used before HI treatment. Bisperoxovanadium can inhibit the dephosphorylation of PTEN, the active form of PTEN, and can lead to an increase in the p-PTEN level. We found that bpv increased the expression of p-PTEN, p-Akt, and p-FOXO3a at 0.5 and 24 h after HI (Figures 4A, 4C, and 4E). Meanwhile, we found that bpv significantly attenuated the expression of FOXO3a in the nucleus but increased it in the cytoplasm, which indicates that bpv attenuated FOXO3a translocation to the nucleus (Figure 5A). The reduction of FOXO3a translocation by PTEN inhibition might be mediated through increased p-PTEN, leading to the enhancement of p-Akt and p-FOXO3a, as the increase in p-FOXO3a will lead to FOXO3a retention in the cytoplasm (Van Der Heide et al, 2004). We further showed whether the reduction of FOXO3a translocation by bpv could downregulate its target gene,
In this study, we also found that p-Akt was increased transiently at 4 h after HI. However, p-FOXO3a was not increased accordingly. The possible reason for this incompatibility may be a temporal increase in p-Akt expression but not the increase in its activity, as the p-Akt level may not entirely represent its kinase activity (Hill et al, 2001). This transient increase in the p-Akt level was also founded in an adult rat ischemic model by Zhao et al (2005) who found that Akt activity was actually decreased after ischemia.
In summary, we have shown that the PTEN–Akt–FOXO3a pathway is involved in neuronal apoptosis in the developing rat brain after HI. Agents targeting PTEN might help to study the protective mechanisms in neonatal HI.
Footnotes
The authors declare no conflict of interest.