
Abstract
The social interaction test is based on spontaneous behaviour observed between pairs of animals, usually rodents. Commonly, the behaviour of one of the members in the dyad is related to the behaviour of its partner; therefore, making accurate predictions about behaviour is difficult, and the behaviour of the dyad cannot be controlled. In the present study, we programmed an e-puck robot with simple behavioural patterns, such as moving around a cage and following and approaching a rat. The results were analysed by comparing behaviour that was displayed by two groups of experimental rats towards different types of partners: (i) in the first group of experimental rats, they interacted with another group of rats and (ii) in the second group of experimental rats, they interacted with the e-puck robot.
Our aim was to study the behaviour of experimental rats in the social interaction test when the interaction partner is a pre-programmed robot, in order to find out whether a rat is able to display social interaction activities in that context. Those activities were evaluated by a structured scheme of possible behaviours, quantified in categories according to currently well-accepted nomenclature and definitions. In order to achieve this goal, we compared the social and non-social behaviour displayed by the experimental rat in rat-rat and rat-robot interactions (such as approaching and following behaviour). We observed predominantly non-social behaviours, such as exploring the cage, when the experimental rats confronted either another rat or the robot. The experimental rats displayed similar periods of approaching, sniffing and crawling (social behaviour), exploring, being quiet, self-grooming and evading (non-social behaviour) in encountering both the rat and the robot. However, in the presence of the robot, the experimental rats displayed long periods of time spent in following, in contrast to short periods of immobility. In the present study, we explored a behavioural repertoire that was classified into the social and the non-social, in which the robot was usually able to elicit social behaviour from the rat. The results of our experiments open possibilities for additional studies on social interaction in robot-live rat dyads (e.g., in predator-prey models).
1. Introduction
Recently, animal models for psychopathology have become an invaluable tool in the analysis of a multitude of causes, e.g., genetic, environmental or pharmacological, that can bring out symptoms homologous to those of patients with a specific disorder [1]. Furthermore, animal models are particularly helpful in situations when the impact of stress cannot be studied in humans because of ethical and other similar reasons [2]. In the field of anxiety research, animal models can be grouped into two main classes. The first involves the animal's conditioned responses to stressful and often painful events (e.g., exposure of the foot to electric shocks), and the second involves unconditioned responses, including ethologically inspired paradigms and the animal's spontaneous and natural reactions (e.g., flight, avoidance or freezing) to stress stimuli that do not explicitly involve pain or discomfort [1].
The social interaction test of unconditioned responses was the first animal test of anxiety that endeavoured to use ethologically relevant sources of anxiety, and to use a natural form of behaviour. The dependent variable is the time spent in social interaction (e.g., sniffing, following or grooming the partner) by pairs of male rats. An increase in social interaction, without a related increase in motor activity, is indicative of an anxiolytic effect, whereas a specific decrease in social interaction indicates an anxiogenic effect [3].
In our work, the social interaction test relies on the study of a rat dyad that is placed in an acrylic box in which the unconditioned responses of one of the members in the pair can be observed [4]. When a pair of rats that had not previously been in contact (and a hierarchy had not been established) are placed together, each member of the dyad exhibits both social and non-social behaviour. Behaviour that is directed towards the other member of the dyad is considered social behaviour. Self-directed and exploratory behaviours are classified as non-social behaviour. These behavioural patterns are quantifiable with regard to their duration, latency of initial occurrence, total duration (i.e., the time that elapses within periods and the summation of periods for any given behaviour during the test) and frequency during the test [2–7]. In a dyad, rats exhibit behaviour spontaneously; thus, the specific stimuli that trigger particular behavioural patterns are difficult to determine in one of the partners. A possible alternative to control for some of these stimuli is a programmable entity that repeatedly executes a particular kind of behaviour. This can be achieved using a robot [8, 9] that can be programmed to perform a particular set of actions. Consequently, at least one individual in the dyad can be controlled.
An industrial classification for robots was developed by the Japanese Industrial Robot Association, which includes autonomous robots [8, 10] and can be considered as a benchmark for research in artificial intelligence. For these robots, some specially designed software roughly emulates the behaviour observed in living organisms [11]. Mobile autonomous robots assume that actions and perceptions are structurally interrelated through the use of sensors and actuators. Robots perceive their environment using sensors. An onboard processor processes information, and the robot responds by activating its motors [12]. Robots act in structured environments by moving, avoiding obstacles and handling objects [12]. Inspired by findings from neuroscience, psychology and ethology [13], the concept of bio-inspired robotics [14–23] allowed the development of robots based on animal behaviour [16]. Behaviour-based robotics is a branch of robotics that bridges artificial intelligence, engineering and cognitive science. Its twin goals are: (1) to develop methods for controlling artificial systems, ranging from physical robots to simulated ones and other autonomous software agents and (2) to use robotics to model and understand biological systems more fully: typically, animals ranging from insects to humans [17].
For example, the e-puck robot was originally designed for micro-engineering teaching and research [24], and it can be programmed to display standardized behaviour similar to that which is commonly observed in social interaction tests in rodents (e.g., approaching and following).
In the literature, Shi et al. [25–27] implemented a pioneering approach for the development of a series of rat-like robots, called WR-3 and WR-4. These devices are able to elicit behavioural responses from rats [25–26]. In this work, the devices were useful for studying the actions of psychotropic drugs [27]. As a result, they employed a social interaction model that consisted of living animals and a robot. In their experiment, the authors included different variables in the open field test (locomotion, rearing and body grooming). Both the study of Shi et al. and the study we are presenting in this paper were based on the hypothesis that relevant responses can be produced in rats using a programmable device; however, there are some differences between the two studies with regards to methodology. Shi et al. mainly focused on the possibility of developing a model for measuring rat behaviour, while, in the present study, we are focusing on robot-rat social interaction in which a robot is programmed to exhibit only two behaviours (approaching and following) but our main focus is the behaviour of the rat in response to the robot. As a result, we based our observations on a repertoire of social and non-social behaviour in rats.
In our work, we explored the ability of an e-puck robot to elicit social and non-social behaviours in the context of the social interaction model in rats [4, 28]. We hypothesized that a complete social interaction test and its evaluation may be fully tested in a rat-robot dyad. The aims of this research have three aspects: 1) studying the behaviour of the experimental rats in the social interaction test when their partner is a robot programmed to perform the social behavioural patterns of the rat (i.e., approach and follow); 2) comparing these responses with the behaviour of experimental rats that interacted with another rat and 3) developing social-interaction studies to provide a better understanding of social behaviour in rat dyads when the behaviour of at least one of the interaction partners is controlled. In order to achieve these goals, we compared spontaneous behaviour exhibited by experimental rats that were placed in dyads with either another rat or with the e-puck robot. The behaviour of the experimental rat in the dyad was evaluated by comparing its social and non-social behaviour exhibited when confronting either another rat or the robot.
2. Material and Methods
2.1. Ethics
The animal procedures were performed in strict accordance with the National Institute of Health guidelines, the international and institutional standards for the care and use of animals in research [29] and Norma Oficial Mexicana (NOM-062-ZOO-1999) [30]. Authorization was obtained from the Biomedical Research Institute Ethical Committee of the National Autonomous University of Mexico.
2.2. E-puck robot
A small mobile robot was programmed to roughly emulate the behaviour of a rat that moves around a box while avoiding walls, and approaching and following a live rat. The e-puck robot [24] (Figure 1) was programmed using an action-selection model to control its motor plant, which is based on its sensor system that detects and responds to the movements of a living partner (i.e., a rat).

The e-puck robot [24]
2.3. E-puck control program
The e-puck robot was programmed in MPLAB [31] using the utility MPLABC30 [32], the E_puck_Lib e-puck library [33] and program code in the C programming language. The final compiled program was downloaded to the PIC micro-controller of the e-puck robot using Tiny Bootloader [34].
An action-selection mechanism was implemented to utilize the infrared sensors on the e-puck robot. Some of the calculations were used to add the internal state of behavioural patterns. In turn, the selection model sends motor commands to the robot. The behavioural patterns of the robot consisted of detecting a rat, following a rat, detecting a wall and avoiding a wall. By default, avoidance was executed, but the robot was able to follow and approach a rat when its presence was detected (Figure 2).
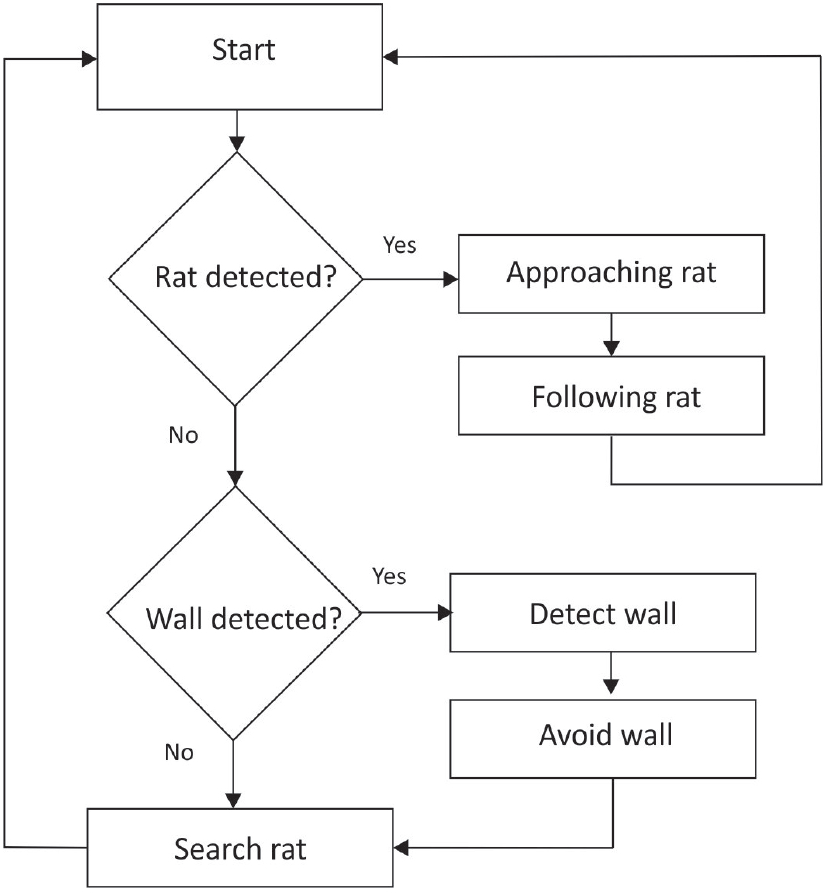
An action-selection program designed for an e-puck robot's behaviour. The robot was programmed to avoid walls in the box, with the priority being approaching and following the rat.
For avoidance behaviour, we calculated the action potential by normalizing readings from the infrared sensors 0 < iri < 4095 in values that ranged from 0 to 1 (0 < irzi < 1). If no obstacle was detected, then the value of the infrared sensor was near 1; otherwise, it was near 0.
Activation was calculated as the following:



Activation passes through a piecewise-linear function li = F(ai), which allows values in the range of −1000 ≤ l ≤ 1000. For following behaviour, the linear velocity (vl) is calculated, together with the angular velocity (va), to set motor speeds (port and starboard).


The linear-velocity function f lin(x,z) calculates speed as the scalar sum of the left infrared sensors (x) and the scalar sum of the right infrared sensors, while f ang(x,z) is used to calculate angular speed, with x for the scalar sum of the left infrared sensors and z as the scalar sum of the right infrared sensors.
2.4. Living subjects
The study included 33 adult male Wistar rats, which were three months old and weighed 300–350 g. The animals were randomly allocated to six groups of five rats, and one group of three rats. All groups were kept in plastic boxes (45 cm × 30 cm × 30 cm), with a 12 h/12 h light/dark cycle (lights on at 7:00 AM), and ad libitum access to water and food. The bedding material was changed daily, and fresh food and water were also placed in the cages every day. Rats were gently handled for 5 min daily for at least one week before the test. For the experiment, we worked with a total of 33 rats, from which we randomly chose 22 individuals that had not previously been in contact, and divided them into pairs (rat-rat dyads). Next, the remaining 11 rats were paired with the robot (rat-robot dyads). Circular marks in different colours on the rats' tails identified individuals belonging to the two different groups.
2.5. Experimental groups of rat-rat pairs
The social interaction test was conducted using the following dyads: (i) experimental rats (n = 11) randomly paired with other rats (n = 11), when the rats had not previously been in contact with each other and (ii) a different group of experimental rats (n = 11) that were paired with the robot. It is important to note that all rats were used only once in the test.
2.6. Social interaction test
In our test, we used an acrylic box (44 cm length × 33 cm width × 20 cm height) lit by low light. To encourage habituation to the cage, rats were separately placed in similarly sized cages for 5 min before being introduced in pairs to the test cage. Each test lasted 10 min and was videotaped for later analysis. After each experimental session, the test cage was carefully cleaned with a cleaning solution (0.5% ammonia, 15% ethanol, 10% Extran, 5% isopropyl alcohol, 19% Pinol and 50.5% water). To allow the scent to disperse, the boxes were left to dry for 5 min before starting a new test.
2.7. Data analysis
The data analysis focused on the behaviour of the experimental rats. Behaviour was divided into categories according to the nomenclature and definitions used by Brain et al. [28] (Table 1). We used two independent observers who were unaware of the correspondence between the coloured marks on the rats' tails or the set-up of the experimental groups. Only data with > 95% agreement among observers were included in the statistical analysis. Tailor-made software allowed us to measure the latency, frequency and duration of each recorded behaviour.
Behavioral categories included in the study (adapted from Brain et al. [28])

The collection and analysis of the data focused only on experimental rat groups when they confronted a different partner (another rat or the robot). The latency (initial occurrence), frequency (number of events during the session) and time (in periods that accumulated over the course of a 10-min session) for each behaviour were evaluated using two-way analysis of variance (ANOVA; SigmaStat 3.5).
For the analysis, one factor was formed with the two different dyads (rat-rat dyad and rat-robot dyad) and was named factor A. A second factor analysed differences in behaviour and was named factor B (approaching, sniffing, following, crawling, exploring, being quiet, immobility, self-grooming and evading). The analysis determined the level of significance between factor A and factor B. When the data failed to follow a normal distribution and equal variance, they were transformed by ranks. Values of p ≤ 0.05 were initially considered to be statistically significant. When comparisons within groups were made (nine behavioural patterns), Bonferroni adjustment indicated a valid p < 0.005. For comparisons of behaviour between groups, Bonferroni adjustment indicated p < 0.025, and the Holm-Sidak post hoc level of significance was fixed at p < 0.01. The results are expressed as mean ± standard error.
3. Results
3.1. Qualitative aspects
Strong similarities were seen in qualitative behavioural patterns in the experimental rats faced with the robot compared with those of experimental rats that were paired with other rats (Figures 3, 4). Two behavioural patterns were excluded from the data analysis. First, push-past was always observed in the presence of another rat but never occurred when the experimental rats confronted the robot. Second, grooming was rarely observed in the presence of either another rat (18%) or the robot (18%). No aggressive behavioural patterns were detected towards either another rat or the robot (e.g., attacking, biting or adopting an upright offensive posture) [28]. Grooming and push-past behaviour were discarded from the data analysis. Both of these behavioural patterns require close proximity to a partner, and push-past behaviour occurs when two rats move in opposite directions. As reported in the methods section, the robot was programmed to follow the rat; as a result of this we observed a low probability in the occurrence of this behaviour.
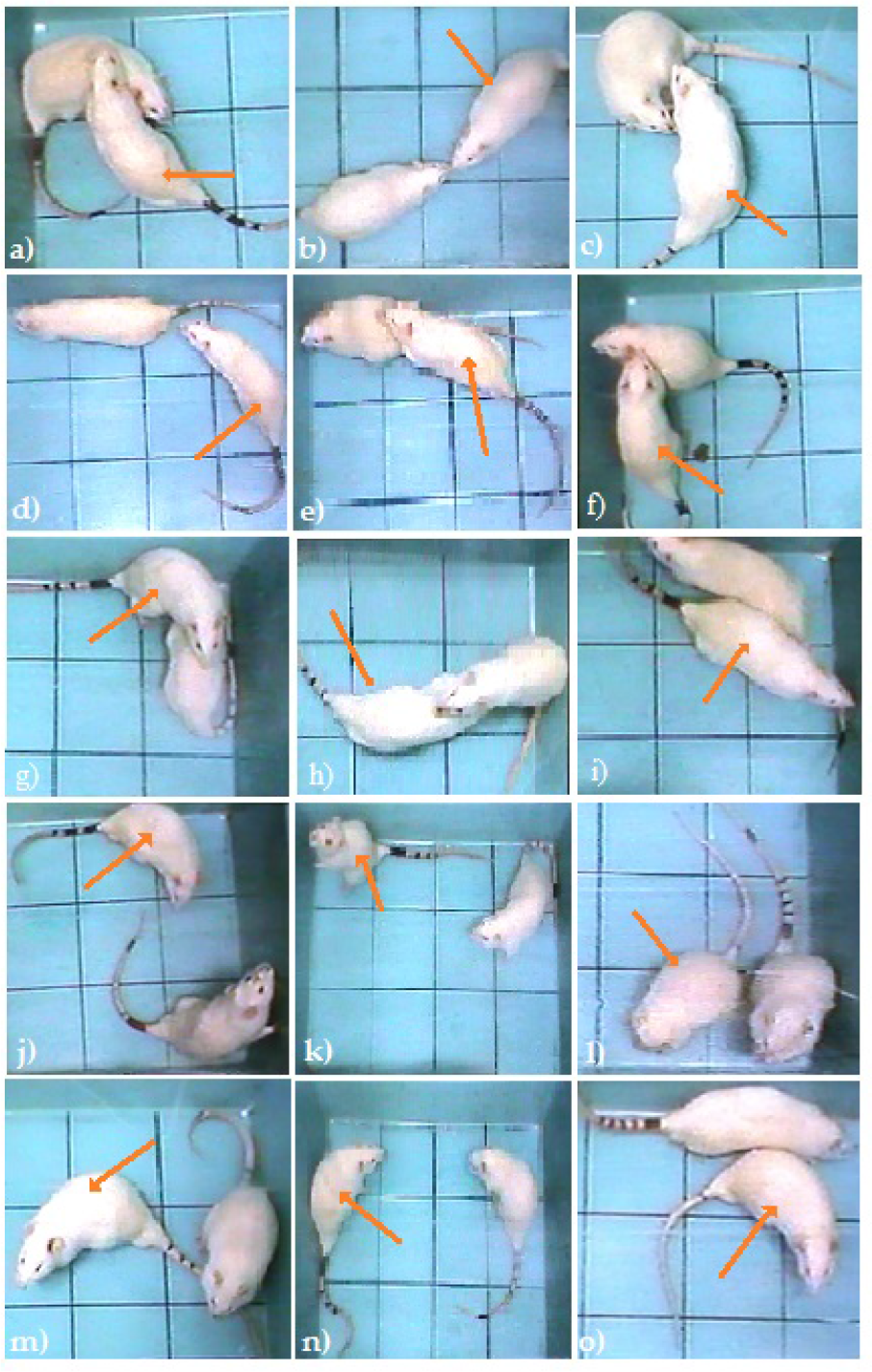
Rat-rat dyads. Behavioural categories included approaching (a), sniffing (b, c), following (d, e), grooming (f), crawling (g, h), push-past (i), exploring (j, k), self-grooming (l), immobility (m), being quiet (n) and evading (o).
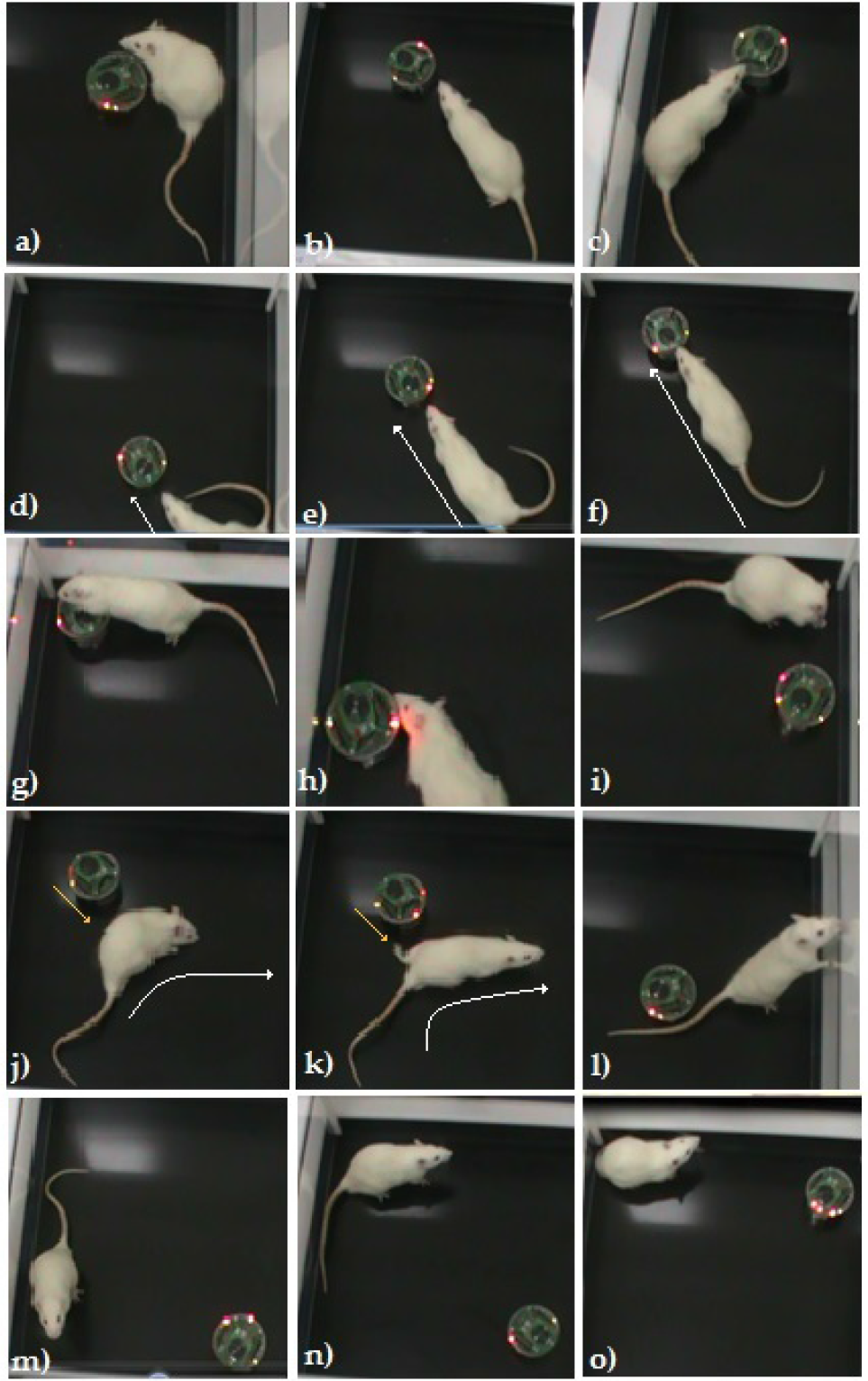
Rat-robot dyads. Behaviour displayed by the experimental rats included approaching (a, b), sniffing (c), following (d–f), crawling (g), grooming (h), self-grooming (i), evading (j, k), exploring (l, m), immobility (n) and being quiet (o). These behavioural patterns were similar to those observed in the rat-rat dyad, except push-past, which was not observed in the presence of the robot.
3.2. Quantitative aspects
3.2.1. Latency
The two-way ANOVA by ranks test did not detect differences in factor A (F1, 180 = 7.022, p = 0.009). The analysis of the latency of behavioural patterns in the dyad, regardless of the partner (either rat or robot), indicated significant differences (F8, 180 = 35.519, p < 0.001) for the initial occurrence of behaviour. The general profile, independent of the partner type, indicated that the first observed behaviour was exploring, which had the shortest latency period. The Holm-Sidak post hoc test (p < 0.01) allowed us to form two groups of different behavioural patterns based on latency. One group was named the short-latency group (< 30 s) and consisted of exploring, approaching, sniffing and evading. The second group was named the long-latency group (> 30 s) and consisted of following, crawling, being quiet, self-grooming and immobility (Table 2).
Behavioral categories according to latency
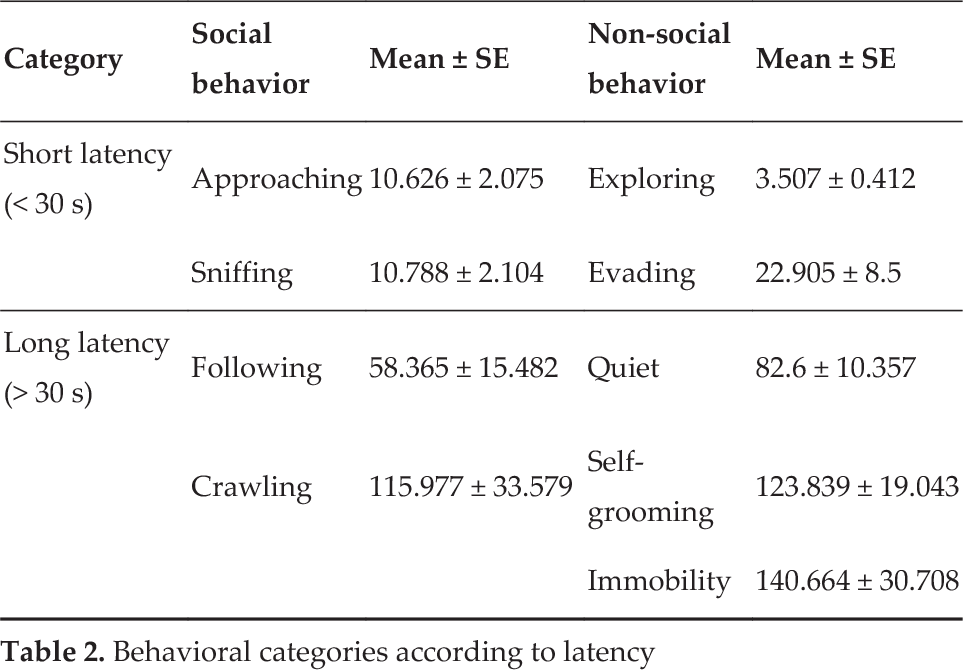
The interaction between factors did not reach the level of significance (F8, 180 = 0.821, p = 0.585).
3.2.2. Frequency
The analysis of the frequency of behaviour was statistically significant (F8, 180 = 146.233, p < 0.001). The post hoc analysis allowed the formation of a general profile. Less frequently observed behaviour included crawling and following. In contrast, behaviour with a high frequency included sniffing, approaching, exploring, being quiet, self-grooming, evading and immobility (Table 3).
General profile of behavioral categories according to frequency

The interaction between factors reached the criterion of significance (F8, 180 = 5.699, p < 0.001). The particular profiles for the experimental rats that confronted another rat and the ones that confronted the robot were similar to the general profile. However, some differences between the subjects were detected (Figure 5). The frequencies of sniffing and approaching were significantly higher (p < 0.01), whereas immobility (p < 0.01) was observed less often in the experimental rats that confronted the robot when compared with those that confronted another rat.

Frequency of the rats' behaviours in the presence of the robot when compared with the rat-rat dyad. The experimental rats frequently exhibited two social behaviours (sniffing and approaching; left) and, less frequently, one non-social behaviour (immobility; right; *p < 0.01; Holm-Sidak post hoc test).
3.2.3. Cumulated time
The differences in the cumulated time of behaviour, regardless of the nature of the partner, reached statistical significance (F8, 180 = 135.354, p < 0.001). Similar to the latency results, the Holm-Sidak post hoc analysis (p < 0.01) allowed the integration of a general profile (Table 4). The expression of behaviour was classified into three different intervals of time depending on the mean time of the behaviour engaged in during the 10 min test. The first interval was named short (< 10 s), in which the experimental rats displayed following and crawling behaviour. The second interval was named moderate (10–60 s), in which the rats displayed approaching, evading, immobility and self-grooming. The third interval was named long (> 60 s), in which the rats displayed sniffing, being quiet and exploring.
Behavioral category according to the test time
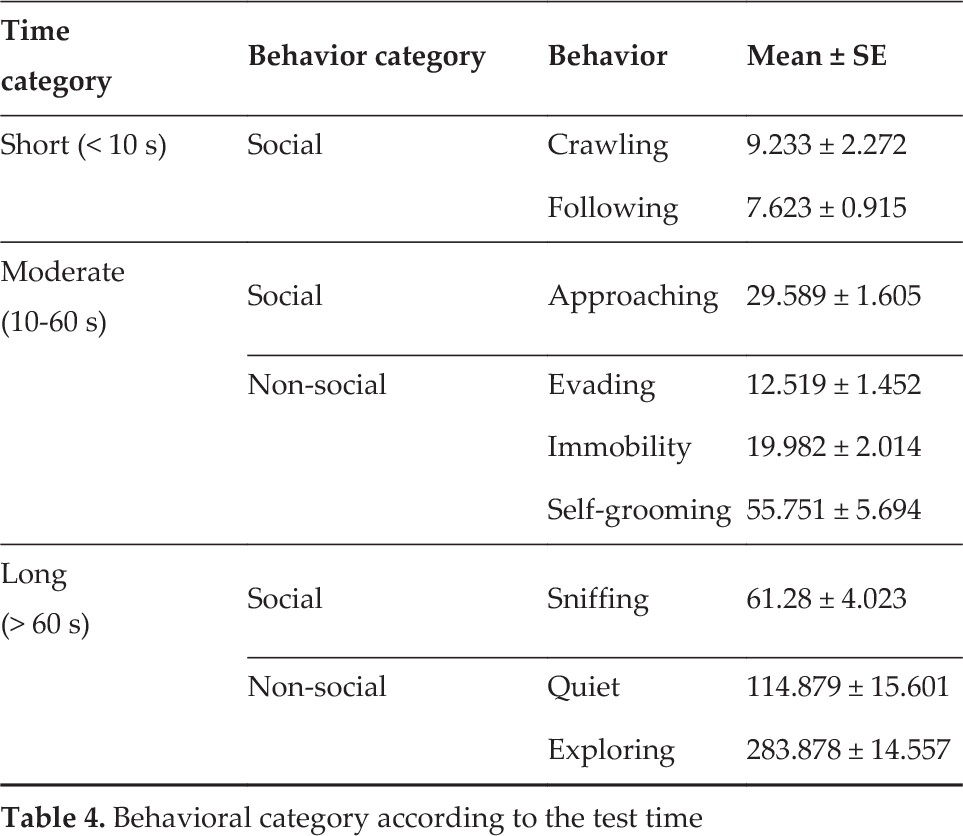
The interaction between factors reached statistical significance (F8, 180 = 4.269, p < 0.001). Behavioural categorization with regard to duration resulted in a general profile that was similar to the one described above for both the rat-robot and rat-rat dyads. Comparisons between subjects revealed a statistically significant difference (p < 0.01, Holm-Sidak). Thus, for the rat-robot pairs, we observed shorter intervals of immobility and longer intervals of following in contrast to rat-rat pairs (Figure 6).

Cumulated time. For the rat-rat and rat-robot dyads, the experimental rats spent a similar time engaged in social (left) and non-social (right) behaviours. However, in the presence of the robot, the experimental rats displayed longer following and shorter immobility times compared with experimental rats that confronted other rats (*p < 0.01) (Holm-Sidak).
3.3. Social vs. non-social behaviours
The two-way ANOVA by ranks test did not indicate differences in latency (F1, 194 = 2.872, p = 0.092), frequency (F1, 194 = 0.336, p = 0.563) and cumulative time (F1, 194 = 0.016, p = 0.899) for both the rat and robot groups. Comparisons of social vs. non-social behaviours did not indicate differences in latency (F1, 194 = 5.589, p = 0.019) and frequency (F1, 194 = 0.570, p = 0.451), but a significant difference in time was observed (F1, 194 = 31.135, p < 0.001). During the 10-min test, the most noticeable behaviour exhibited was non-social with regard to the mean duration (non-social: 97.4 ± 10.52 s; social: 26.9 ± 2.62 s) and cumulative time (non-social: 487.0 ± 52.59 s; social: 107.7 ± 10.48 s).
The interaction between factors did not reach statistical significance for latency (F1, 194 = 0.033, p = 0.856), frequency (F1, 194 = 2.725, p = 0.100) or time (F1, 194 = 0.655, p = 0.419).
4. Discussion
In the social interaction test, the dependent variable is the time spent in social interaction by pairs of male rats. The behaviour of one rat influences that of the other; therefore, making accurate predictions about behaviour is difficult, and controlling behaviour in the dyad is impossible. In the present study, we programmed a robot to produce constant stimuli directed towards the rat for the social interaction test. Therefore, we decided to develop a general behaviour for the e-puck robot, roughly emulating the social behaviour of a rat, e.g., approaching and following behaviour. As a consequence, the e-puck robot acts as a form of controlled stimulus. Our results indicated that the experimental rats generally displayed similar behaviour towards both the rat and the robot, indicating that rat-robot dyads can be used in the social interaction test.
The study of animal behaviour can be achieved by replacing a real animal with a robot [17], relying on an approach based on neuroscience [16, 21–23]. In the present study, a robot was programmed with behavioural patterns that mimicked a living subject, such as following and approaching. Behaviour that had the shortest latency, longest duration and highest frequency, respectively, were exploring (non-social behaviour), approaching and sniffing (social behaviour). Behaviour was exhibited regardless of the nature of the other member in the dyad (rat or robot). In the present study, non-social behaviour exploration was commonly observed as a primary display, which is behaviour commonly observed in most animals and is profoundly rooted in the behavioural repertoire of the rat [28, 35]. Additionally, the robot was able to elicit social behaviour from the experimental rats. Following behaviour had the longest duration, but approaching and sniffing behaviours occurred with a high frequency as well.
In other studies of social interaction between rats and robots [25–27], the avoidance of a moving robot was interpreted as “enemy avoidance,” which ultimately depends on functional connections in the hippocampus [36] and is gender dependent [37]. We did not specifically measure avoidance, but evasion presents behavioural similarities. Differences in evasion were not found in experimental rats when they confronted either the rat or the robot. Obtained results appear to indicate that “enemy avoidance” did not occur. The high levels of social interaction that were directed towards the robot indicate that the experimental rats were not in an “anxious” state, which would be reflected by a reduction of social interaction [38, 39].
In fact, when the dyad was formed by pairing an experimental rat and the robot, social behaviours were frequently observed in the form of approaching, sniffing and following. These social behaviours can be interpreted as exploratory activities directed towards the robot. Therefore, the robot appeared to elicit curiosity from the rat. In contrast, the dyad that exhibited non-social behaviour was characterized by short periods of resting (immobility), given that the robot was programmed to follow the rat continuously.
In summary, the experimental rats indeed confronted the robot, which may open possibilities for further studies. For example, some behavioural profiles in predator-prey interactions have been identified [35, 40 and 41]. Some scents that are produced by natural predators [42, 43] act as alarm pheromones [44] that modify behaviour in receptor animals. A future experiment may consist of adding a sensorial stimulus to the robot that resembles at least one of the main odour features of a natural predator.
Among mammals, different types of communication have been identified, one of which are olfactory signals, which represent a rich source of information about the environment [45]. In primates, visual cues constitute the main source of information concerning the environment [46]. On the other hand, in rodents, the main sensorial cues are olfactory ones [47], including chemosignals [48] and pheromones [49, 50]. Additionally, rats also use their visual system to sense information in the environment. The rat visual system consists of neural circuits that are related to emotional memory (e.g., the amygdala and hippocampus [51]), and relay sensory information to the cingulate and frontal cortical areas [52].
In the present study, when the rat was paired with the robot, the robot should have been perceived as a moving object by the rat's visual system. The experimental rats behaved similarly when placed with either another intact rat or the robot. Thus, it is safe to assume that rats perceived the robot as a non-threatening entity.
Shi et al. [25–27] also utilized a social interaction model that consisted of living animals and a robot. The present authors part from the hypothesis of that contribution, that a programmable device is able to produce relevant responses in rats. We share the same hypothesis, although the main contribution of our work is that we based our observations on a repertoire of social and non-social behaviour in rats. As a result, the robot elicited social behaviour from rats, opening the possibility for further studies of social interaction in robot-rat dyads.
5. Conclusion
A robot action-selection model was used in the social interaction test with experimental intact rats. This set-up promoted patterns of social behaviour in rodents, hence, facilitating behavioural control of at least one interaction partner (i.e., the robot). Therefore, our work provides a general framework for studying different behavioural patterns in rats triggered by robot behaviour.
Footnotes
The authors declare that there are no conflicts of interest.
7. Acknowledgements
The authors are grateful to Michael Arends for revising and editing the manuscript. This work was supported by Universidad Nacional Autónoma de México (UNAM: DGAPA-PAPIIT IN211111-3), CONACyT-MEXICO (Grant SEP no. 0100895) and Sistema Nacional de Investigadores (Exp. 754 [CMC], 32755 [AGGG], 30026 [FMMG]). R. del Angel-Ortiz received fellowships from CONACyT (Reg. 377475).