
Abstract
MicroRNAs (miRNAs) are ∼22 nucleotides long, noncoding RNAs that control cellular function by either degrading mRNAs or arresting their translation. To understand their functional significance in ischemic pathophysiology, we profiled miRNAs in adult rat brain as a function of reperfusion time after transient middle cerebral artery occlusion. Of the 238 miRNAs evaluated, 8 showed increased and 12 showed decreased expression at least at 4 out of 5 reperfusion time points studied between 3 h and 3 days compared with sham. Of those, 17 showed > 5 fold change. Bioinformatics analysis indicated a correlation between miRNAs altered to several mRNAs known to mediate inflammation, transcription, neuroprotection, receptors function, and ionic homeostasis. Antagomir-mediated prevention of mir-145 expression led to an increased protein expression of its downstream target superoxide dismutase-2 in the postischemic brain.
The mRNA levels and function are known to be controlled by several factors including transcription factors, histones, DNA methylation, and RNases. In addition, recent studies indicated that mRNAs are finely controlled by a group of small noncoding, evolutionarily conserved RNAs of ∼22 nucleotides (nt) long called microRNAs (miRNAs) (Fire et al, 1998; Caplen et al, 2001; Grishok et al, 2001; Bartel, 2004; Ambros, 2004). Although their mechanism of action is not completely elucidated, miRNAs are thought to modulate mRNA function by binding to an 8-base pair complementary seed region in the 3'UTR. The binding of miRNAs is thought to either degrade the mRNAs or inhibit their translation (Humphreys et al, 2005; Jing et al, 2005). In addition, recent studies suggested that specific miRNAs can bind to target regions of the promoters of certain genes to induce their expression (Place et al, 2008) as well as can modulate gene expression by binding to a protein-coding region (Duursma et al, 2008). Several miRNAs can bind to a specific mRNA, and a single miRNA can bind to several mRNAs. Structural studies estimated that most miRNAs can act on ∼200 mRNAs (Lewis et al, 2003). Because of this redundancy in function, the small number of miRNAs is thought to effectively control the huge number of mRNAs either sequentially or simultaneously. In addition, certain mRNAs and/or their protein products can control the expression of specific miRNAs in a feed-forward mechanism (Fazi et al, 2005). Many miRNAs were shown to control transcription factors that in turn control thousands of downstream genes (Miska et al, 2004; Krichevsky et al, 2006; Rogaev, 2005). Thus, miRNAs can significantly influence the cellular homeostasis under normal physiologic conditions.
Recent studies showed that many pathologic conditions significantly alter cerebral miRNA profiles, which might have profound effect on the disease outcome. It was shown that miRNAs are involved in brain tumor growth (Nicoloso and Calin, 2008), Alzheimer disease (Hébert et al, 2008), Down syndrome (Kuhn et al, 2008), schizophrenia (Beveridge et al, 2008), and stroke (Jeyaseelan et al, 2008).
An understanding of the functional significance of miRNAs will complement the existing knowledge gained from the mRNA studies to provide better therapeutic targets to prevent the secondary neuronal death after stroke. Hence, we presently analyzed the miRNA profiles as a function of reperfusion time after transient middle cerebral artery occlusion (MCAO) in adult rats. Using bioinformatics tools, we evaluated the target mRNAs and gene promoters of the miRNAs altered in the ischemic brain. We further evaluated the effect of preventing mir-145 induction on its downstream target superoxide dismuate-2 (SOD2) expression employing the antagomir strategy.
In the nucleus, the miRNAs are transcribed as hairpin clusters of primary miRNAs (pri-miRNAs; 5'-capped polyadenylated transcripts), which will be converted to 70-nt stem loop structures (pre-miRNAs) by Drosha (a type-III RNase) in association with a cofactor Pasha (
Materials and methods
Focal Ischemia
Transient MCAO was induced in adult male spontaneously hypertensive rats (280–300 g; Charles River, Wilmington, MA, USA) under halothane anesthesia by the intraluminal suture method as described earlier (Vemuganti et al, 2004; Dhodda et al, 2004; Satriotomo et al, 2006; Kapadia et al, 2006; Tureyen et al, 2007, 2008). All the surgical procedures were approved by the Research Animal Resources and Care Committee of the University of Wisconsin-Madison and the animals were cared for in accordance with the
MicroRNA Microarray Analysis
Groups of rats subjected to transient MCAO were killed at various reperfusion periods (3, 6, 12, 24, and 72 h;
The miRNA hybridization data were corrected by subtracting the background (calculated from the median of 5%–25% of the lowest-intensity cells) and normalized to the statistical median of all detectable transcripts using the locally weighted regression method, which balances the intensities of Cy5 labeled transcripts so that the differential expression ratios can be properly calculated (Bolstad et al, 2003). For background subtracting, the background was defined on each array as the average signal of the BKG0 spots (chemical linkers without the probes). The hybridization intensities above exp(5) (∼150) were considered as significant as described earlier (Vagin et al, 2006), and intensities below 30,000 were considered as nonsaturated, as established with titration of several synthetic 20-nt RNA oligos (external controls) spiked into each sample. In addition, on each array there were 16 sets of spatially distributed internal control probes. These include PUC2PM-20B and PUC2MM-20B, which are the perfect match and the single-base mismatch sequences, respectively. The stringency of the intensity ratio of the PUC2PM-20B and PUC2MM-20B is expected to be larger than 30 indicating proper hybridization in each case. For proper analysis of signal intensities on each chip, both the internal controls and the test miRNA probes were repeated 12 times. On a microarray, the hybridization signal was linearly obtained from 1 to ∼66,000 units. An miRNA transcript was considered detectable if it met the following criteria: (a) Signal intensity higher than three times the maximal background signal, (b) spot CV < 0.5 (CV was calculated as (standard deviation)/(signal intensity)), and (c) the signals from at least 50% of the 12 redundant repeating probes above detection level. To avoid false positives, any spot that deviated <50% from the average value of the 12 repeating spots and/or spots with CV > 0.5 were eliminated.
The data from different groups were normalized and analyzed statistically using ANOVA. The normalized hybridization data of the 36 microarrays (
Real-Time Polymerase Chain Reaction
Real-time polymerase chain reaction (PCR) was performed to confirm the microarray data. The reverse transcription was performed using the TaqMan MiRNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) at 16°C for 30 mins, 42°C for 30 mins, and 85°C for 5 mins in buffer containing 10 ng low-molecular weight enriched RNA, 100 mmol/L dNTPs (with dTTP), 50U reverse transcriptase, 0.4U RNase inhibitor, and reverse transcriptase (RT) primer. PCR reactions were performed using the TaqMan® MiRNA Assay Kit (Applied Biosystems). Briefly, each reaction contained 10 µL TaqMan 2 × Universal PCR Master Mix, 1 µL 20 × TaqMan® MicroRNA Assay reagent and 1.33 µL of the RT reaction product in a total volume of 20 µL. Real-time PCR was conducted at 95°C for 10 mins, followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. The threshold cycle (
In Situ Hybridization
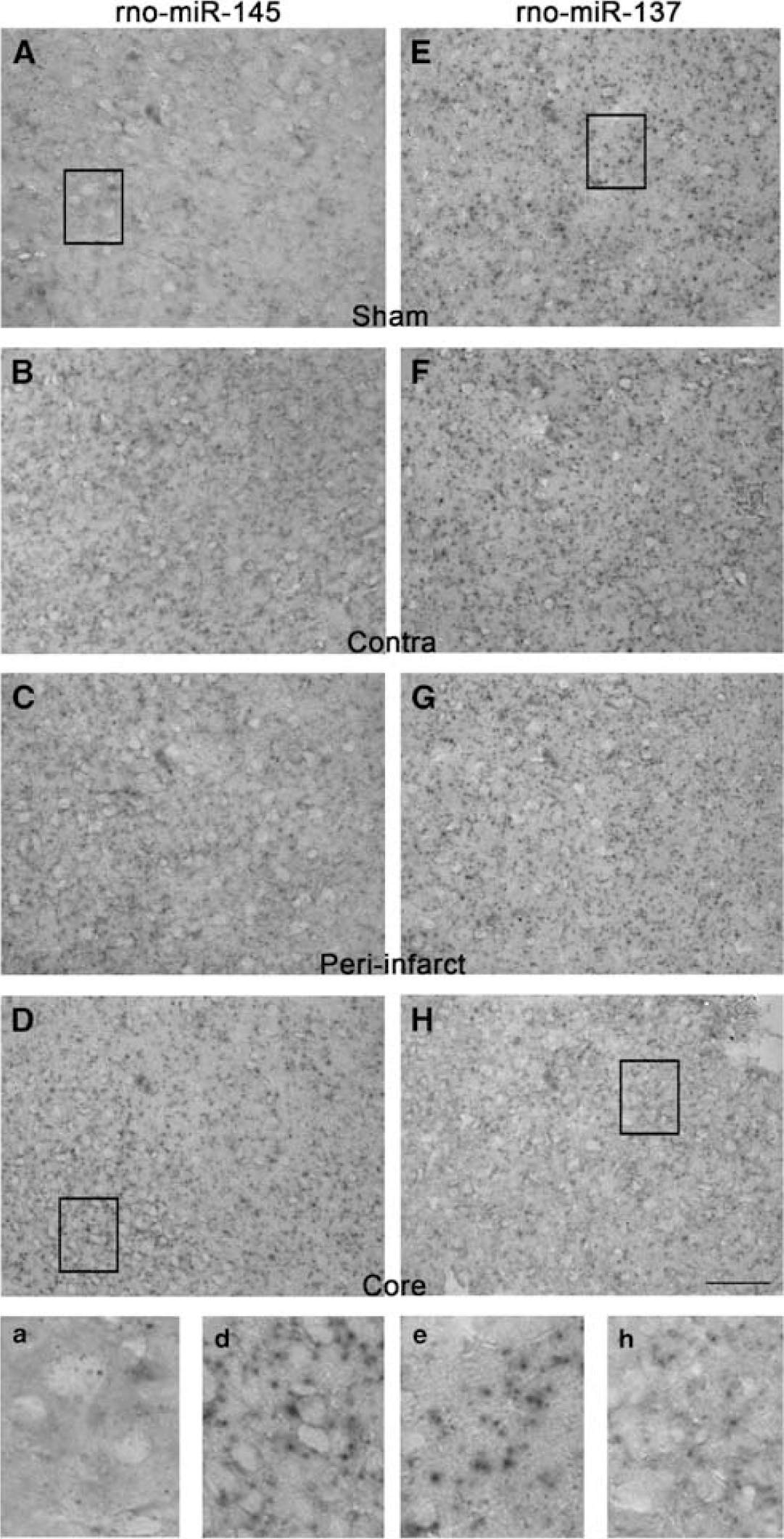
A cohort of rats subjected to transient MCAO or sham surgery were killed at 24 h of reperfusion (
MicroRNA Target identification
The miRNA targets were identified using the RegRNA (http://regrna.mbc.nctu.edu.tw) with a high-stringent score cutoff of ≥140 and minimum free energy cutoff of ≤–18.0. The RegRNA algorithm predicts targets based on a development of the open-source miRanda algorithm (http://www.microrna.org/miranda.html) (John et al, 2004) that incorporates the current biologic knowledge of target rules based on the mature rat miRNA sequences downloaded from mirBASE and the 5'-UTR regions of mRNAs from the NCBI database. An
Antagomir-145 Experiments
Mir-145 is one of the miRNAs upregulated significantly in the postischemic brain from 3 h to 3 days after transient MCAO. Bioinformatics search showed that SOD2 is a major target mRNA of mir-145. To prove this experimentally, we tested the effect of preventing postischemic mir-145 activity using an antagomir that targets mir-145. In brief, antagomir-145 (miRIDIAN Hairpin Inhibitor IH-320377-06-0010; Dharmacon, Chicago, IL, USA) or control antagomir (miRIDIAN Hairpin Inhibitor negative control IN-001005-01; Dharmacon) was dissolved in artificial CSF (aCSF; 119 mmol/L NaCl, 3.1 mmol/L KCl, 1.2 mmol/L CaCl2, 1 mmol/L MgSO4, 0.50 mmol/L KH2PO4, 25 mmol/L NaHCO3, 5 mmol/L D-glucose, 2.2 mmol/L urea, pH 7.4) at a concentration of 20 nmol/mL and continuously infused into the lateral ventricles of cohorts of rats (
Results
Temporal Progression of Neuronal Damage After Transient Middle Cerebral Artery Occlusion
A 1h transient MCAO in adult rats resulted in significant neuronal damage and infarction in the ipsilateral cortex and striatum that progressed with the time of reperfusion from 3 h to 3 days (Supplementary Figure 1). Microscopic examination of the Cresyl violet-stained brain sections showed a progressive loss of pyramidal neurons in the ipsilateral cortex between 6 h and 1 day of reperfusion (Supplementary Figure 1). These observations were similar to our previous studies (Satriotomo et al, 2006).
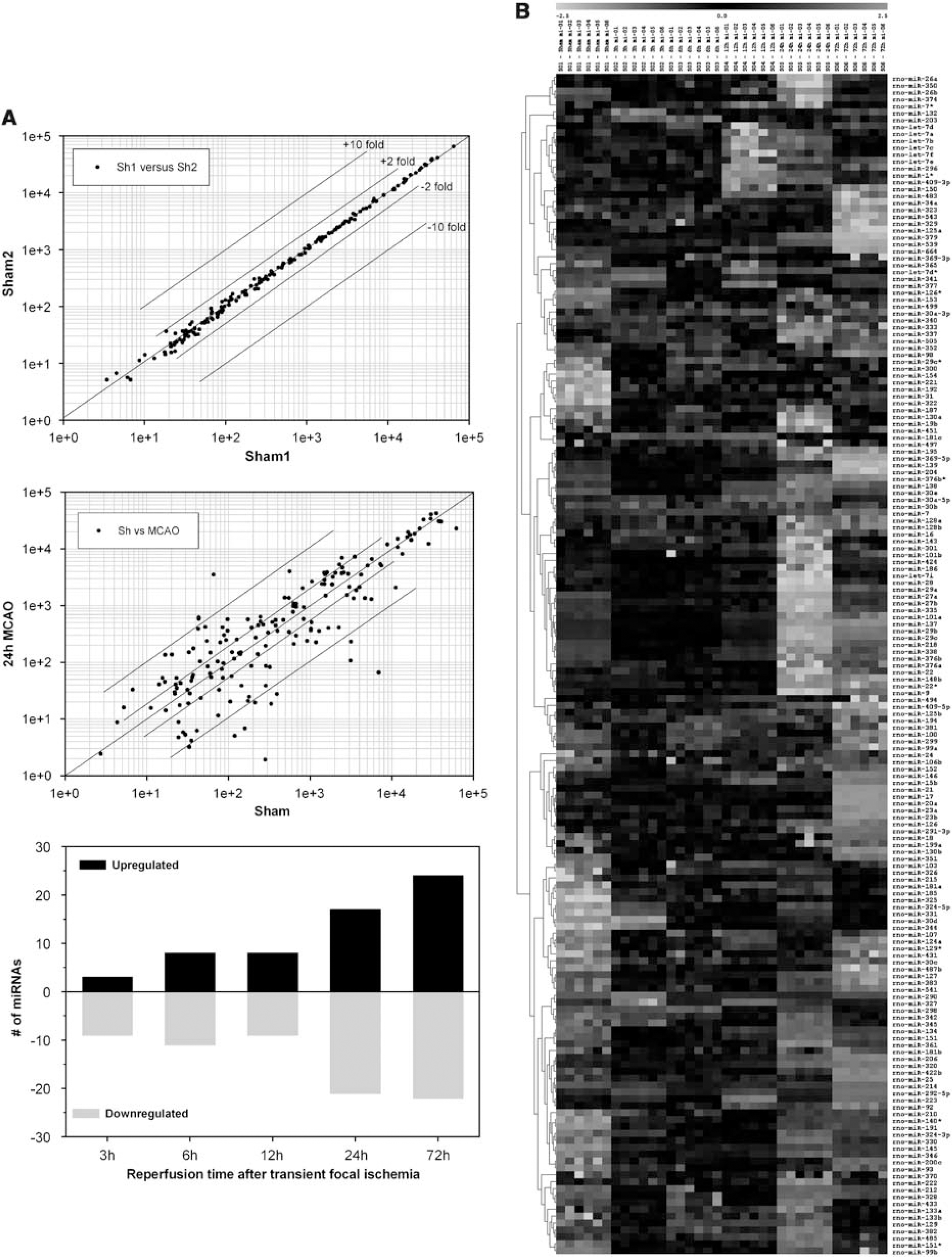
(
Focal Ischemia-Induced MicroRNA Changes
Of the 238 miRNAs spotted on the microarrays, 146 (61%) were observed to be expressed in the normal rat cerebral cortex (Supplementary Table 1). The expression levels for individual miRNAs ranged from 1 to 52,000 units (Figure 1A, top panel). After transient MCAO, 24 miRNAs showed increased expression and 23 miRNAs showed decreased expression at one or more reperfusion time points compared with sham (Table 1). Correlation plots of the expression levels of the miRNAs between sham samples showed no differences (Figure 1A, top panel), but correlation of sham samples with 24 h MCAO samples showed several up- and downregulated miRNAs (Figure 1A, middle panel). The miRNA expression altered rapidly after focal ischemia. Eleven miRNAs altered by 3 h of reperfusion (three increased and eight decreased), and this number gradually increased as the reperfusion time progressed (Figure 1A, bottom panel). A maximal number of 46 miRNA altered at 3 days reperfusion (24 increased and 22 decreased) (Figure 1A, bottom panel). Hierarchical clustering shows the expression levels of all the 238 miRNAs analyzed as a function of reperfusion time after transient MCAO in comparison to sham (Figure 1B). Eight miRNAs showed a significant change (three increased and five decreased) at all time points (3 h–3 days reperfusion) after focal ischemia (Table 1). Whereas, 11 miRNAs showed altered expression (5 increased and 6 decreased) at 4 of 5 time points evaluated (Table 1). Of the 47 miRNAs altered after focal ischemia, 16 showed > 10 fold change (6 increased and 10 decreased), and 11 showed 5 to 9.9-fold change (4 increased and 7 decreased) (Table 1). For a single miRNA, rno-mir-290 showed a maximal increase of 63-fold at 3 days of reperfusion (Table 1), and rno-mir-153 showed a maximal decrease of 52-fold at 1 day reperfusion (Table 1).
miRNAs increased in expression after transient MCAO
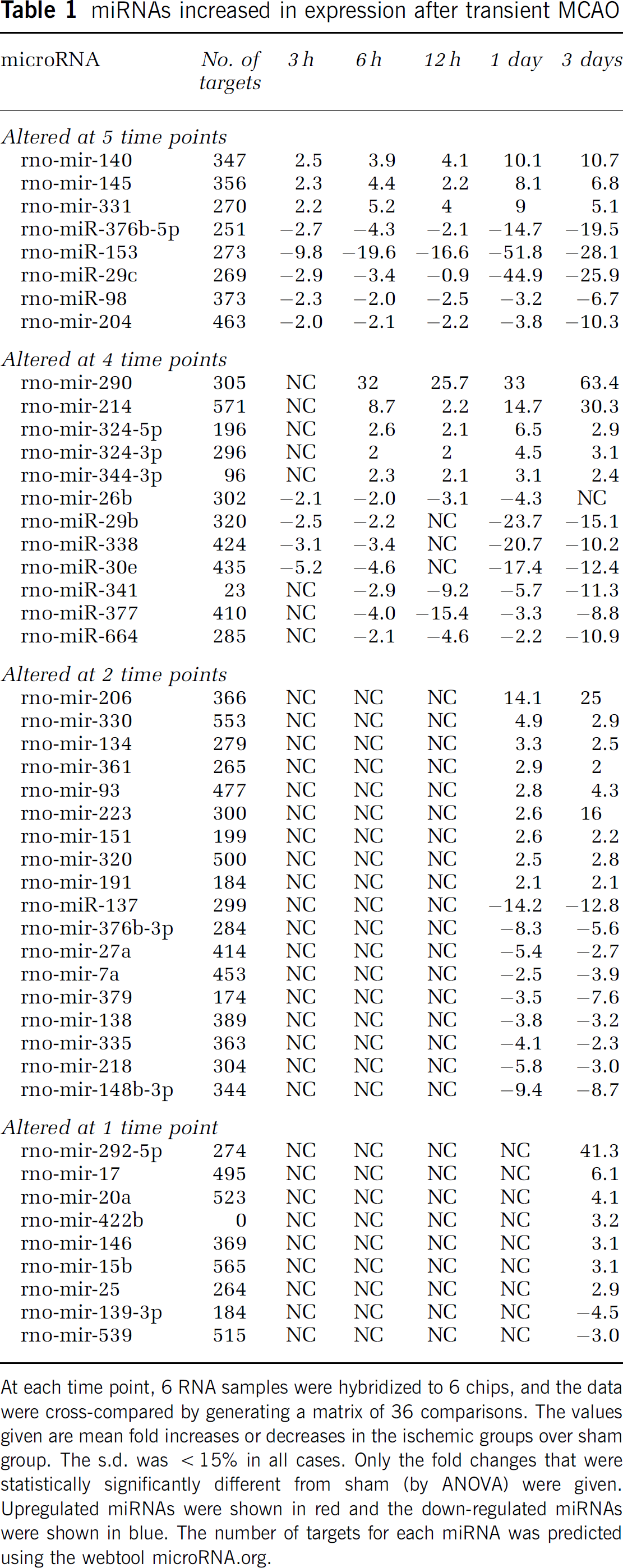
At each time point, 6 RNA samples were hybridized to 6 chips, and the data were cross-compared by generating a matrix of 36 comparisons. The values given are mean fold increases or decreases in the ischemic groups over sham group. The s.d. was >15% in all cases. Only the fold changes that were statistically significantly different from sham (by ANOVA) were given. Upregulated miRNAs were shown in red and the down-regulated miRNAs were shown in blue. The number of targets for each miRNA was predicted using the webtool microRNA.org.
Using
Using real-time PCR analysis, we evaluated the expression levels of three miRNAs that showed increased expression (mir-290, mir-145, and mir-206), three miRNAs that showed decreased expression (mir-137, mir-27a, and mir-218), and one miRNA that did not show any change in expression (let-7a) at 24 h reperfusion after transient MCAO compared with sham control. The real-time PCR essentially showed the same pattern of expression changes as observed with the microarray analysis (Figure 3A).
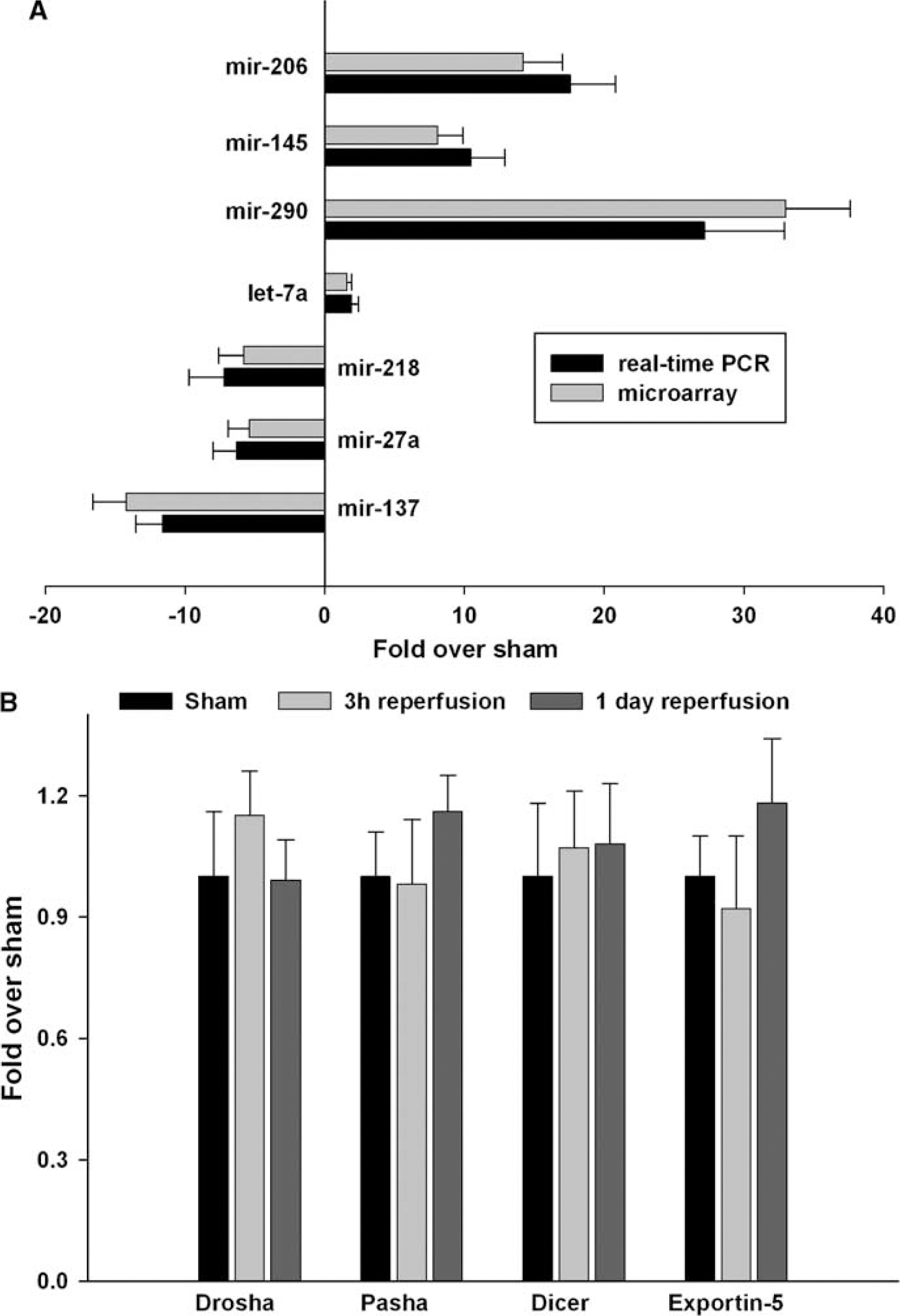
Real-time PCR analysis confirmed the changes in the levels of seven miRNAs at 1 day of reperfusion after transient MCAO observed by microarray analysis (
Focal Ischemia had no Effect on the MicroRNA Synthetic Machinery
The real-time PCR analysis showed no significant changes in the mRNA expression of the RNases Drosha and Dicer, the Drosha cofactor Pasha, and the pre-miRNA transporter exportin-5 at either 3 h or 24 h reperfusion after transient MCAO compared with sham controls (Figure 3B).
Common Targets of MicroRNA Altered After Focal Ischemia
Using the bioinformatics webtool microRNA.org, we searched the targets of each miRNA altered after focal ischemia. The 20 miRNAs altered in the postischemic brain (at least at 4 of the 5 reperfusion time points) showed several common mRNA targets. Surprisingly, many of those are known to be ischemia-responsive genes. In particular, we could correlate 14 inflammatory transcripts including IL1β, IL6, MIP1α, MCP1, ICAM1, Complement C3, COX2, and iNOS (Supplementary Figure 2) and 14 transcription factors including NF-kB, HIF1, Egr1, C/EBPβ, PPARγ, IRF1, STAT3, and ATF3 (Supplementary Figure 3) to multiple miRNAs altered after focal ischemia. In addition, bioinformatics search also correlated ischemia-sensitive miRNAs to several mRNAs that code for neuroprotective proteins (HSP70, HSP27, HO1, PTBR, MT-1/2, Mn-SOD, Catalase, IGF-BP3, and osteopontin) (Supplementary Figure 4), and ion channels and neurotransmitter receptors (VGAT, NCKX2, TWIK, Na Channel I and II, Na/K-ATPase, IP3 receptor, GluR-b, GluR-c, and adrenergic receptor A1d) (Supplementary Figure 5). All these mRNA transcripts were previously shown to be altered in the postischemic brain (Vemuganti et al, 2002).
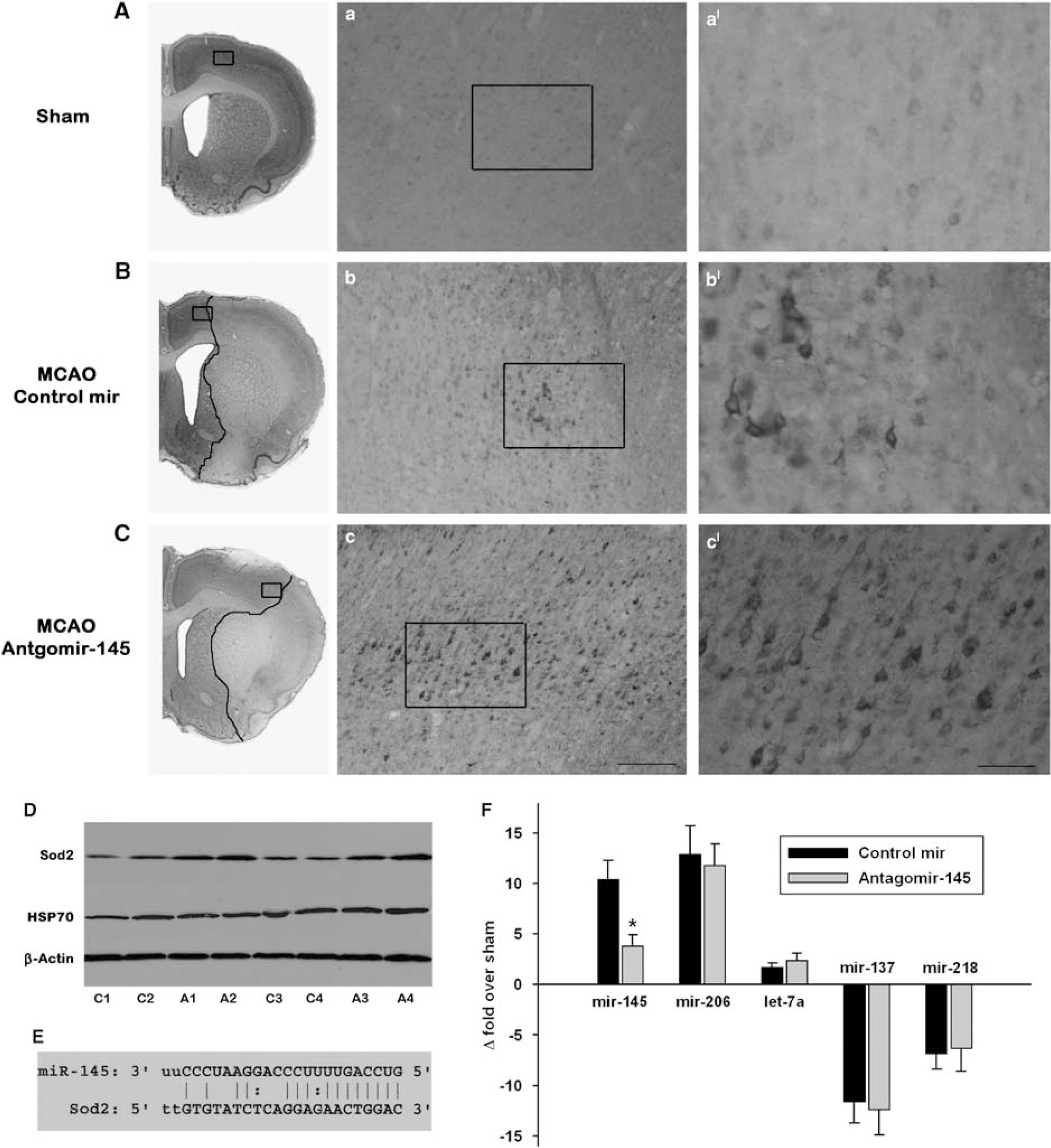
Antagomir-145 infusion resulted in increased SOD2 protein expression and decreased area of infarction at 1 day of reperfusion. Panels
Promoter Targets of Focal Ischemia-Induced MicroRNAs
Unique promoter signature motifs targeted by the stroke-induced miRNAs
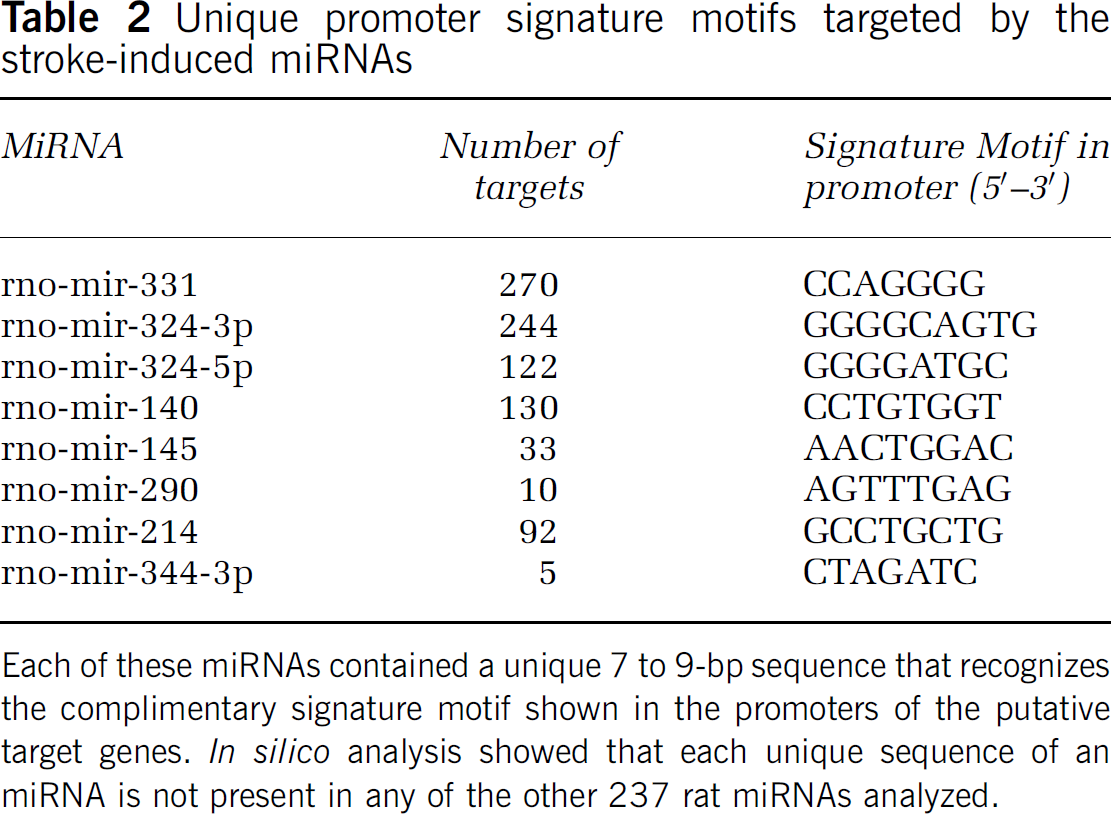
Each of these miRNAs contained a unique 7 to 9-bp sequence that recognizes the complimentary signature motif shown in the promoters of the putative target genes.
Some of the promoter targets of 3 ischemia-induced miRNAs
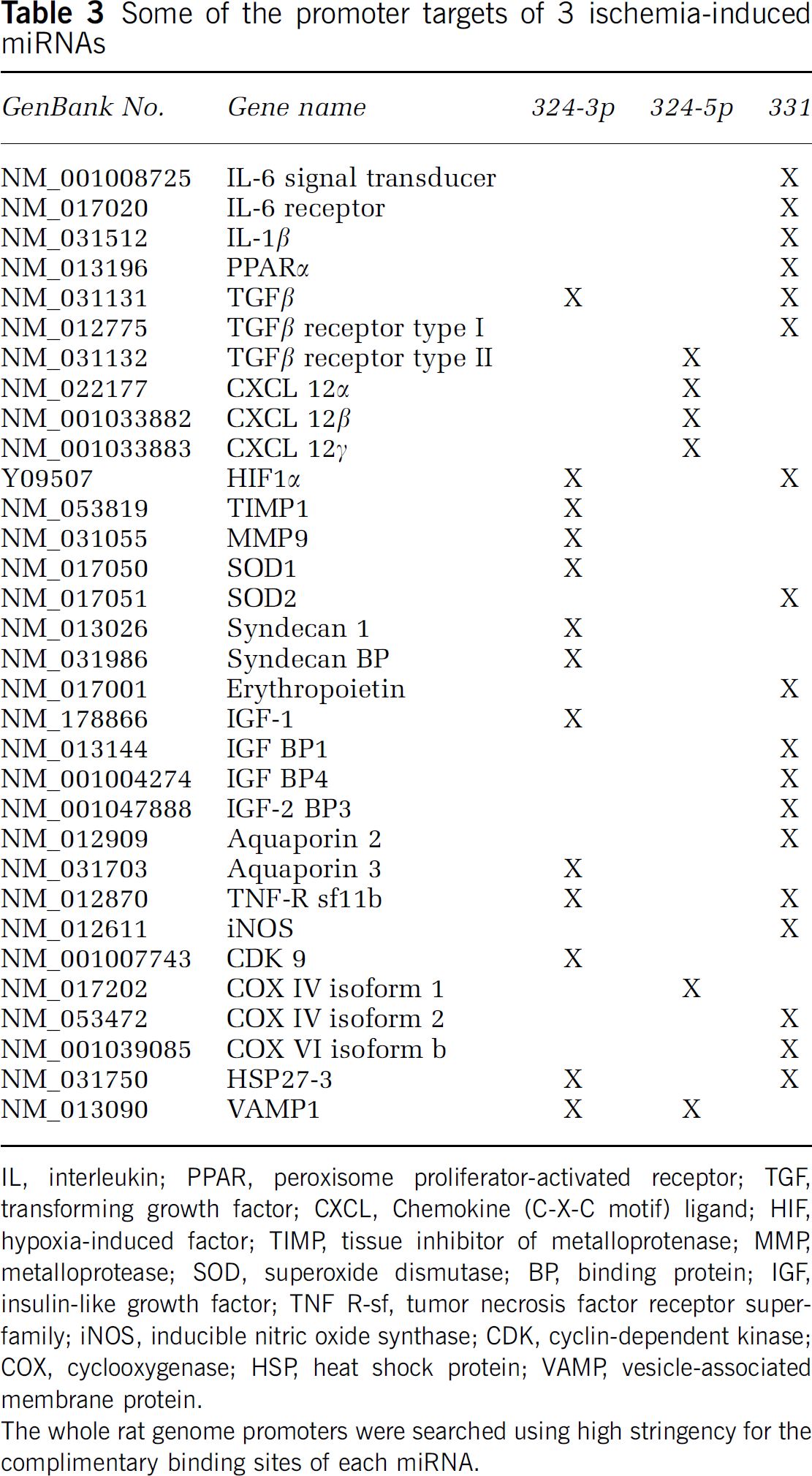
IL, interleukin; PPAR, peroxisome proliferator-activated receptor; TGF, transforming growth factor; CXCL, Chemokine (C-X-C motif) ligand; HIF, hypoxia-induced factor; TIMP, tissue inhibitor of metalloprotenase; MMP, metalloprotease; SOD, superoxide dismutase; BP, binding protein; IGF, insulin-like growth factor; TNF R-sf, tumor necrosis factor receptor superfamily; iNOS, inducible nitric oxide synthase; CDK, cyclin-dependent kinase; COX, cyclooxygenase; HSP, heat shock protein; VAMP, vesicle-associated membrane protein.
The whole rat genome promoters were searched using high stringency for the complimentary binding sites of each miRNA.
Mir-145 Controls Postischemic Superoxide Dismuate-2 Protein Expression
Mir-145 is one of the miRNAs that showed significant upregulation from 3 h to 3 days of reperfusion after transient MCAO (Table 1). Bioinformatics analysis with RegRNA and Miranda showed that rat miR-145 sequence has a complementary 8-bp targeting site in the 3'-UTR of rat SOD2 (GenBank# NM_017051) (Figure 4), which showed a very high minimum free energy of 165 and a score of −20.3. To experimentally show that mir-145 controls SOD2 protein expression in postischemic rat brain, we used an antagomir strategy. The cortex of sham-operated rats showed a very faint cytosolic immunostaining of SOD2 (Figure 4). In the control antagomir infused rats, the SOD2 immunoreactivity increased in the neurons in the periinfarct area, but not in the ischemic core in the ipsilateral cortex at 1 day of reperfusion after transient MCAO (Figure 4). Whereas, infusion of an antagomir-145 that targets mir-145, the area that showed SOD2 immunoreactivity was much bigger with more number of neurons showing intense SOD2 immunostaining (Figure 4). Antagomir-145 group also showed smaller cortical infarcts than the control antagomir group (by 24%;
Discussion
In brief, the results of the present study showed that transient focal ischemia induces an extensive temporal change in the cerebral miRNAome but has no effect on miRNA synthetic machinery. Bioinformatics analysis showed that many of the miRNAs altered in the postischemic brain can target several mRNAs previously demonstrated to be altered after stroke. Furthermore, the miRNAs altered after focal ischemia showed complementarity to several gene promoters.
The molecular mechanisms that promote neuronal death, and thus, neurologic dysfunction after stroke are not understood completely. We and others showed that focal ischemia leads to extensive changes in the cerebral mRNA expression in rodents (Soriano et al, 2000; Vemuganti et al, 2002; Lu et al, 2003; Sharp et al, 2006; Du et al, 2006; Tang et al, 2006; Kapadia et al, 2006; Yan et al, 2007). This altered expression of various families of genes plays an important role in promoting the postischemic pathologic mechanisms like inflammation, ionic imbalance, edema, and receptor dysfunction that precipitate neuronal death after focal ischemia. Many studies showed that pharmacological manipulations can induce neuroprotection after focal ischemia by affecting the gene expression (Wang et al, 2004; Xu et al, 2005; Chen et al, 2006; Tureyen et al, 2007).
In the adult mammalian brain, thousands of mRNAs continuously transcribe and translate to form various proteins. Transcription factors, DNA methylation, RNA polymerases, ribosomes, and other components of the transcriptional and translational machinery are known to control mRNA and protein expression under normal as well as pathologic states. In addition, recent studies showed that noncoding RNAs including miRNAs are potent regulators of gene and protein expression. In rats, 286 miRNAs are identified so far and these are thought to control ∼30% of all the mRNAs (http://microrna.sanger.ac.uk). As the action of an miRNA is dependent on the presence of a ∼8-bp signature sequence in the 3'-UTR regions of mRNAs, most miRNAs can bind to a huge number of mRNAs either sequentially or simultaneously. In addition, as the 3'-UTR of most mRNAs contains complementary binding sites for multiple miRNAs, a specific mRNA can be controlled by several miRNAs (structural studies estimated that most miRNAs can inhibit ∼200 mRNAs). Because of this redundancy in function, the small number of miRNAs can effectively control the huge number of mRNAs.
We currently observed that several miRNAs rapidly respond to focal ischemia and their expression changes by a very high magnitude. Furthermore, the ischemia-induced miRNA changes sustain for at least up to 3 days of reperfusion. We observed that 20% of the miRNAs analyzed (47 of the 238) were altered at one or more reperfusion time points after focal ischemia. This is striking as the number of mRNAs that show altered expression after focal ischemia is > 3% of those profiled (Soriano et al, 2000; Vemuganti et al, 2002; Lu et al, 2003; Sharp et al, 2006; Du et al, 2006; Tang et al, 2006; Kapadia et al, 2006; Yan et al, 2007). Thus, the effect of focal ischemia seems to be greater on the genes that transcribe miRNAs than those transcribe protein-coding RNAs. In vertebrates, many miRNA genes are located in the intragenetic regions and the introns of the coding regions (Wang et al, 2007). Although ischemia can independently influence the transcription of mRNAs and miRNAs, an altered miRNA can significantly influence the translation of its downstream mRNAs. A recent study showed altered miRNA expression at 1 and 2 days of reperfusion after transient MCAO in Sprague–Dawley rats (Jeyaseelan et al, 2008). Despite the fact that we used spontaneously hypertensive rats, many miRNAs observed to be up- or downregulated at 1 and 3 days of reperfusion in the present study were also reported to follow the same pattern of change in the Sprague–Dawley rats (Jeyaseelan et al, 2008). In addition, our studies also showed that many of these miRNAs change as early as 3 h and the altered expression persists up to 3 days. Another recent study showed altered miRNA levels in rat carotid arteries after angioplasty and demonstrated that preventing miR-21 upregulation had a significant negative effect on neointimal lesion formation (Ji et al, 1997).
Although, the mechanisms that regulate miRNA transcription after focal ischemia are not known, changes in the miRNA synthetic RNases (Dicer and Drosha) and 5-exportin is not responsible. Bioinformatics showed that many ischemic-responsive miRNAs target several mRNAs known to be altered after focal ischemia. This indicates a possible crosstalk between mRNAs and miRNAs at the level of transcriptional regulation. It is also known that certain mRNAs and/or their protein products can control the expression of specific miRNAs (Fazi et al, 2005). The complex nature of miRNA–mRNA interaction controls granulopoiesis by the reciprocal regulation of transcription factors NFI-A and C/EBPα and miRNA mir-223 (Fazi et al, 2005). Although NFI-A maintains mir-223 at low levels, replacing NFI-A by C/EBPα upregulates mir-223 which in turn suppresses NFI-A promoting granulocyte differentiation (Fazi et al, 2005). If the levels of a specific mRNAs are increased, that mRNA can instruct the expression of an upstream miRNA to prevent the translation of the mRNA as a feed-forward mechanism (Taganov et al, 2006). In addition, recent studies showed that miRNAs can also bind to promoter regions of certain genes to induce gene expression by a mechanism known as RNA-induced gene activation (Place et al, 2008). Thus, miRNAs and mRNAs can control each other bidirectionally, and hence, the relationship between an miRNA and an mRNA is not a simple, inverse one. Thus, after focal ischemia, an increased miRNA could prevent the translation of a downstream mRNA (by binding to the 3'UTRs) or could increase the expression of the mRNA transcription (by binding to the gene promoter). However, if an miRNA is decreased in expression, it can allow a downstream mRNA to translate or fail to sustain a gene transcription (if the binding of that miRNA to the promoter is a prerequisite for the normal level of transcription of that mRNA). Hence, altered miRNA expression has several consequences on mRNA transcription as well as translation.
Bioinformatics analysis plays a significant role in miRNA target prediction. However, to develop an miRNA as a therapeutic target, it is essential to experimentally validate its relation to the expression of the target protein. Mir-145 was one of the miRNAs observed presently to be upregulated after transient MCAO. Bioinformatics analysis with RegRNA and Miranda showed SOD2 as a likely target of mir-145 with very high stringency. As SOD2 is known to play a role in the antioxidant defense, its induction might promote cell survival after ischemia. We observed that after transient MCAO, the neurons that survived in the periinfarct area of the ipsilateral cortex show SOD2 protein expression, but mir-145 induction might not be allowing SOD2 protein to the necessary levels increase to promote any neuroprotection. When the postischemic mir-145 was neutralized with antagomir-145, the number of neurons that showed SOD2 was much higher in the ipsilateral cortex as well as the periinfarct area was much larger. Although these studies indicate a relationship between mir-145 and its target SOD2 in the postischemic brain, the protection afforded by antagomir-145 was only marginal as other miRNAs and mRNAs might also play a role in the postischemic neuronal damage.
As multiple miRNAs acting on common targets is an effective way to alter mRNA function, we evaluated the targets of some miRNAs altered after focal ischemia. In particular, we tested the 20 miRNAs that showed persistent change (altered expression at 4–5 consecutive reperfusion time points between 3 h and 3 days). We used the high-stringency bioinformatics tool microRNA.org that allows finding targets common to multiple miRNAs. It is striking that 14 miRNAs in various combinations target mRNAs that code for inflammatory genes, transcription factors, HSPs, anti-oxidant enzymes, growth factors, ion channels, and neurotransmitter receptors that are known to be altered after focal ischemia (Vemuganti et al, 2002). This indicates that altered expression of specific miRNAs and mRNAs after focal ischemia is not completely random. However, it is essential to experimentally establish the role of miRNA changes in modulating mRNAs that control various pathologic mechanisms in the postischemic brain.
To understand the possibility of RNA-induced gene activation in the ischemic brain, we conducted an
Thus, the results of the present study indicate that focal ischemia significantly alters the temporal expression of many miRNAs, which might be controlling the mRNA transcription and translation, and thus, the resulting stroke pathophysiology. Future studies will show if modulating specific miRNAs can be a therapeutic option to prevent postischemic pathophysiological events and/or to promote plasticity and regeneration.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.