
Abstract
Objective
In this study, we aimed to investigate the expression and function of interleukin-4 receptor (IL-4R) in hepatocellular carcinoma (HCC).
Methods
We collected 40 pairs of human HCC and adjacent normal tissue specimens and examined the expression levels of IL-4R. After IL-4R knockdown in HCC cell lines, cell proliferation and invasion ability were examined. Cell cycle and apoptosis were analyzed by flow cytometry. The activity of multiple signaling pathways was examined by Western blot.
Results
IL-4R was overexpressed in HCC tumors compared with adjacent normal control tissues and was associated with tumor differentiation status. IL-4R knockdown resulted in enhanced apoptosis, impaired proliferation and reduced invasion of HCC cells. Furthermore, IL-4R knockdown abolished IL-4-induced activation of the Janus Kinase 1 (JAK1)/signal transducer and activator of transcription 6 (STAT6) and JUN N-terminal kinase (JNK)/extracellular signal-regulated kinase 1/2 (ERK1/2) signaling pathways.
Conclusions
IL-4R plays an important role in regulating HCC cell survival and metastasis, and regulates the activity of the JAK1/STAT6 and JNK/ERK1/2 signaling pathways. We therefore suggest that IL-4/IL-4R may be a new therapeutic target for HCC.
Introduction
Liver cancer is the second most common cause of death from cancer worldwide (1). Hepatocellular carcinoma (HCC) is a major form of liver cancer. Despite recent advances in interventional surgery, radiofrequency ablation and chemotherapy, advanced-stage and metastatic HCC remains challenging to treat. Accordingly, understanding the molecular mechanisms underlying HCC tumorigenesis and finding novel therapies for HCC is urgently needed.
Interleukin-4 (IL-4) is a multifunctional pleiotropic cytokine secreted by activated T helper 2 (Th2) cells. The interaction between IL-4 and IL-4 receptor (IL-4R) plays an important role in regulating immune responses, including Th2 cell differentiation (2, 3), parasite clearance (4), mucus secretion and eosinophil granulocyte activation (5). In addition to its immune functions, recent studies suggest that IL-4R is frequently overexpressed in different cancer types including breast cancer, colon cancer and malignant pleural mesothelioma (6-10), and might directly regulate multiple signaling cascades implicated in tumorigenesis. For example, hyperactivation of IL-4/IL-4R signaling might induce a prometastatic phenotype in breast cancer via regulating tumor cell migration, proliferation and survival (11). It has also been shown that IL-4/IL-4R might contribute to resistance to chemotherapeutic drugs in Hodgkin/Reed-Sternberg lymphoma (12). Moreover, IL-4/IL-4R might enhance chemoresistance in epithelial tumors through inhibition of apoptosis (13). Notably, it has been shown that IL-4/IL-4R protects HCC cells from apoptosis induced by transforming growth factor-beta (TGF-β) (14).
We previously showed that a natural product, pigenin, could inhibit the growth of HCC in vivo and in vitro through targeting IL-4R and inducing apoptosis (15). Recently, we performed immunohistochemistry analysis of HCC tumor samples and found that IL-4R expression is strongly elevated in tumor tissues compared to adjacent normal tissues. These data suggest a putative oncogenic role of IL-4/IL-4R in HCC. To further investigate the biological function of IL-4/IL-4R in HCC, we knocked down IL-4R in HCC cells and found that IL-4R knockdown resulted in enhanced apoptosis, impaired proliferation and reduced invasion of HCC cells. The mechanisms might involve inhibition of the JAK1/STAT6 and JNK/ERK1/2 signaling pathways. We thus suggest that IL-4R is a key regulator of HCC cell survival and metastasis and may be a new therapeutic target for HCC.
Materials and methods
Immunohistochemistry
We collected 40 paraffin samples of human HCC tissues and corresponding adjacent nontumor tissues at the Second Affiliated Hospital of Nanchang University between September 2012 and April 2013. The study was approved by the Second Affiliated Hospital Review Board and signed informed consent was obtained from each study participant. Each sample was cut into 6 slices and 3 pieces were randomly selected. The sections were washed with distilled water, placed in antigen retrieval buffer (pH 6.0) and antigen-restored in a microwave oven. After cooling and washing with PBS, sections were incubated in 3% hydrogen peroxide solution for 20 minutes in the dark to block endogenous peroxidase and then blocked by goat serum at room temperature for 15 minutes. Next, primary antibody diluted with 5% BSA (IL-4R, 1:100) was added and incubated overnight at 4°C. The next day, sections were washed with PBS and incubated for 50 minutes at room temperature with secondary antibody (1:100). The sections were washed again, and color was developed by incubating the sections with freshly prepared DAB solution followed by reaction termination with distilled water. Lastly, samples were counterstained with Harris hematoxylin, differentiated by 1% hydrochloric acid alcohol, turned back to blue with ammonia, dehydrated to transparency, and sealed with neutral gum.
Cell lines and culture
Human HCC cell lines (HepG2, HepG2.2.15, Hep3B and Huh7) and the normal liver cell line THLE-2 were purchased from the Cell Bank of the Chinese Academy of Sciences. Cells were cultured in Dulbecco's modified Eagle's medium (DMEM; Solarbio) and supplemented with 10% fetal bovine serum (FBS) (ExCell Biological Products). G-418 sulfate (380 μg/mL; Solarbio) was added to DMEM with 10% FBS to maintain HepG2.2.15 cells. All cells were incubated at 37°C with 5% CO2.
RNA interference
Small interfering RNAs (siRNAs) targeting IL-4R were designed and synthesized by GenePharma. Nontargeting siRNA was used as negative control. The siRNA sequences were as follows: IL-4R siRNA, sense 5′-CAGUGCGGAUAACUAUACATT-3′, antisense 5′-UGUAUAGUUAUCCGCACUGTT-3′; control siRNA, sense 5′-UUCUCCGAACGUGUCACGUTT-3′, antisense 5′-ACGUGACACGUUCGGAGAATT-3′. Individual siRNAs (80 nM) were transfected into cells with Lipofectamine® 2000 Transfection Reagent (Invitrogen) according to the manufacturer's instructions. After incubation for 48 and 72 hours, transfected cells were collected for analysis of mRNA and protein expression.
Real-time PCR analysis
Total RNA was extracted using Trizol (TransGen Biotech) and cDNA synthesis was performed using the PrimeScript RT reagent Kit (Takara). Furthermore, qPCR was performed using the SYBR Premix Ex Taq™ II (Takara) on an Applied Biosystems 7900 Real-Time PCR System. The IL-4R primer sequences were as follows: sense 5′-CGTGGTCAGTGCGGATAACTA-3′, anti-sense 5′-TGGTGTGAACTGTCAGGTTTC-3′. The primer sequences for amplification of GAPDH were: sense 5′-GTTGGAGGTCGGAGTCAACGGA-3′, anti-sense 5′-GAGGGATCTCGCTCCTGGAGGA-3′. All PCR amplifications were performed with an initial denaturation at 95°C for 30 seconds, followed by 40 cycles of 95°C for 5 seconds and 60°C for 30 seconds.
Western blot analysis
The whole cell lysates were prepared using lysis buffer (50 mmol/L Tris-HCI [pH 8.0], 150 mmol/L NaCI, 0.5% NP-40, 5 mmol/L EDTA [pH 8.0], and 0.1% SDS) containing aprotinin (2 mg/L), the phosphatase inhibitor leupeptin (5 mg/L), and phenylmethylsulfonyl fluoride (PMSF; 1 mmol/L; Solarbio). Protein concentrations were monitored using the BCA protein assay (Thermo Fisher Scientific). Equal amounts of proteins were loaded and separated by 10% SDS-polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes. The membranes were blocked with 5% fat-free milk at least 2 hours and incubated with primary antibodies at 4°C overnight, followed by incubation with secondary antibodies for 1 hour. Densitometric analysis was performed with the ImageJ software. The primary antibodies used in this study were mouse monoclonal anti-β-actin (Proteintech, 1:10,000), mouse monoclonal anti-GAPDH (Proteintech, 1:10,000), mouse monoclonal anti-tubulin (Cell Signaling Technology, 1:1,000), rabbit polyclonal anti-IL-4R (Abcam, 1:1,000), rabbit polyclonal anti-MMP-2, rabbit polyclonal anti-MMP-9 (Cell Signaling Technology, 1:1,000), rabbit polyclonal anti-JAK1 (Cell Signaling Technology, 1:1,000), rabbit polyclonal anti-phospho-JAK1 (Cell Signaling Technology, 1:500), rabbit polyclonal anti-STAT6 (Cell Signaling Technology, 1:1,000), rabbit polyclonal anti-phospho-STAT6 (Cell Signaling Technology, 1:1,000), rabbit polyclonal anti-ERK1/2 (Cell Signaling Technology, 1:500), rabbit polyclonal anti-phospho-ERK1/2 (Cell Signaling Technology, 1:500), rabbit polyclonal anti-p38 (Cell Signaling Technology, 1:1,000), rabbit polyclonal anti-phospho-p38 (Cell Signaling Technology, 1:1,000), rabbit monoclonal anti-JNK (Cell Signaling Technology, 1:1,000), rabbit monoclonal anti-phospho-JNK (Abcam, 1:1,000), rabbit monoclonal anti-Akt (Abcam, 1:1,000), rabbit monoclonal anti-phospho-Akt (Abcam, 1:500), rabbit monoclonal anti-mTOR (Abcam, 1:500), rabbit monoclonal anti-phospho-mTOR (Abcam, 1:10,000), ProteinFind goat anti-mouse IgG (H + L) and ProteinFind goat anti-rabbit IgG (H + L) (HRP Conjugate, TransGen Biotech, 1:10,000).
Migration and invasion assay
Cells were resuspended in serum-free DMEM supplemented with BSA and seeded in chambers with or without Matrigel coating (Corning). The lower compartment of the chamber contained 20% FBS as a chemoattractant because the HCC cell doubling time is about 24 hours. In order to rule out the influence of cell proliferation on the invasion and migration experiments, after incubation for 24 hours, cells on the membrane were scrubbed, washed with PBS, fixed in paraformaldehyde for 30 minutes and stained with 0.1% crystal violet for 20 minutes. The cells that migrated and invaded through the membrane were visualized with an inverted microscope (Olympus).
Cell proliferation assay
Cell proliferation assay was performed using the Cell Counting Kit (Sigma) according to the manufacturer's protocol. Briefly, HCC cells were seeded in the 96-well plates at a density of 3,000 cells per well and incubated for 0, 24, 48 and 72 hours. Then the cells were incubated with 10 μL CCK-8 solution for 2 hours. The OD values were measured at a wavelength of 450 nm using a microplate reader (Thermo Fisher).
Flow cytometry analysis
Flow cytometry assay was performed using the Annexin V-PE/7-AAD Apoptosis Detection Kit I (BD Biosciences) according to the manufacturer's protocol. Briefly, 1 × 10 6 cells per well were collected and incubated with Annexin V/FITC and propidium iodide (PI) in the dark at room temperature for 15 minutes. Stained cells were examined with a FACScan flow cytometer (BD Biosciences), which was also used for cell cycle analysis. Before the test, cells were collected and fixed in 70% cold ethanol overnight at −20°C, followed by incubation with DNA staining solution and PI solution for 30 minutes in the dark at room temperature (Cell Cycle Analysis Kit, MultiSciences).
Results
Human HCC tissues express higher levels of IL-4R
To investigate the expression levels of IL-4R in human HCC, we collected 40 pairs of human HCC cancers and adjacent normal tissues. The clinical characteristics of the patients are described in Supplementary Table S1 - Clinical information of HCC patients (available online at www.biological-markers.com). Interestingly, immunohistochemistry revealed that IL-4R expression was higher in tumor tissues than adjacent normal tissues (Fig. 1A). Moreover, high expression of IL-4R was significantly associated with poor differentiation of HCC (Fig. 1A, 1B). This was independent of patient age, serum alpha-fetoprotein and serum hepatitis B surface antigen (HBsAg) (Fig. 1B), supporting a putative role in HCC tumorigenesis.
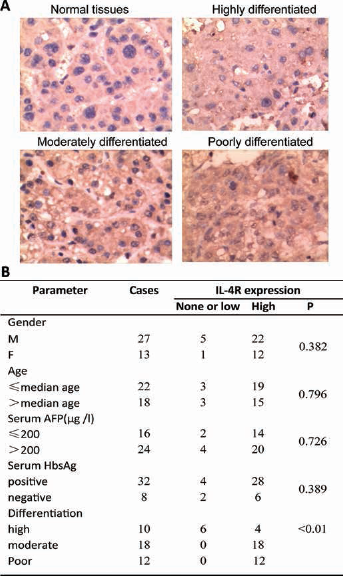
IL-4R expression in HCC tumors. (
IL-4R expression in human HCC cell lines
We then examined the expression of IL-4R in several HCC cell lines (HepG2, HepG2.2.15, Hep3B, Huh7) and a normal liver epithelial cell line (THLE-2) using Western blot. As shown in Figure 2A, protein expression of IL-4R was strongly upregulated in HepG2, Hep3B and HepG2.2.15 cells compared with Huh7 and THLE-2 cells. This was consistent with the real-time PCR results of IL-4R mRNA expression in these cells, which suggested that the expression of IL-4R was highest in HepG2.2.15 cells (Fig. 2B). We thus used HepG2.2.15 cells in the follow-up study to investigate the function of high IL-4R expression in HCC cells.
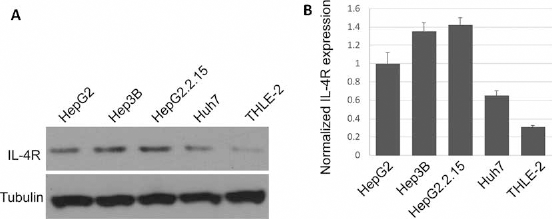
IL-4R expression in HCC cell lines. (
IL-4R depletion inhibits proliferation and induces apoptosis
To investigate the functions of IL-4R in HCC tumorigenesis, we used siRNA to knock down IL-4R expression in HepG2.2.15 cells. As shown in Figure 3A, significant downregulation of IL-4R was observed in cells transfected with IL-4R siRNA compared with cells transfected with control siRNA. Interestingly, we observed that downregulation of IL-4R by siRNA significantly reduced the proliferation rate of HCC cells compared with the no-siRNA group (mock) and control siRNA transfection group (Fig. 3B). Moreover, using flow cytometry, we observed that IL-4R knockdown significantly induced apoptosis in these cells compared with mock or control transfection (Fig. 3C and 3D). In contrast, knockdown of IL-4R had no obvious effect on the cell cycle of HCC cells. These results suggested that IL-4R might play an important role in facilitating proliferation and inhibiting apoptosis in HCC cells.
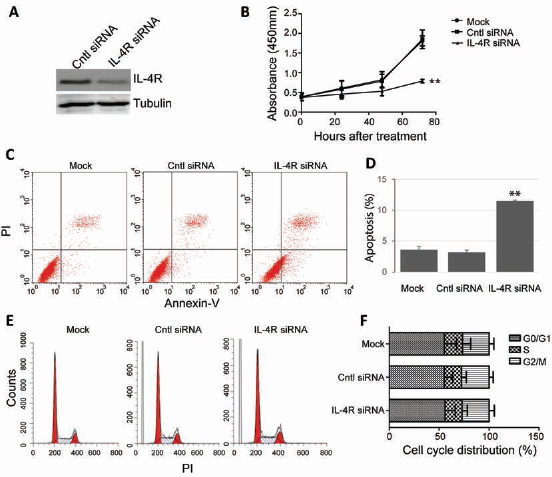
IL-4R knockdown blocks proliferation and induces apoptosis. (
IL-4R knockdown inhibits HCC cell invasion
It is well established that matrix metalloproteinases (MMPs) play an important role in cell migration/invasion and are hallmarks of tumor metastasis. Previous studies suggested a critical role of IL-4 in modulating MMP expression and tumor cell invasion. Following treatment of IL-4, the expression of MMP2 but not MMP9 was markedly elevated in HepG2.2.15 cells (Fig. 4A), suggesting that MMP2, but not MMP9, is regulated by transcription factors activated by IL-4 signaling. Strikingly, IL-4-induced MMP2 upregulation was abolished in IL-4R knockdown cells, while no effect was observed for MMP9 (Fig. 4B and 4C). We then used Transwell assays to investigate the role of IL-4/IL-4R in regulating the invasion of HCC cells. First we tested the migration of HCC cells. Significantly, IL-4R knockdown resulted in reduced cell invasion with respect to controls (Fig. 4D and 4E). These results suggested that downregulation of IL-4R can inhibit invasion of HCC cells.
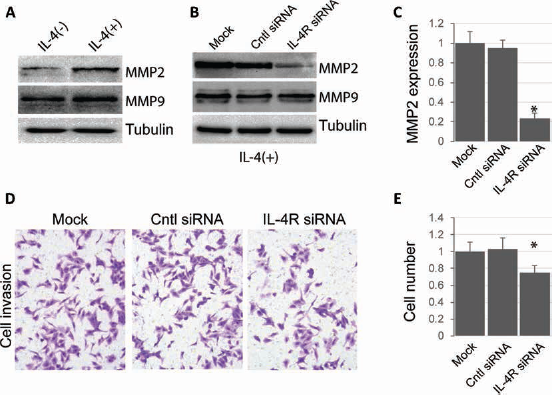
IL-4R knockdown blocks IL-4-induced cell invasion. (
IL-4/IL-4R regulates multiple signaling pathways
To investigate the mechanisms underlying the regulation of HCC cell survival and invasion by IL-4/IL-4R, we used Western blot to examine the activity of several signaling pathways in response to IL-4 treatment and IL-4R knockdown. First we tested the activity of the JAK1/STAT6 signaling pathway, which is known to be regulated by IL-4/IL-4R and plays an important role in HCC migration. Upon IL-4 treatment, the phosphorylation levels of JAK1 and STAT6 (p-JAK1 and p-STAT6, respectively) were significantly enhanced in HCC cells, while the protein levels remained largely unaffected (Fig. 5A). In addition, we also observed activation of the JNK/ERK1/2 signaling pathway by IL-4 treatment, as evidenced by enhanced levels of p-JNK and p-ERK1/2 but not total proteins (Fig. 5A). Interestingly, IL-4-induced activation of the JAK1/STAT6 signaling pathway and JNK/ERK1/2 signaling pathway was largely abolished by IL-4R knockdown, suggesting an important role of IL-4/IL-4R in the transduction of these signaling cascades. The implications of these signaling pathways in IL-4R-positive HCC require further investigation; nevertheless, our results support an important role of IL-4/IL-4R in regulating multiple oncogenic signaling pathways in HCC cells.
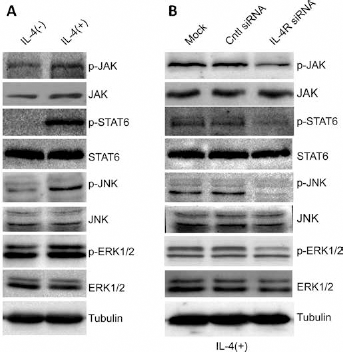
IL-4R regulates the activity of multiple signaling pathways. (
Discussion
In this study we reported that the expression levels of IL-4R in HCC tumor tissues were significantly higher than in adjacent normal tissues. In HCC cells, IL-4R knockdown resulted in cell death, proliferation inhibition and reduced invasion. This is consistent with the oncogenic roles of IL-4/IL-4R in promoting tumor proliferation and survival in various cancer types (16-19). To investigate the underlying mechanisms, we examined several key signaling pathways involved in tumor proliferation and survival, and found that IL-4R knockdown mainly blocked activation of the JAK1/STAT6 and JNK/ERK1/2 signaling pathways in HCC. These results suggest a role of IL-4/IL-4R in promoting HCC tumorigenesis.
Activation of JAK1/STAT6 by IL-4/IL-4R has been reported in multiple cell types. When IL-4 binds IL-4R at the cell surface, several Janus tyrosine kinases associated with the receptor, including JAK1 and JAK3, are activated (20). This is followed by recruitment and tyrosine-phosphorylation of transcription factor STAT6. Once phosphorylated, STAT6 is translocated to the nucleus to promote the transcription of several regulators involved in prometastatic processes including migration and invasion, enhanced proliferation and resistance to apoptosis in epithelial cancer cells (20). Abnormal activation of the JAK/STAT pathway has been implicated in the development of HCC (21). Consistent with this notion, pharmacological inhibition of JAK1 and/or JAK2 has recently emerged as an efficient approach to attenuate the oncogenic effects of IL-4/IL-4R signaling (22, 23).
The JNKs and ERKs are master protein kinases that regulate various cellular processes, such as proliferation, differentiation and apoptosis. Abnormal activation of the JNK/ERK1/2 signaling pathway has been implicated in survival and proliferation of multiple cancers (24). After activation, these protein kinases are translocated to the nucleus, where they can phosphorylate and regulate the activities of various transcription factors, leading to the systematic remodeling in gene expression essential for tumor survival. In animal models, inhibition of JNK/ERK1/2 results in major defects in liver regeneration following damage (25). Furthermore, depletion of JNK in vivo could protect the animal against carcinogen-induced HCC formation (26). Consistent with our results, previous studies showed that IL-4-induced expression of suppressor of cytokine signaling-3 (SOCS3) in B cells was dependent on activation of the JNK/ERK1/2 pathway (27).
Taken together, our results show that IL-4/IL-4R signaling plays a critical role in promoting HCC cell survival and migration. This will contribute to a better understanding of HCC's molecular etiology and establish the inhibition of IL-4R as a promising therapeutic approach for HCC in clinical practice.
Footnotes
Financial support: This work was supported by the National Natural Science Foundation of China (81560397).
Conflict of interest: The authors have no conflict of interest related to this article.