Automated Yeast Mating Protocol Using Open Reading Frames from Saccharomyces cerevisiae Genome to Improve Yeast Strains for Cellulosic Ethanol Production
Engineering the industrial ethanologen Saccharomyces cerevisiae to use pentose sugars from lignocellulosic biomass is critical for commercializing cellulosic fuel ethanol production. Approaches to engineer pentose-fermenting yeasts have required expression of additional genes. We implemented a high-throughput strategy to improve anaerobic growth on xylose and rate of ethanol production by evaluating overexpression of each native S. cerevisiae gene from a collection of haploid PJ69–4 MATa strains expressing the gene open reading frames (ORFs) mated to a haploid PJ69–4 MATalpha strain expressing the Piromyces sp.E2 xylose isomerase (XI) gene. The resulting 6113 diploid strains containing the XI gene and a different yeast gene ORF were screened for growth on xylose in anaerobic plate cultures using an integrated robotic workcell. Nine unique strains were isolated; two were found to no longer grow on glucose; seven were further evaluated for fermentation of alkaline peroxide pretreated enzymatically saccharified wheat straw hydrolysate. All successfully used glucose and xylose, consuming most of the glucose and a small amount of the xylose. Transforming the strains with an additional vector expressing xylulokinase gene did not improve anaerobic growth on xylose but improved glucose use and ethanol production on the hydrolysate, with three strains giving maximum ethanol production ≥ 14.0 g L−1.
Over 95% of U.S. fuel ethanol is produced using corn. Eventually, it is envisioned that annual corn ethanol production can expand to 12–15 billion gallons, consuming 31% or more of the corn harvest. For this reason, commercializing lignocellulose as a feedstock for further ethanol production has been made a national priority. Despite a growing commitment by industry to move toward these more challenging feedstocks, technical barriers still remain. One critical need is for robust microbial strains capable of fermenting the more diverse mixture of neutral sugars released by the hydrolysis of lignocellulose. Plant cell-wall lignocellulose contains, in decreasing order of relative importance, glucose, xylose, arabinose, ga-lactose, and various other sugars. Saccharomyces strains are capable of fermenting hexoses; however, they do not ferment the pentose sugars, arabinose, or xylose.
Several yeast, such as Pachysolen tannophilus, Pichia stipitis, and Candida shehatae, naturally ferment xylose. Although some of these are being pursued for commercialization, they have several drawbacks, including the inability to grow anaerobically on xylose, low tolerance to acetic acid and other inhibitory chemicals common to biomass hydrolysates, and generally low productivity and yields compared to glucose-fermenting Saccharomyces cerevisiae. Attention in recent years has turned to engineering bacteria to selectively produce ethanol,1 improving the performance of native xylose-fermenting yeast,2 or engineering Saccharomyces strains to ferment pentose sugars, especially xylose.3–7
Saccharomyces yeast can naturally use the pentose phosphate pathway intermediate xylulose. Engineering strategies to enable Saccharomyces to ferment xylose have centered on introducing the needed activities for converting xylose to xylulose. Naturally xylose-fermenting yeasts convert xylose into xylitol using xylose reductase (XR) and xylitol into xylulose using xylitol dehydrogenase (XDH), but the process gives rise to a cofactor imbalance that results in the production of xylitol.46Saccharomyces yeast strains have previously been engineered to express XR and XDH genes,8 and several have reasonable ethanol yields and reduced xylitol production, conceivably because enough oxygen enters the system to regenerate nicotinamide adenine dinucleotide (NAD+) from NADH via respiration instead of xylitol pro-duction.45 Precisely controlled oxygen levels are nearly impossible to maintain in large-scale industrial operations, which limit the industrial potential of these biocatalysts.
In an effort to convert xylose to xylulose without creating cofactor imbalances, Saccharomyces yeast strains were engineered to express a heterologous xylose isomer-ase (XI), which catalyzes this conversion directly.59 However, the activity of the XI enzyme was too low for efficient xylose metabolism. It was discovered that the XI from Piromyces sp.E2 can be expressed at sufficient levels in S. cerevisiae.610 After evolutionary engineering and expression of all genes for the enzymes involved in the conversion of xylose into intermediates of glycolysis in addition to expression of XI and deletion of the gene-encoding aldose reductase, a Saccharomyces strain was constructed that had an ethanol production rate of 0.46 g per gram xylose per hour under anaerobic batch cultivation on xylose. When grown on 20 g per liter glucose and xylose each, an exponential glucose consumption phase followed by a slower, almost linear, xylose consumption phase was observed.7 Further selection for xylose growth yielded a strain that fermented all sugars in 24 h when cultivated in anaerobic batch culture with 20 g per liter each of glucose and xylose, an improvement of 20 h over the time required before strain selection. On xylose alone, it had an ethanol production rate of 0.49 g per gram xylose per hour under anaerobic batch cultivation.4 Anaerobic growth in xylose cultures is a highly desirable quality in industrial fermentation because it reflects cell viability and increases rate of ethanol production. Although uptake kinetics were also improved, the engineered Saccharomyces strains are only now moving toward commercialization.4 Co-fermentation of hexose and pentose sugars is still a major challenge.3
It is significant that engineering approaches to improve pentose-fermenting yeasts have required expression of auxiliary genes to complement activity of XI. Despite evidence that overexpression of further genes is required, no systematic screening of the yeast genome has been undertaken to identify the genes that need to be overexpressed for improved xylose fermentation. The goal of this work was to evaluate overexpression of each S. cerevisiae gene in a strain also expressing XI and determine which of the genes, if any, confer the ability for anaerobic growth on xylose. These genes would be appropriate targets for further improving the fermentation characteristics of xylose-fermenting Saccharomyces strains. The present study used a collection of S. cerevisiae gene open reading frames (ORFs) in pOAD LEU selectable vectors driven by an alcohol dehydrogenase (ADH) promoter in the PJ69–4 MATa S. cerevisiae strain. Each of these was mated to the haploid PJ69–4 MATalpha S. cerevisiae strain containing the Piromyces sp.E2 XI gene expressed from a pDEST32 TRP selectable vector with an ADH promoter. The resulting diploids were selected for anaerobic growth on xylose medium. The effect of xylulokinase (XKS) activity on ethanol production was also evaluated by transforming the diploid strains containing the XI gene and each of the S. cerevisiae gene ORFs with pSUMOduoRGStetHisXKS URA selectable vector. To mate and screen the entire collection of gene ORFs, an automated high-throughput strategy incorporating the essential features of the conventional manual process was developed and implemented on an integrated robotic workcell.1113
Materials and Methods
Strains
Two haploid S. cerevisiae strains were used.14 They are identical except for mating type:
Production of pDEST32-XI TRP Selectable Haploid PJ69–4 MATalpha Strain
The pDEST32-XI TRP selectable plasmid was constructed by replacing the LEU2 gene in the pDEST32 LEU2 selectable bait plasmid commercially available in the ProQuest Two-Hybrid System kit (Invitrogen, Carlsbad, CA) with the TRP1 gene. The TRP1 marker was PCR amplified from pRS404 using the forward primer, leu2::TRP1 (S): 5′CATTTCAGCAATATATATATATATATTTCAAGGATATACCATTCTAATGTCTGTTATTAATTTCACAGGT3′ and the reverse primer, leu2::TRP1 (AS): 5′TTTCATTTATAAAGTTTATGTACAAATATCATAAAAAAAGAGAATCTTTCTATTTCTTAGCATTTTTGA3 to generate 40 nucleotides of flanking homology to the 5′ and 3′ ends of the LEU2 ORF fused to the TRP1 marker (5′leu2::TRP1::3′leu2 fragment). Yeast cells (InvSc1) were transformed with 500 ng of the 5′leu2::TRP1::3′leu2 fragment and 25 ng of linearized pDEST32 with LEU marker using a standard lithium acetate procedure. Trp+ Leu-isolates were identified and the plasmid from these strains was rescued into Escherichia coli. This pDEST32 plasmid has the LEU2 marker deleted and replaced with the TRP1 marker. Plasmid preparation was performed and the plasmid rescued into the PJ69–4 MATalpha haploid yeast strain. The Piromyces XI gene was cloned into the resulting TRP selectable modified bait plasmid using standard Gateway recombination procedures with LR clonase, as described previously.11 The XI gene was obtained using forward primer 5′ATGGCTAAGGAATATTTCCCAC3′ and reverse primer 5′CGAAGCTATTGTTGCCATGTACCAATAA3′ to amplify the XI coding region from the pENTR D TOPO-XI vector constructed previously.15 The amplified fragment was isolated using GENECLEAN II PCR kit (MP Biomedicals, Solon, OH) according to the manufacturer's directions and ligated into the pDEST32 TRP selectable vector using LR clonase II (Invitrogen). The resulting ligations were transformed into TOP10 competent cells (Invitrogen) according to manufacture's instructions and grown overnight at 37 °C. Colonies were picked into 1.6 mL LB AMP 50 medium (Teknova, Hollister, CA). Plasmids were isolated from the cultures using Qiagen (Germantown, MD) plasmid preparation kit. The resulting yeast expression plasmid pDEST32-XI TRP selectable was verified by sequencing and transformed into PJ69–4 MATalpha using EZ yeast clone procedures. Yeast cells (3–4-mm size colonies) were inoculated into YPD +ADE (Teknova) (25 mL) using a sterile loop and incubated for 2 days at 30 °C with shaking at 150 rpm. One milliliter of culture was pelleted at 1300 rpm in a microfuge (Thermo Electron, Waltham, MA) and 125 μL of EZ-Transformation solution, 2 μg of plasmid DNA, and 5-μL carrier DNA were added. The cells were resuspended by vortexing at moderate speed for 1 min, incubated at 30 °C for 30 min, and transferred to CM glucose TRP selective medium plates, spread with a sterile spreader, and incubated at 30 °C until transformants were observed (typically 3 days).
Production of Haploid PJ69–4 MATa Strain Containing Collection of pOAD Plasmids with ORFs from S. cerevisiae Genome
A collection of PCR-generated ORFs predicted from the S. cerevisiae genome in pOAD LEU selectable vectors transformed into PJ69–4 MATa haploid yeast strain and spotted in a 384-grid pattern on 16 86 × 128-mm Omnitrays containing CM glucose –LEU + ADE medium was provided by Dr. Stanley Fields at University of Washington, Seattle, WA.1617
Automated Mating of PJ69–4 MATalpha Haploid Strain Containing pDEST32-XI TRP Selectable Vector to a PJ69–4 MATa Haploid Strain Containing ORFs from S. cerevisiae Genome in pOAD LEU Selectable Vectors
The 384-grid colony plate containing the PJ69–4 MATa haploid strains with the S. cerevisiae gene ORFs was placed on the liquid handler deck and used to inoculate 50-μL CM 2.0% glucose –LEU + ADE liquid medium (Teknova, Hollister, CA) in 16 86 × 128-mm Omnitray agar plates (VWR International, Westchester, PA) with 384 × 5-μL spot colonies representing the yeast gene collection for 2-day culture. A 384-well Matrix deep-well plate (Fisher Scientific, Waltham, MA) containing 50-μL CM glucose –LEU + ADE medium was inoculated with each of the pOAD-gene ORFs in haploid PJ69–4 MATa yeast strains and grown for 2 days at 30 °C. The plate was sealed and brought to the liquid handler for mating with the PJ69–4 MATalpha strain containing pDEST32-XI TRP selectable vector to produce diploid PJ69–4 yeast strains each containing two plasmids.
A sterile trough containing 150 mL of PJ69–4 MATalpha haploid yeast with pDEST32-XI TRP selectable vector in CM glucose –TRP + ADE (Teknova) grown for 2 days at 30 °C was placed onto the liquid handler deck. Mating was performed in a Matrix 384-well deep-well plate containing 50 μL of YPD +ADE (Teknova) seeded with 30 μL of PJ69–4 MATa cultures with pOAD-gene ORF LEU selectable plasmids and 20 μL of MATalpha PJ69–4 yeast strain with pDEST32-XI TRP selectable plasmid. Cultures were grown for 1 day at 30 °C in YPD +ADE medium.
Sealed Matrix 384-well deep-well 1-day culture plates containing mated diploid PJ69–4 strains with pOAD-gene ORF plasmids and pDEST32-XI plasmid were pierced by the sterile stainless steel pipet tips of the liquid handler, and 20 μL were diluted in a new 384-well deep-well plate containing 100-μL CM glucose –LEU-TRP + ADE liquid medium and grown for 10 h at 30 °C. The 384-well deep-well dilution cultures in CM glucose –LEU-TRP + ADE medium were broadcast from the 384-well format into 4 × 96-well ABgene pyramid bottom deep-well plates containing the same medium, and grown at 30 °C for 2 days.
Sealed ABgene 96-well deep-well cultures containing the diploid PJ69–4 yeast strain with pDEST32-XI TRP selectable plasmid and pOAD LEU selectable plasmids with the yeast gene ORFs were pierced by the sterile stainless steel pipet tips of the liquid handler, and 20 μL were diluted in a new 384-well deep-well plate containing 100-μL CM glucose –LEU-TRP + ADE liquid medium. Three microliters of the dilution cultures were spotted in 96-well format onto CM xylose –LEU-TRP + ADE medium on 86 × 128-mm Omnitray agar plates and incubated in a fully anaerobic chamber (Becton Dickinson, Franklin Lakes, NJ) for 6 days at 30 °C.
Ten colonies that grew out on xylose medium after 6 days in an anaerobic chamber were picked into CM glucose –TRP-LEU + ADE medium to make glycerol stock. Two of the 10 strains did not grow out on glucose medium; those that did were replated onto CM xylose –LEU-TRP + ADE plates grown anaerobically again, and these colonies were used for growth and ethanol production analyses.
Construction of RGStetHisXKS Plasmid and Production of Triple-Plasmid Diploid Strain
The RGStetHis-tagged XKS plasmid was constructed by using the forward XKS primer with RGSHHHH tag, Oligo 1 (88-mer; GC content: 46/88), BBCHTS 7–2–08A:
and reverse XKS primer, Oligo 2 (27-mer; GC content: 12/27), BBCHTS 7–2–08B:
5′CTACTCACACAATGGCAGCAATTGTTG3′
to add the RGStetHis tag to the XKS sequence in the pENTR D TOPO-XKS plasmid. The RGStetHisXKS gene was then Gateway cloned into the pSUMOduo URA selectable vector.1315 The resulting vector was used to transform TOP10 competent cells, and plasmids were produced as described previously.11 The resulting pSUMOduo-RGStetHisXKS URA selectable plasmid was transformed into the double-plasmid diploid strains to give the triple-plasmid diploid strains.
Anaerobic Growth of Multiplasmid Strains on Single Sugars (Glucose or Xylose)
Plates from the liquid handler containing multiplasmid diploid yeast strains were placed in an incubator (Becton Dickinson) at 30 °C with anaerobic conditions maintained using BD GasPak EZ Gas Generating Container Systems with Indicator (Becton Dickinson). For growth on glucose, samples were streaked on CM glucose selective plates. For growth on xylose, similar plates were used but CM xylose was substituted for CM glucose. For anaerobic growth, the plates were first placed in an anaerobic chamber for at least 2 h and then samples were applied. The plates were wrapped and placed back into the anaerobic chamber for 6 days.
Analysis of Ethanol from Single Sugar (Glucose and Xylose) Anaerobic Cultures
Plate cultures were analyzed in triplicate for ethanol production using gas chromatography (6890N; Agilent Technologies, Wilmington, DE) with a packed column as described previously.18 Before injection into the GC, the samples were diluted fourfold with distilled water. The GC was equipped with an autosampler and an integrator. Ethanol concentration was calculated as gL-1. Plate cultures were analyzed in triplicate for xylose and glucose concentrations using Surveyor high-pressure liquid chromatography (HPLC) equipped with an automatic sampler/injector (Thermo Electron Corporation, West Palm Beach, FL). The HPLC column (Aminex HPX-87P; Resin based) was obtained from BioRad (Hercules, CA). Solvent (MilliQ water) flow rate was maintained at 0.6 mL min-1. For sugar analysis, the mixture was centrifuged at 15,000 × g for 15 min followed by 20-fold dilution and injection into the HPLC.18
Wheat Straw Alkaline Hydrolysate Production
Wheat straw was purchased from a local farmer, dried in a forced-air oven at 55 °C for 24 h, and milled in a hammer mill to pass through a 1.27 mm screen. Milled wheat straw was slurried in water (8.6%, w/v) containing H2O2 (2.25%, v/v), adjusted to pH 11.5 using NaOH, and shaken in an incubator at 250 rpm at 35 °C for 24 h. The pretreated wheat straw was adjusted to pH 5.0 using concentrated HCl and saccharified at 45 °C for 72 h using commercial enzymes as described previously.19 Activity levels of these enzymes were reported in a previous paper.20
Analysis of Hydrolysates for Cell Growth and Concentrations of Glucose and Xylose
Samples (1 mL) were taken after 5 and 8 days growth in the hydrolysate at 30 °C with shaking at 100 rpm. The flask was closed with a rubber stopper and covered with porous tape to maintain sterility. Glucose, xylose, and ethanol were analyzed by HPLC.19 Cell growth of the yeast strains was monitored by measuring the optical density of the appropriately diluted culture sample at 660 nm.
Results and Discussion
Production and Screening of Anaerobic Multiplasmid Diploid Yeast Strains
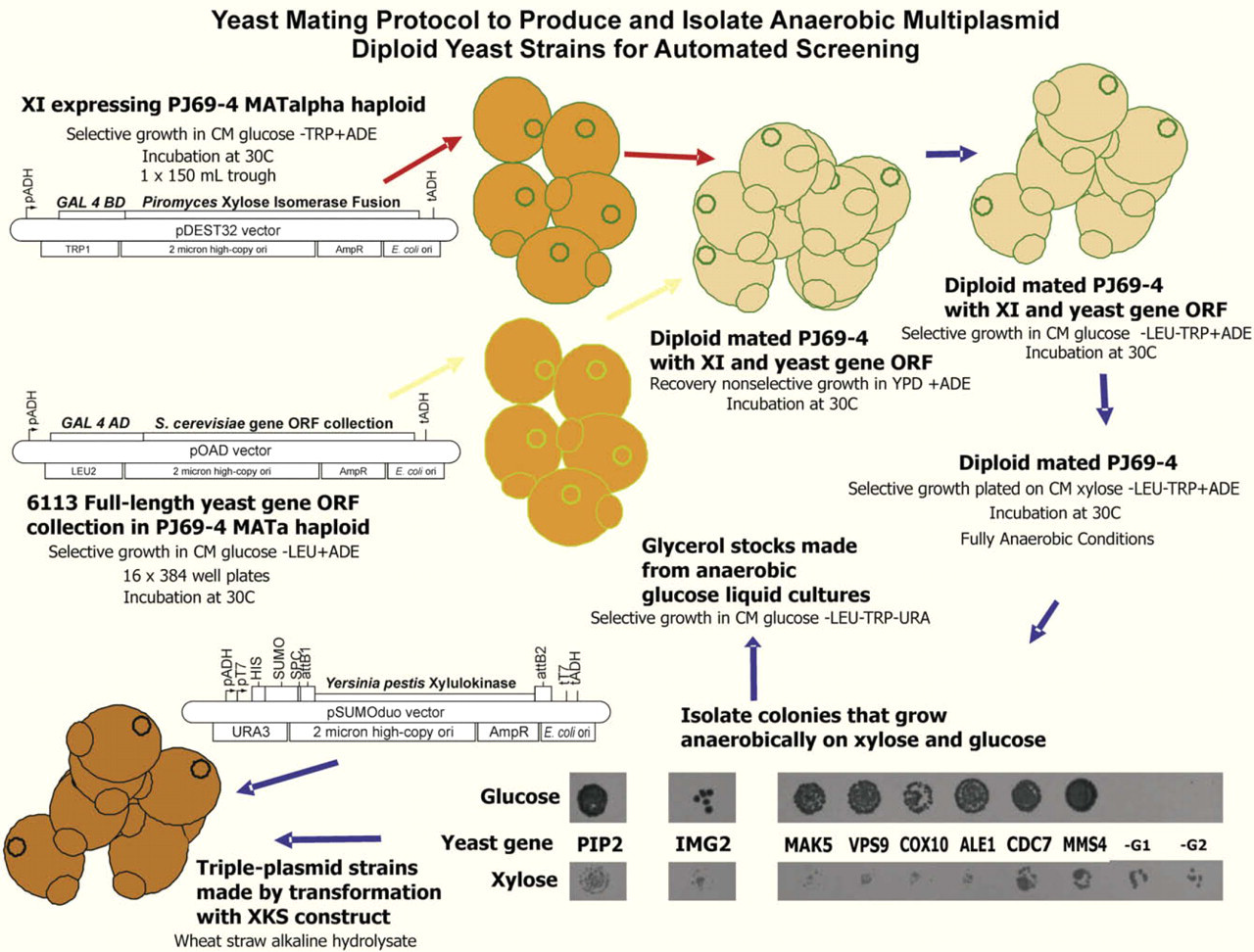
A strategy was successfully developed for production and screening of multiplasmid diploid yeast strains (Fig. 1). This strategy was used to mate a haploid PJ69–4 MATalpha strain expressing the Piromyces sp.E2 XI ORF to haploid PJ69–4 MATa strains containing a collection of S. cerevisiae gene ORFs. A total of 10 diploid S. cerevisiae PJ69–4 yeast strains were obtained (9 were unique) that grew anaerobically on xylose selective medium. Previously, we had isolated S. cerevisiae INVSc1 strains capable of only aerobic growth on xylose.21 The selected PJ69–4 strains were analyzed and found to contain the following genes in the pOAD LEU selectable plasmid: PIP2, IMG2, MAK5, VPS9, COX10, ALE1, CDC7, MMS4, –G1, and –G2 (latter two were not identified because they could no longer grow on glucose). Strains containing the first eight of these genes retained the ability to grow anaerobically on glucose (PIP2 and CDC7 have the same sequence). In addition to the plasmid containing one of the genes, these diploid PJ69–4 yeast strains contained the Piromyces sp. XI ORF in the pDEST32 TRP selectable plasmid. The three-plasmid diploid PJ69–4 yeast strains were additionally transformed with the pSUMOduo-RGStetHisXKS URA selectable plasmid. The entire production and screening process was carried out on the integrated robotic platform described previously.1112 The steps and materials used are summarized in Figure 2. The two-plasmid and three-plasmid strains with the seven unique genes that grew anaerobically on xylose and glucose were analyzed for growth rate and ethanol production to identify the strains that demonstrated optimum anaerobic growth on xylose and glucose combined with high-ethanol production from wheat straw alkaline hydrolysate.
Schematic of automated yeast mating protocol to produce and isolate yeast strains capable of anaerobic growth on xylose and glucose.
Outline of the automated protocol steps on the robotic platform showing plates required for mating the yeast PJ69–4 MATalpha haploid strain, expressing Piromyces XI gene from plasmid pDEST32, with the PJ69–4 MATa haploid strain, expressing one of the yeast genes from plasmid pOAD.
Growth of Multiplasmid Diploid PJ69–4 Yeast Strains
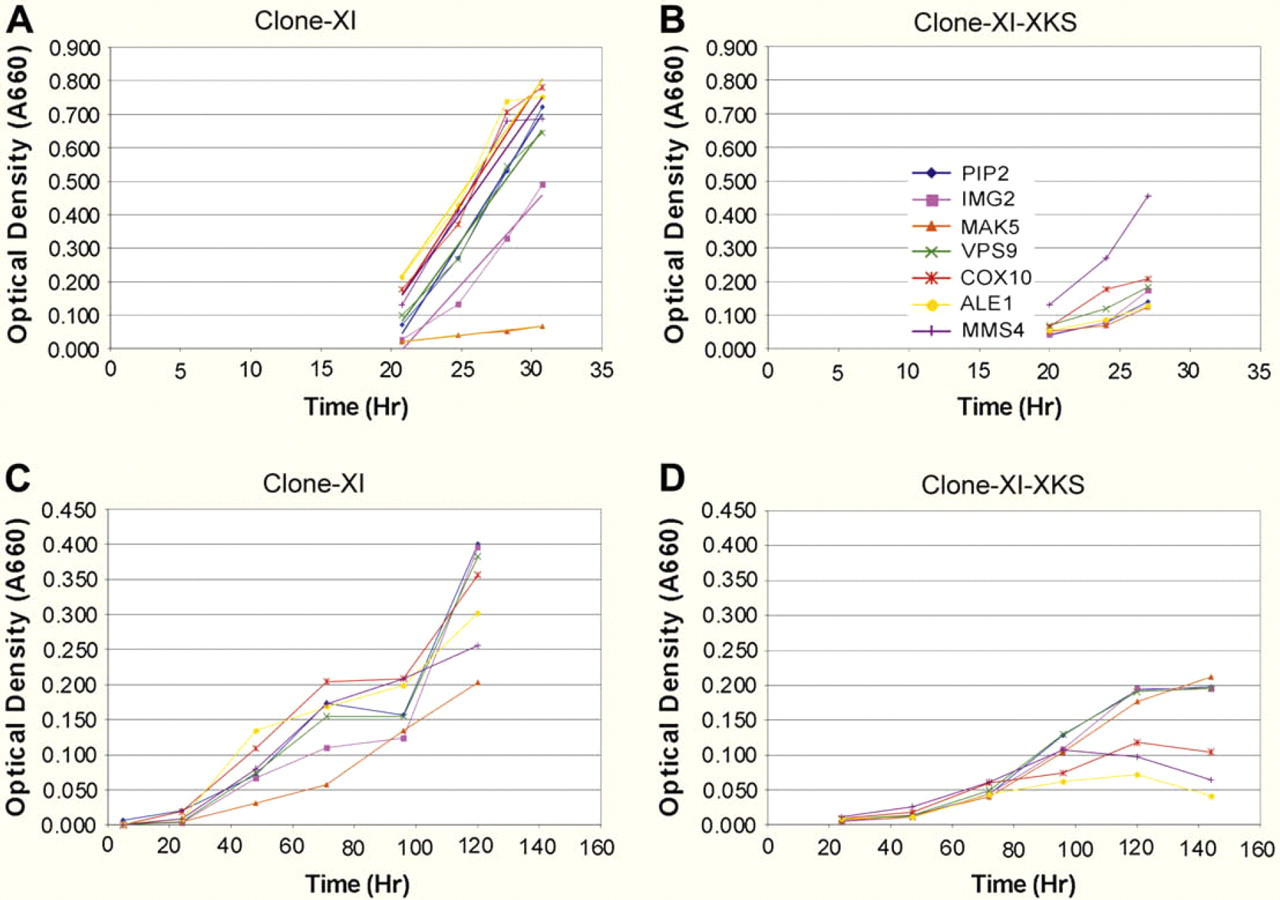
Growth curves of the two-plasmid diploid strain (XI gene) obtained from mating of the PJ69–4 MATalpha strain containing Piromyces XI to the PJ69–4 MATa strain containing one of the yeast gene ORFs with or without expression of XKS grown anaerobically on glucose (Fig. 3A, B) or xylose (Fig. 3C, D) indicate, as expected, that all the strains grew more slowly on xylose than glucose. It is also clear from comparing the double-plasmid growth curves to the triple-plasmid growth curves on either glucose or xylose medium that addition of XKS did not improve the growth rate.
Growth curves based on optical density at 660 nm of diploid strain cultures with and without expression of XKS. Double-plasmid strains were derived from mating PJ69–4 MATalpha strain expressing Piromyces XI with PJ69–4 MATa strain expressing one of the yeast genes. Triple-plasmid strains were produced by transformation of double-plasmid strains with pSUMOduo-RGStetHisXKS URA selectable plasmid. Strains were grown in anaerobic culture at 30 °C with 100 rpm shaking in CM 2% glucose TRP LEU selective medium or in CM 2% xylose TRP LEU selective medium. (A) Strains without XKS in glucose; (B) Strains with XKS in glucose; (C) Strains without XKS in xylose; (D) Strains with XKS in xylose. Note: x Axis in xylose graphs covers longer timeframe than in glucose graphs and y axis covers larger OD range in glucose graphs than in xylose graphs.
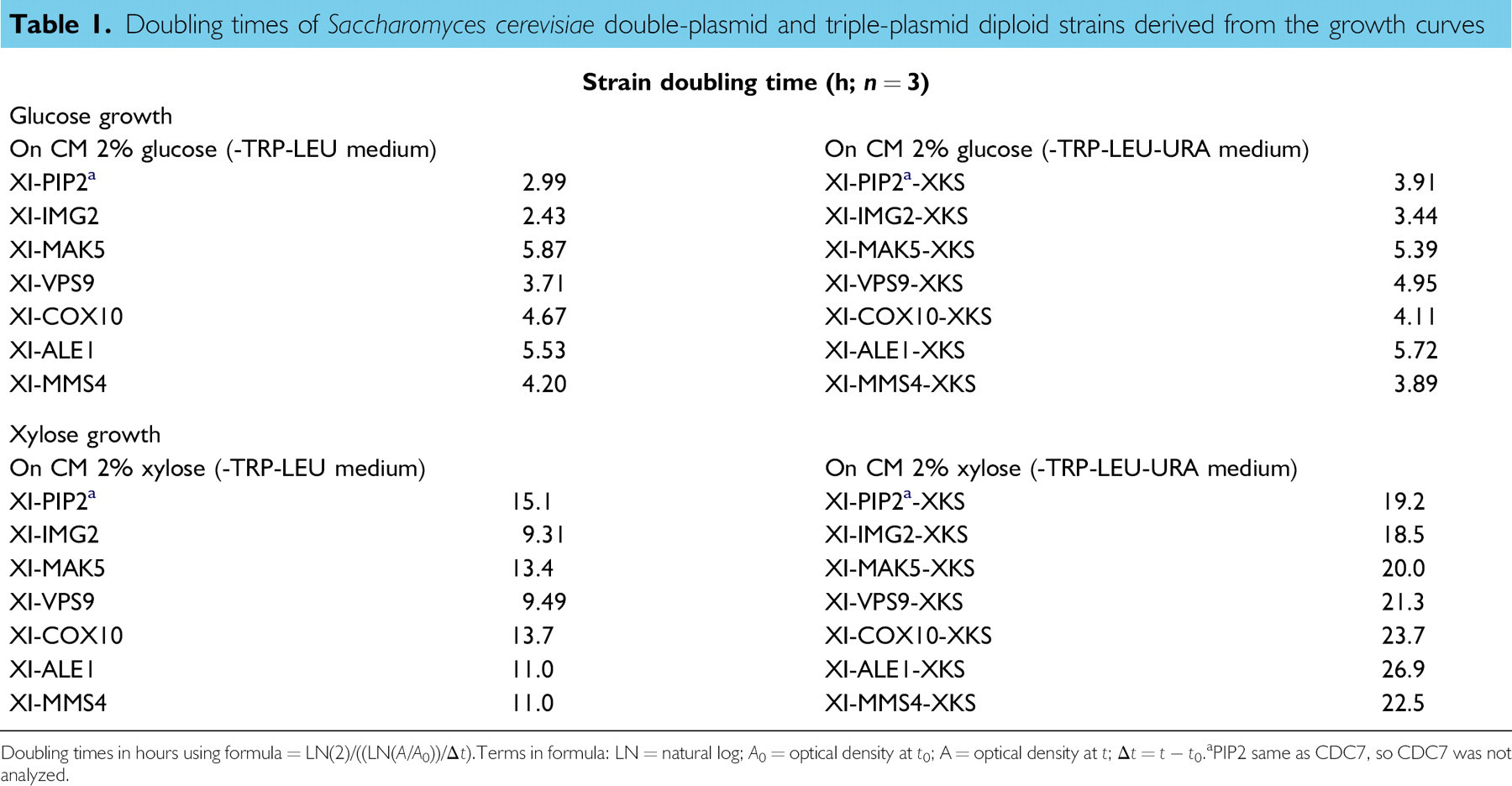
Doubling times derived from the growth curves for anaerobic growth in glucose or xylose medium of S. cerevisiae double-plasmid and triple-plasmid diploid strains are presented in Table 1. In general, doubling times were much longer for all strains in xylose medium than in glucose medium and for triple-plasmid strains in xylose medium than for double-plasmid strains in xylose medium. For specific gene ORFs, the double-plasmid strains expressing PIP2 or IMG2 demonstrated the shortest doubling times on glucose medium. Similarly, the shortest doubling times on glucose medium for the triple-plasmid strains again included the strains expressing PIP2 or IMG2, with the doubling time for the strain expressing MMS4 essentially the same as for PIP2. Among the triple-plasmid strains grown in xylose medium, the strains expressing PIP2 or IMG2 also had the shortest doubling times (although much longer than in glucose medium). However, among the double-plasmid strains grown in xylose medium, the strain expressing PIP2 had the longest doubling time, whereas the strains expressing IMG2 or VPS9 had the shortest doubling times.
Doubling times of Saccharomyces cerevisiae double-plasmid and triple-plasmid diploid strains derived from the growth curves
Doubling times in hours using formula = LN(2)/((LN(A/A0))/Δt). Terms in formula: LN = natural log; A0 = optical density at t0; A = optical density at t; Δt = t – t0.
PIP2 same as CDC7, so CDC7 was not analyzed.
Ethanol Productivity of Multiplasmid Diploid PJ69–4 Yeast Strains Grown in Glucose
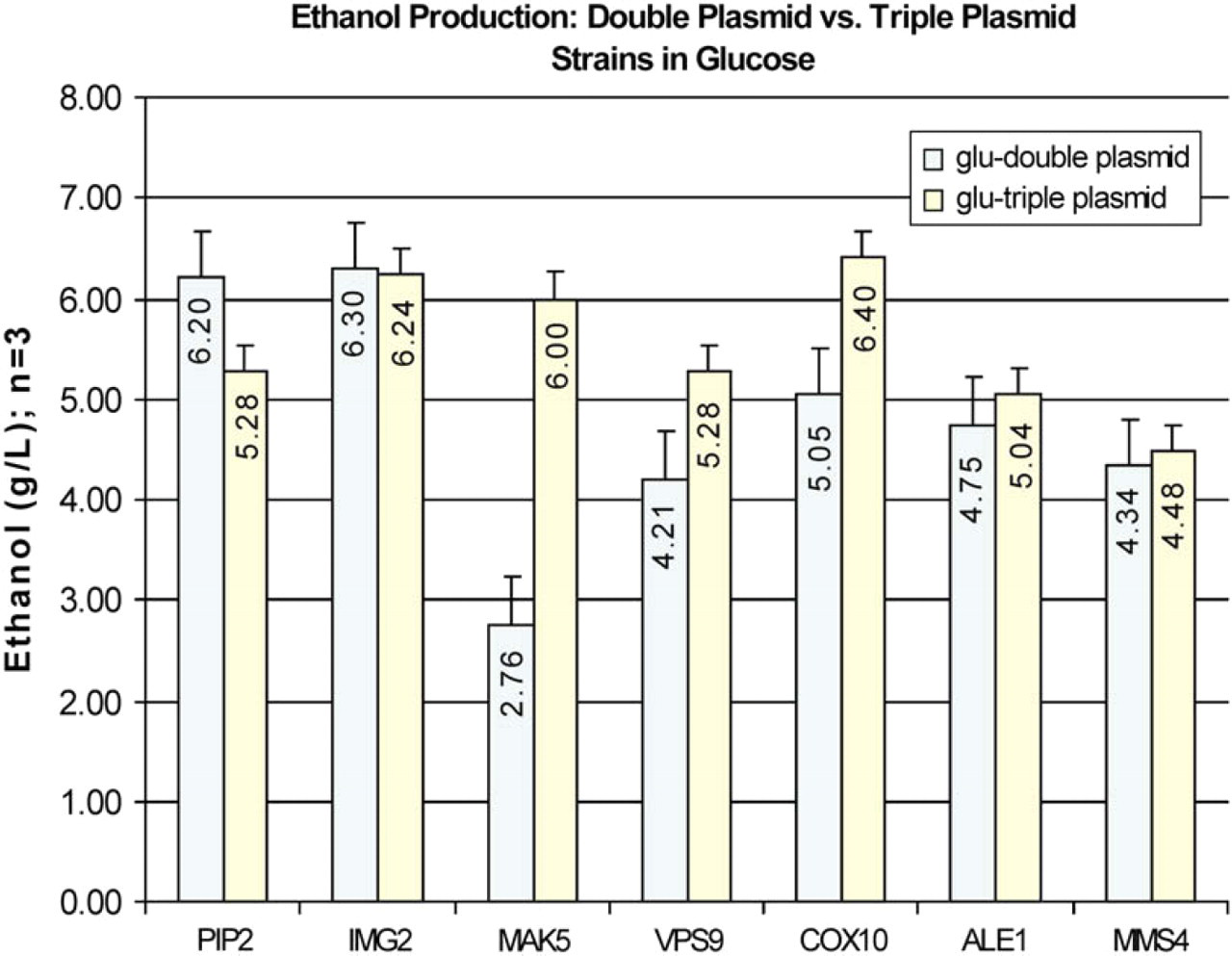
A comparison of ethanol production from double-plasmid and triple-plasmid strains grown in glucose medium shows that the addition of XKS did not greatly increase ethanol production in the triple-plasmid strains with the exception of the strain expressing MAK5 mainly because the ethanol production in the two-plasmid MAK5 strain was much lower than in the other two-plasmid strains (Fig. 4). No ethanol was obtained from the double-plasmid or triple-plasmid strains grown in xylose alone. The double-plasmid strains expressing PIP2 and IMG2 and the triple-plasmid strains expressing IMG2 and COX10 gave the highest ethanol concentration in glucose.
Ethanol production after 100 h incubation from double-plasmid and triple-plasmid strains in glucose selective medium. No ethanol was obtained from the double-plasmid or triple-plasmid strains grown in xylose selective medium.
Ethanol Production, Glucose and Xylose Usage, and Optical Density of Multiplasmid Diploid PJ69–4 Yeast Strains Grown on Wheat Straw Alkaline Hydrolysate
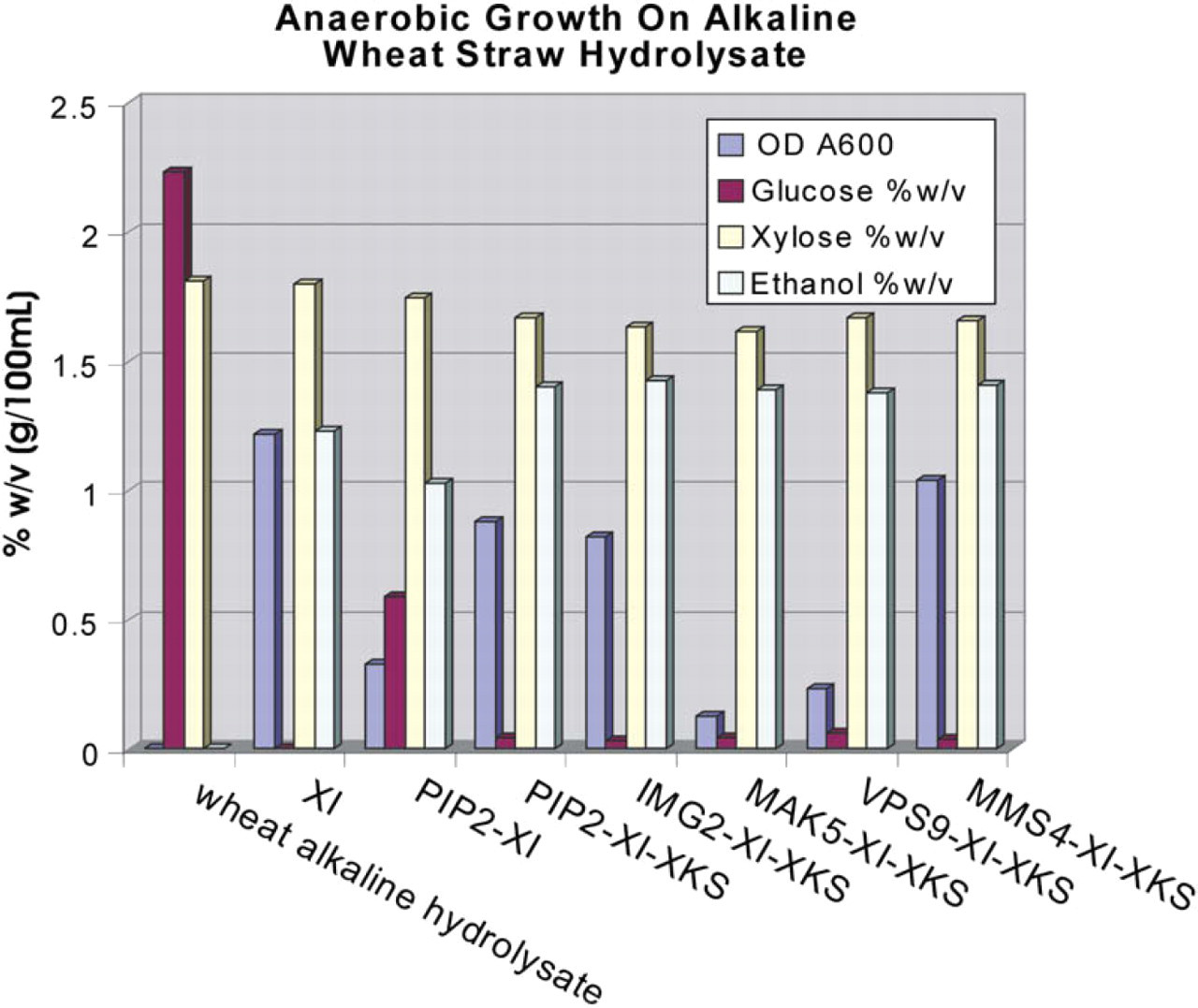
Maximum amounts of ethanol produced after 100 h reaction from double-plasmid (XI-PIP2) and triple-plasmid (XI-Genes PIP2, IMG2, MAK5, VPS9, or MMS4-XKS, respectively) diploid PJ69–4 yeast strains that gave optimum anaerobic growth on alkaline hydrolysate are shown in Figure 5, in addition to the xylose and glucose levels and optical density at 600 nm. When grown on wheat straw alkaline hydrolysate, the highest ethanol production, best glucose and xylose usage, and greatest optical density occurred in triple-plasmid strains expressing PIP2, IMG2, and MMS4. The triple-plasmid strains expressing MAK5 and VPS9 gave almost as much xylose and glucose use and ethanol production; however, the optical density was dramatically lower. The reason for this result is not clear. The single-plasmid control strain expressing the XI gene (no yeast gene or XKS gene) used glucose but not xylose and produced less ethanol than the triple-plasmid strains. On wheat straw alkaline hydrolysate, all double- and triple-plasmid strains analyzed first used glucose with production of ethanol, followed by slower consumption of xylose accompanied by further ethanol production from the remainder of the glucose (data not shown).
Maximum amounts of ethanol from double-plasmid and triple-plasmid diploid PJ69–4 yeast strains that gave optimum growth on alkaline hydrolysate, and the xylose and glucose levels after 100-h incubation in 25 mL hydrolysate. Optical density at 600 nm is also shown. Control is Saccharomyces cerevisiae strain engineered to express XI only (no yeast gene or XKS).
Description of Gene ORF and Maximum Ethanol Production
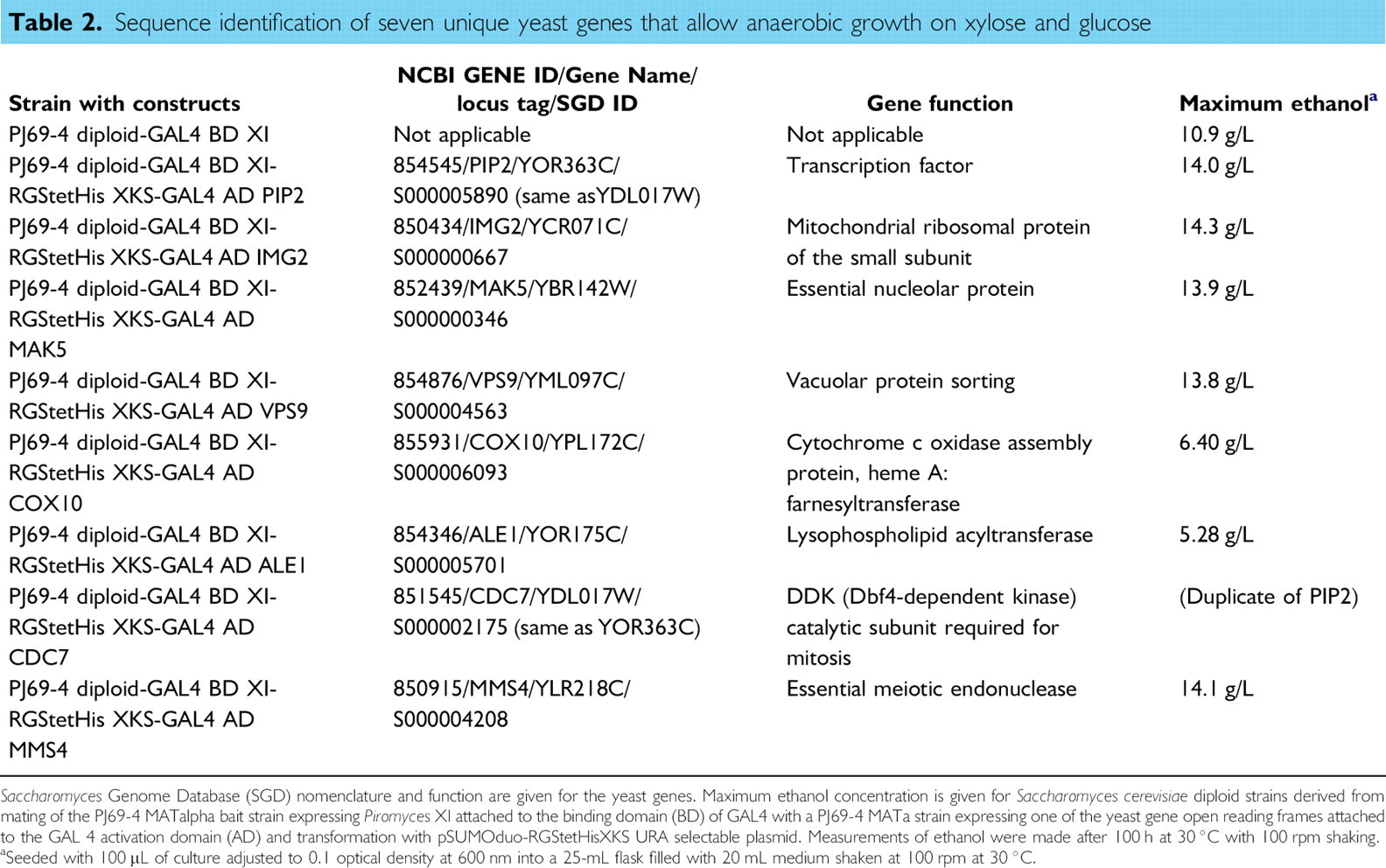
Sequence identification and function of the seven unique gene ORFs that allow anaerobic growth on xylose and maximum ethanol production for triple-plasmid S. cerevisiae strains are presented in Table 2. Those triple-plasmid strains expressing PIP2, IMG2, and MMS4 gave the highest ethanol concentration at 14.0, 14.3, and 14.12 g/L, respectively.
Sequence identification of seven unique yeast genes that allow anaerobic growth on xylose and glucose
Mitochondrial ribosomal protein of the small subunit
14.3 g/L
RGStetHis XKS-GAL4 AD IMG2
PJ69–4 diploid-GAL4 BD XI-
852439/MAK5/YBR142W/S000000346
Essential nucleolar protein
13.9 g/L
RGStetHis XKS-GAL4 AD MAK5
PJ69–4 diploid-GAL4 BD XI-
854876/VPS9/YML097C/S000004563
Vacuolar protein sorting
13.8 g/L
RGStetHis XKS-GAL4 AD VPS9
PJ69–4 diploid-GAL4 BD XI-
855931/COX10/YPL172C/S000006093
Cytochrome c oxidase assembly protein, heme A: farnesyltransferase
6.40 g/L
RGStetHis XKS-GAL4 AD COX10
PJ69–4 diploid-GAL4 BD XI-
854346/ALE1/YOR175C/S000005701
Lysophospholipid acyltransferase
5.28 g/L
RGStetHis XKS-GAL4 AD ALE1
PJ69–4 diploid-GAL4 BD XI-
851545/CDC7/YDL017W/S000002175 (same as YOR363C)
DDK (Dbf4-dependent kinase) catalytic subunit required for mitosis
(Duplicate of PIP2)
RGStetHis XKS-GAL4 AD CDC7
PJ69–4 diploid-GAL4 BD XI-
850915/MMS4/YLR218C/S000004208
Essential meiotic endonuclease
14.1 g/L
RGStetHis XKS-GAL4 AD MMS4
Saccharomyces Genome Database (SGD) nomenclature and function are given for the yeast genes. Maximum ethanol concentration is given for Saccharomyces cerevisiae diploid strains derived from mating of the PJ69–4 MATalpha bait strain expressing Piromyces XI attached to the binding domain (BD) of GAL4 with a PJ69–4 MATa strain expressing one of the yeast gene open reading frames attached to the GAL 4 activation domain (AD) and transformation with pSUMOduo-RGStetHisXKS URA selectable plasmid. Measurements of ethanol were made after 100 h at 30 °C with 100 rpm shaking.
Seeded with 100 μL of culture adjusted to 0.1 optical density at 600 nm into a 25-mL flask filled with 20 mL medium shaken at 100 rpm at 30 °C.
Conclusion
Nine unique yeast genes were identified that allowed growth on xylose plates in an anaerobic chamber of the double-plasmid diploid S. cerevisiae strains expressing the yeast gene and XI. Seven of these strains were still able to grow anaerobically in glucose liquid culture. Additionally, it was found that transforming the double-plasmid strains with a plasmid expressing XKS did not aid anaerobic growth of these strains in xylose. No ethanol production occurred in xylose alone. The double-plasmid strains expressing PIP2 and IMG2 and the triple-plasmid strains expressing IMG2 and COX10 gave the highest ethanol concentration when grown anaerobically in glucose. When grown on wheat straw alkaline hydrolysate, the highest ethanol production, best glucose and xylose usage, and greatest optical density occurred in triple-plasmid strains expressing PIP2, IMG2, and MMS4. The triple-plasmid strains expressing MAK5 and VPS9 demonstrated almost as much xylose and glucose consumption and ethanol production; however, the optical density was dramatically lower. It is not intuitively obvious how the added yeast genes function to improve anaerobic growth and ethanol production in these strains. Further investigation is warranted to address this question and also to determine if combinations of these genes will provide further improvement.
Acknowledgments
We thank Karen Hughes for critical reading and formatting of the manuscript. We also thank Nathane Orwig for performing the sequencing work. The gift of the pOAD plasmid library from Stanley Fields at the University of Washington, Seattle, WA is greatly appreciated. This work was supported by the National Center for Research Resources of the National Institutes of Health by a grant to Trisha N. Davis entitled “Comprehensive Biology Exploiting the Yeast Genome,” P41 RR11823.
References
1.
DienB. S.CottaM. A.JeffriesT. W.Bacteria engineered for fuel ethanol production: Current status. Appl. Microbiol. Biotechnol.2003, 63(3), 258–266.
Van MarisA. J. A.AbbottD. A.BellissimiE.van den BrinkJ.KuyperM.LuttikM. A. H.WisselinkH. W.ScheffersW. A.van DijkenJ. P.PronkJ. T.Alcoholic fermentation of carbon sources in biomass hydrolysates by Saccharomyces cerevisiae: current status. Antonie van Leeuwenhoek2006, 90(4), 391–418.
5.
KarhumaaK.Hahn-HägerdalB.Gorwa-GrauslandM. F.Investigation of limiting metabolic steps in the utilization of xylose by recombinant Saccharomyces cerevisiae using metabolic engineering. Yeast2005, 22, 359–368.
6.
KuyperM.WinklerA. A.van DijkenJ. P.PronkJ. T.Minimal metabolic engineering of Saccharomyces cerevisiae for efficient anaerobic xylose fermentation: a proof of principle. FEMS Yeast Res.2004, 4, 655–664.
7.
KuyperM.ToirkensM. J.DiderichJ. A.WinklerA. A.van DijkenJ. P.PronkJ. T.Evolutionary engineering of mixed-sugar utilization by a xylose-fermenting Saccharomyces cerevisiae strain. FEMS Yeast Res.2005, 5, 925–934.
8.
JeffriesT. W.JinY. S.Metabolic engineering for improved fermentation of pentoses by yeasts. Appl. Microbiol. Biotechnol.2004, 63, 495–509.
9.
WalfridssonM.BaoX.AnderlundM.LiliusG.BulowL.Hahn-HägerdalB.Ethanolic fermentation of xylose with Saccharomyces cerevisiae harboring the Thermus thermophilus xylA gene, which expresses an active xylose (glucose) isomerase. Appl. Environ. Microbiol.1996, 62, 4648–4651.
10.
HarhangiH. R.AkhmanovaA. S.EmmensR.van der DriftC.de LaatW. T.van DijkenJ. P.JettenM. S.PronkJ. T.Op den CampH. J.Xylose metabolism in the anaerobic fungus Piromyces sp. E2 follows the bacterial pathway. Arch. Microbiol.2003, 180, 134–141.
11.
HughesS. R.RiedmullerS. B.MertensJ. A.LiX.-L.BischoffK. M.CottaM. A.FarrellyP. J.Development of a liquid handler component for a plasmid-based functional proteomic robotic workcell. J. Assoc. Lab. Autom.2005, 10(5), 287–300.
12.
HughesS. R.RiedmullerS. B.MertensJ. A.LiX.-L.BischoffK. M.QureshiN.CottaM. A.FarrellyP. J.High-throughput screening of cellulase F mutants from multiplexed plasmid sets using an automated plate assay on a functional proteomic robotic workcell. Proteome Sci.2006, 4, 10.
13.
HughesS. R.DowdP. F.HectorR. E.RiedmullerS. B.BartolettS.MertensJ. A.QureshiN.LiuS.BischoffK. M.LiX.-L.JacksonJ. S.Jr.SternerD.PanavasT.RichJ. O.FarrellyP. J.ButtT. R.CottaM. A.Cost-effective high-throughput fully automated construction of a multiplex library of mutagenized open reading frames for an insecticidal peptide using a plasmid-based functional proteomic robotic workcell with improved vacuum system. J. Assoc. Lab. Autom.2007, 12(4), 202–212.
14.
DreesB. L.SundinB.BrazeauE.CavistonJ. P.ChenG.-C.GuoW.KozminskiK. G.LauM. W.MoskowJ. J.TongA.SchenkmanL. R.McKenzieA.IIIBrennwaldP.LongtineM.BiE.ChanC.NovickP.BooneC.PringleJ. R.DavisT. N.FieldsS.DrubinD. G.A protein interaction map for cell polarity development. J. Cell Biol.2001, 154(3), 549–576.
15.
HughesS. R.DowdP. F.HectorR. E.PanavasT.SternerD. E.QureshiN.BischoffK. M.BangS. S.MertensJ. A.JohnsonE. T.LiX.-L.JacksonJ. S.Jr.CaugheyR. J.RiedmullerS. B.BartolettS.LiuS.RichJ. O.FarrellyP. J.ButtT. R.LaBaerJ.CottaM. A.Lycotoxin-1 insecticidal peptide optimized by amino acid scanning mutagenesis and expressed as a coproduct in an ethanologenic Saccharomyces cerevisiae strain. J. Pept. Sci.2008, 14(9), 1039–1050.
16.
UetzP.GiotL.CagneyG.MansfieldT. A.JudsonR. S.KnightJ. R.LockshonD.NarayanV.SrinivasanM.PochartP.Qureshi-EmiliA.LiY.GodwinB.ConoverD.KalbfleischT.VijayadamodarG.YangM.JohnstonM.FieldsS.RothbergJ. M.A comprehensive analysis of protein-protein interactions in Saccharomyces cerevisiae. Nature2000, 403, 623–627.
17.
PhizickyE.BastiaensP. I. H.ZhuH.SnyderM.FieldsS.Protein analysis on a proteomic scale. Nature2003, 422, 208–215.
18.
QureshiN.SahaB. C.CottaM. A.Butanol production from wheat straw hydrolysate usingClostridium beijerinckii. Bioprocess Biosyst. Eng.2007, 30, 419–427.
19.
SahaB. C.CottaM. A.Ethanol production from alkaline peroxide pretreated enzymatically saccharified wheat straw. Biotechnol. Prog.2006, 22, 449–453.
20.
SahaB. C.CottaM. A.Lime pretreatment, enzymatic saccharification and fermentation of rice hulls to ethanol. Biomass Bioenergy.2008, 32, 971–977.