
Abstract
Since the discovery of microRNAs (miRNAs) in 1993, their role in controlling a wide variety of complex and seminal cellular functions through control of gene expression continues to be elucidated. Studies of the past decade have shown that miRNAs are able to activate or suppress target genes that are key players in the molecular pathways found to be deregulated in myelodysplastic syndromes (MDS) and acute myeloid leukemia (AML). The ability of miRNAs to act as tumor suppressor genes has been demonstrated in a number of studies in both human samples and cell lines as well as in murine models of AML and MDS. The focus of this review will be to examine the complex interaction of specific miRNAs with genes that have been implicated in MDS/AML and which may eventually become therapeutically relevant.
Introduction
In the last 30 years, the understanding of molecular mechanisms of neoplasia and malignancy has increased exponentially, with mutations in oncogenes and tumor suppressors as some of the earliest culprits described. In 1986, Friend et al used restriction fragment length polymorphisms to discover the first of such mutations that causes a familial form of cancer known as retinoblastoma, fittingly naming the encoded protein the retinoblastoma protein (pRb). 1 Proof that mutations resulting in aberrant gene product from a single gene could cause cancer was revolutionary, and sparked the rush for further discovery of genomic mutations. It took years to recognize that there may be mechanisms outside of traditional genomics underlying cancer development. Eventually, the advent of epigenetics and the discovery of gene regulation via microRNA (miRNA) revealed a multifaceted and complex understanding of cancer development.
miRNAs were discovered in Caenorhabditis elegans in 1993 as regulators of a mechanism by which small RNA transcripts could regulate expression of larger genes with vital cellular functions. 2 Shortly after, many miRNAs were both identified and mapped within the human genome. The potential for miRNA to act in a manner similar to these tumor suppressor proteins was established when miRNA targets were found to be related to fundamental cellular processes such as proliferation, apoptosis, and differentiation, 3 while downregulation of these miRNAs was seen in many types of cancerous tissues.4–7 Our understanding of the complex mechanisms by which miRNA regulates larger cellular processes is incomplete, and the way they act as tumor suppressors is even less clear. Nevertheless, many recent developments have highlighted their critical importance in neoplasia, and investigation into miRNAs’ roles as tumor suppressors is an exploding field of research in modern oncology. Our research and clinical practice focus on myelodysplastic syndromes (MDS). Here, we will analyze recent developments in miRNA research as tumor suppressors as well as discuss future directions for this research using MDS and acute myeloid leukemia (AML), a clinically distinct yet related entity, as models.
MDS
MDS comprise a heterogeneous group of hematologic disorders that arise from a clonal expansion of hematopoietic stem and progenitor cells. As in all cancers, this clonal expansion leads to a competitive advantage over neighboring cells. The clinical effects include refractory anemia, leukopenia, and thrombocytopenia leading to chronic fatigue and transfusion dependence, as well as increased risk of infection and bleeding. There may also be a limited increase in bone marrow blast cells, insufficient to be considered leukemia, with one-third of MDS cases eventually transforming to AML. The cytopenias found in this disease arise from early apoptosis in the myeloid maturation process,8,9 but unlike in leukemia, cells retain their capacity to mature fully. 10 There are many subtypes of this disease, stratified by pathologic findings in peripheral blood samples and bone marrow biopsies, karyo-type abnormalities, and defining mutations. The combination of these factors determines the patient's prognosis, risk for transformation to AML, and response to treatment. However, many factors driving disease phenotype and treatment susceptibility or resistance have yet to be determined. Because of this, approved therapies such as lenalidomide, azacitidine, and decitabine work only in some MDS patients who have the subtypes they are approved for. The complex nature of these syndromes’ underlying causes makes targeted therapy difficult, because even within defined subtypes there is great heterogeneity. The activity of miRNA suggests that they may play an important role in the development of MDS and many other cancers by potentiating the effects of abnormal cellular processes and even driving disease progression as the primary underlying cause, making them additional targets for therapy.
Biogenesis of miRNAs
miRNAs are transcribed as primary RNA (pri-miRNA), ranging from hundreds to thousands of base pairs long with a hairpin-loop secondary structure (Fig. 1). 11 Processing with the nuclear microprocessor complex by the proteins Drosha (an RNase III) and DiGeorge critical region 8 (DGCR8) leaves a hairpin-loop precursor miRNA (pre-miRNA) of approximately 70 base pairs.12–15 These molecules leave the nucleus and are further processed by Dicer, another RNase III, leaving short 18-25 base pair dsRNAs in the cytoplasm. 16
Denaturation to ssRNA allows the active miRNA strand to complex with RNA-induced silencing complex (RISC) to bind complementary sequences on target mRNAs. This complex then uses enzymatic endonuclease activity to silence the target mRNA. 17 miRNA may also act to block or enhance translation by binding to their target mRNA's complementary sequence in the 5’ untranslated region (UTR) and either activating or inhibiting the ribosomal complex.18,19 Mechanisms by which miRNA may act in a cell are still being discovered, though most commonly they act as inhibitors of their target mRNA.
Molecular Mechanisms of miRNAs as Tumor Suppressors
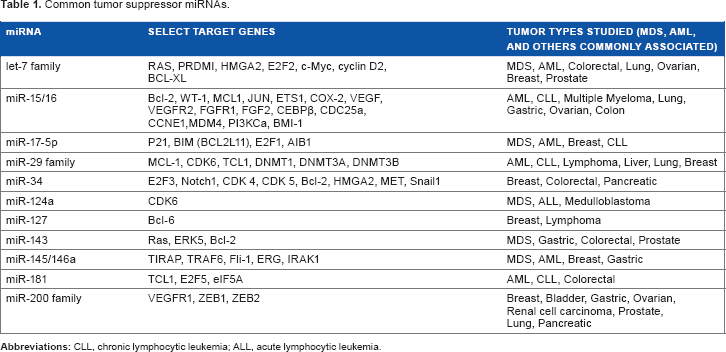
To act as a tumor suppressor, a miRNA must downregulate target mRNA that could otherwise cause a neoplastic or dysplastic phenotype by leading to aberrant proliferation, reversion of differentiation, or increased metastasis. Well-characterized examples of these tumor suppressor miRNAs are becoming increasingly abundant in the literature (Table 1).
Owing to their short length of approximately 21 base pairs, miRNAs have complementary sequences throughout the genome (target multiplicity), leading to an exponentially larger effect on the cell than the suggested small change in miRNA levels. This is important when considering what type of effect miRNAs have on cells when downregulated in cancer or when increasing expression for therapeutic purposes (Tables 2 and 3).
A schematic showing the miRNA biogenesis and mechanism of action. Common tumor suppressor miRNAs.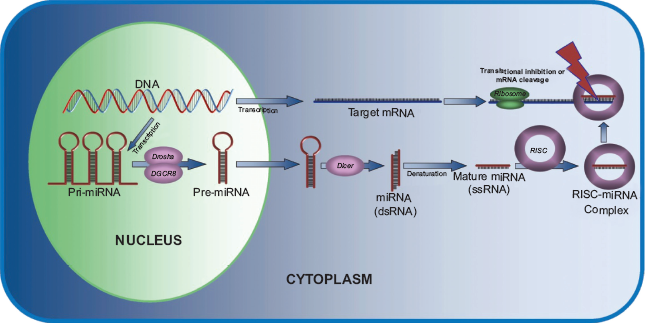
The essential role of miRNA dysregulation in development of myelodysplasia and leukemia was well demonstrated by one group who showed that in mice osteoprogenitor cells (not hematopoietic progenitor cells), knockout of DICER1 led to MDS and AML because of abnormal niche regulation. 20 This finding reinforces the fact that not only are miRNAs important to cancer development and progression of the cells in which they are deregulated, but they can also transform other cell types located regionally or distantly by altering their signaling (Fig. 2). This review will explore this complex relationship with a number of examples showing the different mechanisms by which miRNA may act as tumor suppressors.
miR-145 and miR-146a in MDS, and miRNA as a Primary Driver of Disease
Located near the commonly deleted region (CDR) of chromosome 5 in the del(5q) subtype of MDS, miR-145 and miR-146a are more commonly downregulated in patients who have 5q-syndrome than MDS patients without this deletion.21,22 These two miRNAs have been shown to be mediators of the toll-like receptor (TLR) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-KB) pathways,22,23 which are essential inflammatory signals in MDS. Abnormal niche maintenance has been implicated in development of hematopoietic dysplasia, 20 which may be because of abnormal cytokine release or cell signaling abnormalities, which can be modulated by these two miRNA.
Both of these miRNAs act as tumor suppressors in CD34+ bone marrow (BM) stem cells by targeting TIRAP and TRAF6, respectively, which allows for regulation of NF-KB signaling. When TRAF6 is upregulated or these miRNAs are downregulated, there is constitutive activation of this pathway and development of a del(5q) phenotype. miR-145 is also a regulator of FLI1, which has an important role in the balance between erythroid and megakaryocyte lineage differentiation. 24 The combined hemizygous knockdown of miR-145 and RPS14 (another gene in the CDR) led to an increase in the megakaryocyte/ erythroid ratio, a del(5q)-like phenotype, and thus suggests the important role of miR-145 in this process.
miR-146a has also been shown to be important to the regulation of inflammation, myeloproliferation, and oncogenesis. 25 Knockout of this gene led to hypersensitivity to lipopolysaccharide (LPS), leading to over-production of many cytokines, increase in TRAF6 and IRAK1 protein levels in macrophages, and autoimmunity. Aging null mice also develop MDS, BM failure, and myeloproliferative neoplasms (MPNs). These findings have direct implications for the evolution of MDS and highlight the role of abnormal niche maintenance as an underlying cause of MDS.
miRNA as tumor suppressors in MDS.

miRNA as tumor suppressors in AML, with focus on those relevant to MDS pathogenesis.
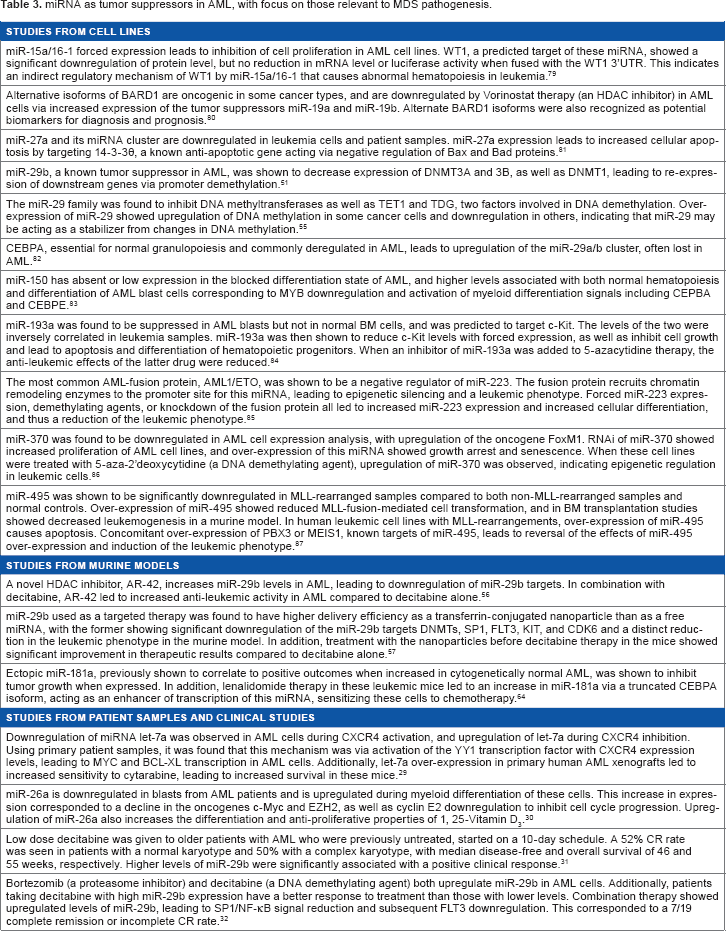
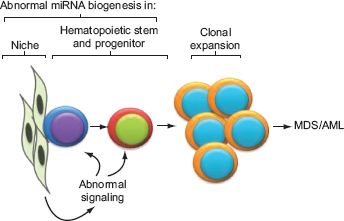
A schematic showing how abnormal miRNA biogenesis or regulation results in MDS/AML. Deregulation of one or more miRNAs in the hematopoietic microenvironment (niche) results in abnormal signaling to hematopoietic cells. Also, deregulation of miRNA within the hematopoietic stem and progenitor cells results in a clonal expansion leading to MDS/AML.
Let-7a in MDS and AML, and the Many Roles of a miRNA
MiRNA array analysis has consistently revealed downregulation of miR-let-7a in MDS, 28 but shows a more variable expression pattern in AML with low levels in some studies and elevated levels in others.23,25,26,29–33 Let-7a is traditionally considered a tumor suppressor, 34 downregulating various oncogenes including all three members of the Ras family, 35 EZH2, 36 and Myc. 37 In a study utilizing in silico modeling of miRNA function, analyzing most of the known miRNAs and their target pathways to relate expression and function, let-7a was near the top of the list of miRNAs having the largest number of targets with 45 types of pathways predicted to be targeted. 38 This indicates a large magnitude of effect within the cell with altered regulation and likely an extreme importance to cellular function.
Further studies have revealed the important role for this miRNA in abnormal hematopoiesis. Downregulation of let-7a leads to upregulation of important genes in carcino-genesis, including c-Myc and BCL-XL in AML cell lines, leading to a worsening leukemic phenotype. 29 Interestingly, the variability of this miRNA in AML indicates a more complex role in AML pathogenesis. Elevated levels were found to be associated with poor outcome, 39 while downregulation increased chemoresistance of tumor cells in vitro. 29 In addition, a recent study in MDS patients showed that lower levels of let-7a in circulation were predictive of a better prognosis in MDS patients, though lower levels were found in MDS patients overall compared to healthy controls with normal hematopoiesis. 40
Although circulating levels do not correlate necessarily to the activity level within a cell, this discrepancy between serum levels and function indicates a poor understanding of how this miRNA acts in hematopoietic cells versus other types of cancer to lead to either MDS or AML development, or in the transition to secondary AML as disease progresses. Virtually all miRNAs have been shown to exhibit target multiplicity within a cell, including the tumor suppressor miRNAs miR-202, miR-101, miR-302b, and miR-29b with 90, 65, 49, and 40 predicted target pathways, respectively. 38 This indicates that tumor suppressor miRNAs have extremely complex regulatory mechanisms in many cancers, and that their many roles will lead them to be difficult yet highly important targets for therapy.
miR-124 in MDS and Epigenetic Suppression
miR-124 is an established tumor suppressor and has been shown to be downregulated in MDS.41,42 This miRNA is involved in a number of cancers, targeting oncogenes such as Slug in metastatic breast cancer, 43 cyclin-dependent kinase 6 (CDK6) in acute lymphoblastic leukemia, 44 and CDK4, cyclin D2, and EZH2 in uveal melanoma. 45 The most well-recognized mechanism for its downregulation in cancer is via epigenetic silencing because of methylation of its promoter.
EVI-1 is an oncogene that acts via epigenetic regulation of the phosphatase and tensin homolog/Akt/mammalian target of rapamycin (PTEN/Akt/mTOR) axis, an important pathway in cancer development. 46 EVI-1 has been implicated in the pathogenesis of MDS,42,47,48 as well as many other types of cancers, and has been shown to be involved in miR-124-3 regulation via promoter methylation.
EVI-1 has been shown to directly interact with DNMT3A and 3B at the miR-124 promoter site, 49 indicating its role in de novo sequence-targeted DNA methylation leading to leukemogenesis. The driver of disease is abnormal regulation of EVI-1; however, it seems that suppression of miR-124 in this pathway ultimately causes the disease phenotype. Promoter methylation as the driver of this suppression was confirmed, and it was shown that this downregulation of miR-124 causes a significant oncogenic phenotype of cell cycle abnormalities, BM failure, and death in mice.42,47 Additionally, it was found that while epigenetic regulation was usually the primary mechanism for miR-124 suppression, it was not the mechanism driving suppression in all diseased individuals, 50 indicating that miR-124 dysregulation may be as important in pathogenesis as EVI-1, and thus just as important as a target for therapy.
Many other miRNAs are involved in epigenetic regulation as well, including miR-29b, 51 let-7a, 36 and miR-26a in MDS and AML, 30 and more in other cancer types. From the study of miR-124, it appears that epigenetic dysregulation of miRNA can alone lead to a cancerous phenotype, and that miRNA changes seen in MDS and AML are often not unique to these diseases, and may be applicable to many cancer subtypes.
miR-29b in AML, and miRNA as Targets of Therapy and as Therapy Themselves
Independently, methylation status and NF-KB signaling are potential mechanisms driving cancer; however, these pathways have been found to be modulated by the tumor suppressor miR-29b in AML pathology. This miRNA downregulates a number of oncogenes including Mcl-1 (a protein in the Bcl-2 family) which regulates the rate of apoptosis, 52 Tcl-1 in T-cell leukemias, 53 and DNMT3A and 3B in both AML and lung cancer,51,54 with a total of 40 predicted target pathways. 37 miR-29b is downregulated in AML, 26 and was shown to act as a driver of the disease phenotype when suppressed because of increased methylation via DNMT1/3A/3B. 51 It was also found to inhibit Ten-Eleven Translocation 1 (TET1) and Thymine-DNA Glycosylase (TDG), both involved in DNA methylation regulation. 55
Most miRNAs that are dysregulated in cancer are potential targets for therapy, including all discussed here, though some miRNAs will work better than others as monotherapy or single drug targets. For example, miR-29b has been found to be specifically targeted by bortezomib and decitabine, commonly available therapies. In AML, decitabine was shown to have a good response as the first-line therapy in a subset of AML patients, and higher levels of miR-29b significantly correlated with a positive clinical response. 31 Combining the two drugs for treatment of elderly AML patients led to increased expression of miR-29b, with corresponding NF-KB signal reduction, FLT3 downregulation, and clinical response in 44% of the patients in the phase I trial. 32 In addition, in experimental therapies on leukemic and patient cell lines and in mouse models, upregulation of miR-29b in combination with decitabine actually improves therapeutic efficacy compared to decitabine alone. This was achieved with the experimental drug AR-42 and with miR-29b conjugated to nanoparticles,56,57 both leading to a reduction in leukemogenesis in their respective models. This indicates that not only are tumor suppressor miRNAs good targets for therapy and markers of response via endogenous upregulation, but they may also be used eventually in a clinical setting by exogenous delivery.
The Future of Therapy Related to miRNA
Available chemotherapeutic drugs often already target miRNA expression as part of their mechanism of action. In MDS, this includes lenalidomide, 27 HDAC inhibitors, 58 DNA demethylating agents like decitabine, 59 and other chemotherapeutic agents across all cancer types. However, more specific targeting of the miRNA identified as tumor suppressors and identifying the signaling pathways they affect for further drug development are the ultimate goals for personalized therapy to improve patient care.
To achieve this goal, we must consider which miRNAs are drivers of the malignant phenotype and how we want to upregulate that tumor suppressor activity. There are a number of mechanisms being implemented now which use exogenous miRNA in the lab to directly treat cancer cells. Two well-studied models are (1) attaching the target miRNA to a molecule that stabilizes it in the blood and leads to improved tissue delivery, commonly studied as conjugation to liposomes or other more advanced nanoparticles,60,61 or (2) delivery using a virus (commonly adenovirus, though other models exist) that contains the target miRNA of interest in its genome. 62 Both methods have advantages and disadvantages, and need much more testing before reaching clinical practice.
Resistance to chemotherapy has also been found to be mediated by miRNA and their target oncogenes. One group showed that downregulation of miR-181a and subsequent upregulation of Bcl-2 in leukemia cells drove chemoresistance to Ara-C, 63 one of the most common drugs used in AML. The introduction of this miRNA into the cells increased chemosensitivity via apoptosis. Additionally, miR-181a upregulation after lenalidomide therapy led to chemotherapy sensitization to Ara-C in a subset of AML patients. 64 This type of miRNA treatment, as adjunct therapy for other chemotherapeutic agents, is a mechanism that has therapeutic implications in clinical practice.
Levels of miRNA in patients with cancer can also be helpful, especially to determine responsiveness to treatment or relapse of disease, as discussed previously related to let-7a. 40 Ezatiostat treatment in low-risk MDS was found to be more effective in patients with low miR-129 and high miR-155 expressions before therapy. 65 Both of these studies indicate the variability and utility that tracking miRNA levels can offer.
Although the use of natural agents for therapeutic purposes has existed for centuries, the molecular mechanisms of their efficacy have only recently been investigated. The potential use of such substances in combination with other drugs for cancer therapy is an active area of research. Green tea contains a compound known as epigallocatechin gallate (EGCG) that was recently shown to increase expression of the tumor suppressor miR-16. This subsequently suppresses Bcl-2 in hepatocellular carcinoma cells, leading to apoptosis. 66 Similar results were found for curcumin, a compound found naturally in turmeric (a spice that has been praised for its anti-cancer properties). This compound blocks the NF-KB pathway to lead to increased apoptosis in prostate cancer cells, by lowering Bcl-2 expression and increasing procaspase-3 and procaspase-8 levels, 67 as well as downregulating a number of oncogenic factors and growth factors in breast cancer cells. 68 It has since been shown to have a large role in altering expression of miRNA in cancer cells. 69 Curcumin was also shown to decrease WT1 levels in leukemic cell lines and primary AML cells via upregulation of miR-15a/16-1. 70 Researchers are even synthesizing analogs to study their effects on miRNA to strengthen this therapeutic effect. 71 EGCG and curcumin are only two of many natural compounds being studied for their anti-cancer properties, making this an exciting young field in the search for drugs that can increase treatment efficiency.
Conclusion
While numerous miRNAs act as tumor suppressors, and more are likely to be identified, we have limited our review to those that have been shown to have a modulating role on pathways known to be deregulated in MDS and AML. The initial realization that miRNAs were active in hematopoiesis opened an exciting new avenue for drug development. AML and MDS are both diseases that desperately need new approaches for therapy, as a majority of patients are either resistant to current therapies or eventually relapse. The studies outlined here provide insight into how knowledge of miRNA expression and function in cancer evolution can be translated into developing targeted therapies. First, miRNA dysregulation may act as part of the larger mechanism underlying what is driving the cancerous phenotype, and may also be the primary driver of the disease. Second, miRNA can cause an oncogenic phenotype, and can also act as mediators of chemoresistance and chemosensitivity in cells, both of which are important therapeutic targets. Third, altered epigenetics such as DNA methylation status and chromatin alteration often play a role in miRNA dysregulation in cancer. Fourth, miRNA signatures or circulating miRNA levels may act as markers for personalization of therapy or prognostic determination. Fifth, miRNA can be targeted to improve cancer therapy by agents that are already in the market or are in development, and also may eventually be used as therapeutic agents themselves. Finally, natural agents are currently being explored for their effects on oncogenesis related to miRNA expression, and are showing promising results for future use as isolated natural compounds or as analog therapies.
These discoveries lead to the conclusion that tumor suppressor miRNAs are essential mediators of cancer development and will undoubtedly be crucial targets for cancer therapy and clinical practice in the future. The complexities of miRNA regulation of target oncogenes, and their effects on subsequent signaling pathways and disease phenotype, are under intense investigation to develop a more comprehensive model. This is especially important in light of the multiple target genes that can be modulated by a single miRNA. However, tissue or cell-type specificity may minimize some of the unwanted side effects. miRNAs are the new weapons in the arsenal of the fight against this extremely complex disease called cancer.
Author Contributions
NI researched and wrote the first draft of the manuscript. NI, NG, AA, and AR contributed to the writing of the manuscript, agreed with manuscript conclusions, jointly developed the structure and arguments for the paper, and made critical revisions and approved the final version. All authors reviewed and approved the final manuscript.
Footnotes
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests. Provenance: the authors were invited to submit this paper.