
Abstract
Background and Purpose
Transmissible spongiform encephalopathy (TSE) diseases are fatal, progressive neurodegenerative disorders affecting both humans and animals. Clinical signs typically appear after years and even decades of silent disease progression. This study was aimed at investigating whether altered brain MRI patterns may precede clinical signs in a TSE rodent model.
Methods
In vivo T2-weighted (T2W) MRI examinations (4.7 T) were performed on Golden Syrian hamsters (GSH) intracerebrally, orally, or intraperitoneally (i.p.) infected with the 263K scrapie strain. Histopathological analyses were performed on i.p. infected GSH at the end of one-day or longitudinal MRI sessions.
Results
T2W-MRI hyperintensity was detected in the thalamic nuclei of GSH with clinical signs, irrespective of the infection route. Hyperintensity in the thalamus was also observed in pre-clinical animals, between 106 and 121 days post-infection (dpi), while normal T2W intensity was detected in four animals examined between 72 and 96 dpi. Pathological prion protein deposition (but no astrogliosis and only occasionally, weak spongiosis) was detected between 106 and 121 dpi.
Conclusions
The altered T2W-MRI pattern detected in the thalamus of asymptomatic i.p. infected GSH provides a useful basis for evaluating the effectiveness of possible therapeutic approaches at early stages of TSE disease.
Introduction
Transmissible Spongiform Encephalopathy (TSE) or prion diseases are fatal, progressive neurodegenerative disorders affecting both humans, e.g. Creutzfeldt-Jakob disease (CJD), and animals, e.g. scrapie in sheep and goats, and bovine spongiform encephalopathy (BSE) in cattle. 1 Clinical signs of TSE diseases may take months, years, or even decades to develop. A distinct form of human TSE diseases, generally affecting young patients, is variant CJD (vCJD), which has been linked to the consumption of BSE contaminated meat products or, in a few cases, to secondary transmission by infected blood transfusion.2,3 TSE diseases are typically characterized by a triad of neuropathological changes, i.e. neuronal loss, hypertrophy and hyperplasia of astrocytes and microglia, and spongiform degeneration associated with accumulation of the pathological prion protein (PrPSc). This latter is a misfolded isoform of a host-encoded glycoprotein named PrPC, located at the cell surface through a glycosyl-phosphatidylinositol anchor (GPI), mainly expressed in the central nervous system (CNS), but also present in other organs and tissues. TSE diseases may occur in sporadic, infectious or genetic forms. Several allelic forms of PrP are linked to disease susceptibility in mice, sheep, and man. Mutations of the human PrP gene (PRNP) have been found in familial TSE cases and the polymorphic site at codon 129 (coding either for methionine or valine) is a genetic risk factor for sporadic and variant CJD.
Neuro-imaging has played an increasingly important role in the diagnosis of human prion disease. Magnetic Resonance Imaging (MRI) has a relevant position in the diagnostic criteria to differentiate vCJD from other forms of CJD. 4 The characteristic hyper-intensity of the pulvinar and dorso-medial thalamic nuclei, also referred as the “hockey stick” sign, 5 detected by T2-weighted (T2W), fluid attenuated inversion recovery, proton density, and diffusion weighted MRI sequences represents a highly sensitive (90%) and specific (96%) diagnostic indicator of the disease, and was entered as one of the recommendation criteria for the diagnosis of vCJD.4,6–9
The prolonged asymptomatic phase of TSE is the reason for numerous studies aimed at identifying affected individuals at early stages of the pathogenesis.10,11 Non invasive methods, such as brain MRI, able to identify patients during early or pre-clinical phases of disease, may allow treatment with a better chance of success. 12
MRI allows non invasive longitudinal monitoring of signal changes in the brain of TSE animal models13–15 and therefore may contribute to identify early indicators of disease progression and clarify the underlying pathological mechanisms. Vacuolation of neurones/neuropil and gliosis were reported to respectively correlate with hypo-intense and hyper-intense changes in brain T2W MRI images of a hamster model of scrapie. 13 Furthermore, increased signal intensity was observed in T2W MRI of brain areas presenting accumulation of PrPSc and astrogliosis in a murine model of prion disease. 14
On the basis of these findings, the objective of the present study was to evaluate, in a rodent scrapie model, whether in vivo MRI would be able to discriminate between infected and uninfected animals before the onset of clinical signs and to correlate possible signal alterations with specific brain lesions.
Materials and Methods
Scrapie Infection Models
The hamster-adapted 263K scrapie strain was used in consideration of its short incubation period (IP) and the extensive experience gained on the 263K-infected hamster model. 16 Weanling female Golden Syrian hamsters (GSH, Charles River, Como, Italy) were inoculated with a 10% (wt/vol) suspension in physiological saline of pooled brain homogenates derived from clinically affected hamsters infected with the 263K scrapie strain. Hamsters were infected intracerebrally (i.c., 0.05 ml), intraperitoneally (i.p. 0.10 ml), or orally (per os, 0.10 ml). Groups of 8–10 hamsters were housed in cage with food and water ad libitum, scored 5 days a week for early (increased alertness to noise or irritability and aggressiveness when handled) and late clinical signs (tremor of head, wobbling gait and spontaneous backrolls) verified by two independent observers five days per week as previously described. 17
The research protocol was approved by the Service for Biotechnology and Animal Welfare of the Istituto Superiore di Sanità and authorized by the Italian Ministry of Health, according to Legislative Decree 116/92, which in Italy has implemented the European Directive 86/609/EEC on laboratory animal protection.
MRI
In vivo MRI examinations were performed at 4.7 T on a Varian Inova SIS 200/183 system (Varian, Palo Alto, CA, USA) utilizing either a 4.0 cm internal diameter Litz volume coil (Doty Scientific, Inc, Columbia, SC, USA) or a 2.5 cm diameter surface coil.
T1-weighted (T1W) scout imaging was obtained by a spin echo sequence with TR/TE = 450/16 ms (4 scans, slice thickness 1 mm; FOV = 4 × 4 cm2; matrix 128 × 256). A spin echo sequence with TR/TE = 4000/90 ms (4 scans; slice thickness 1 mm; FOV = 4 × 4 cm2; matrix 128 × 256) was utilized for T2-weighted (T2W) imaging.
MRI signal intensity was evaluated by two independent investigators, using a dedicated Varian image browser program.
For MRI examinations animals were anesthetized with 1%–2% isoflurane in a mixture of O2:N2O (60:40), 1 L/min. The animals were sacrificed with CO2 at the end of the last scheduled MRI examination.
Histopathology
Histopathological analyses were only performed on i.p. infected hamsters. At post-mortem, brains were fixed for 5–7 days in 10% buffered formalin. They were subsequently trimmed at standard coronal levels, decontaminated with formic acid for 1 h, and embedded in paraffin. Six-μm-thick brain sections were serially cut (3 sections every 100 μm), for haematoxylin/eosin staining, PrPSc and glial fibrillary acid protein (GFAP) immuno-histochemistry.
PrPSc immunohistochemistry was carried out as previously described. 18 Briefly, sections were hydrated, pre-treated with 98% formic acid for 1 min, submitted to hydrating autoclaving for 30 min at 121 °C, and finally left cooling overnight. Incubation with antibody, avidin-biotin complex treatment and antigen detection, were carried out with Dako-Autostainer (Dako-Cymation). Sections were treated with 6% normal goat serum (Vector, Burlingame, California, USA) in PBS for 30 min. Immunohistochemical detection of PrPSc was performed with mAb 3F4 (SIGNET, Emeryville, California, USA) at 1:100 in PBS with 3% normal goat serum (Vector) for 45 min. After washing with PBS, sections were treated with ABC Complex (Vector) for 45 min and with diaminobenzidine (Dako-Cytomation) for 7 min. Sections were finally counterstained with Mayer's haematoxylin. Adjacent sections were stained with haematoxylin/eosin for the evaluation of spongiform changes.
For GFAP immunolabelling, deparaffinized sections were treated with 3% H2O2 in methanol for 10 min at room temperature to abolish endogenous peroxidase activity. After heat-induced antigen retrieval (citrate buffer 0.01 M, pH 6.0, 1 × 5 min in microwave oven, 750 watt), the sections were incubated for 30 min with polyclonal rabbit anti-GFAP (1:500, Dako-Cytomation). Immunoreactivity was detected with ABC method (Dako-Cytomation) as described above. In Table 1, the intensity of lesions in the thalamus is reported as the mean of four different nuclei (lateral, ventral, mediodorsal, and central medial), which were scored from 0 (no lesion) to 4 for spongiosis and presence of PrPSc and from 0 to 3 for GFAP staining.
MRI and histopathology examinations in the thalamus of hamsters infected with the 263K scrapie strain.

Neuropathological features were blindly scored by two independent observers.
Results
In this study attention was focussed on MRI signal alterations occurring in the brain of scrapie infected hamsters, at the level of the thalamic nuclei. In this brain region most of the previous studies consistently showed T2W imaging signal hyperintensity in symptomatic hamsters infected by the scrapie strain 263K.13,19,20 MRI signal changes were also reported for other brain areas (such as cortex, corpus callosum and hippocampus), but these alterations were not always simultaneous and were only occasionally found in limited percentages of infected animals during the clinical phase. Therefore, these brain areas were not selected for our study, which was aimed at monitoring reproducible MRI changes in relation to the appearance of clinical signs.
MRI examinations were performed on a total of 14 hamsters infected with the 263K by different routes and 14 healthy age-matched controls. The time schedule of MRI examinations on scrapie-infected hamsters, including that of two i.p. infected hamsters (#7 and #8) which were longitudinally monitored at different days post-infection (dpi) is reported in Table 1.
MRI Alterations in the Thalamus of Scrapie Infected Hamsters Showing Clinical Signs
MRI experiments were performed on three groups of hamsters, all presenting clinical signs: i.c. infected GSH examined at 70 dpi, three GSH infected per os and examined at 158, 179 and 180 dpi, and two i.p. infected GSH which were longitudinally monitored at 120 and 124 dpi and at 119 and 131 dpi, respectively. A schematic representation of brain areas under investigation is reported in Figure 1. T2W signal hyperintensity was detected in the thalamic nuclei of all these symptomatic hamsters (Table 1), irrespective of the route of scrapie infection examples in Figure 2A (i.c. inoculation), 3A (per os) and 4A (i.p.), compared with the respective controls (panels 2B, 3B and 4C).
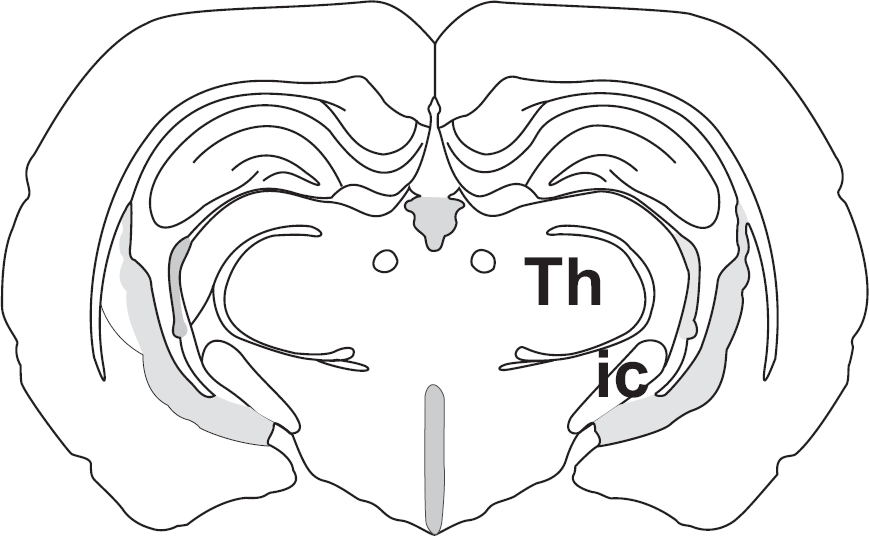
Schematic representation of brain areas under investigation. IC: internal capsula; Th: thalamic nuclei. Stereotaxic Atlas of The Golden Hamster Brain. Lawrence P. Morin and Ruth I. Wood. Academic Press.
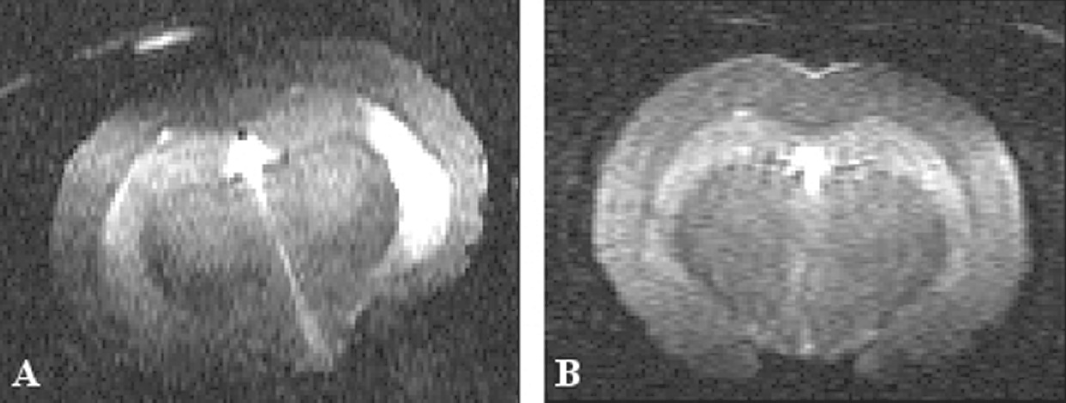
Coronal T2W MR images of golden syrian hamsters intracerebrally infected with the 263K scrapie strain and examined in the clinical phase (70 dpi). Signal hyperintensity was consistently detected in the thalamic nuclei of infected animals (example in panel A), compared with negative controls (example in B).
No signal alteration was observed in T1W images of any of these animals (data not shown).
Histopathology, carried out only on the two i.p. infected hamsters at the end of longitudinal MRI examinations, showed PrPSc immunostaining in both animals and spongiosis in one, without evidence of astrogliosis (Table 1). PrPSc deposition was characterized by a diffuse and punctuated pattern (Fig. 5).
T2W Signal Hyperintensity in the Thalamus of I.P. Infected Hamsters before the Appearance of Clinical Signs
Peripherally (i.p.) infected hamsters were selected for investigating MRI alterations in infected asymptomatic animals, to avoid any possible imaging artefact associated with brain lesions and/or scars produced by i.c. inoculation.
Eleven MRI sessions on eight asymptomatic hamsters examined between 72 and 121 dpi (including two hamsters longitudinally monitored at 72, 93 and 107 dpi and at 96 and 110 dpi, respectively, and six animals subjected to single examinations between 84 and 121 dpi) showed: a) normal MRI patterns in four animals monitored between 72 and 96 dpi (five examinations); b) T2W signal hyperintensity in six animals examined between 106 and 121 dpi (six examinations). An example of signal hyperintensity in thalamic nuclei of an asymptomatic GSH is shown in Figure 4B, compared with that of an animal with clinical signs (Fig. 4A) and a control (Fig. 4C). Interestingly, two hamsters (#7 and #8) that respectively showed normal scans at 93 and 96 dpi, exhibited hyperintensity in the thalamus when examined again two weeks later, still in the absence of clinical signs (Table 1). These hamsters were kept alive for further MRI evaluation in the clinical phase (see Table 1 and previous section).
The signal hyperintensity was evaluated by referring the T2W signal intensity of the thalamus to that of the internal capsula. For the hyperintense thalamic nuclei of i.p. infected animals this ratio was 1.24 ± 0.05 (mean ± SD; range 1.19–1.29, n = 6), a value exceeding by more than two standard deviations the mean value of normal appearing thalamic areas (1.11 ± 0.04, n = 4).
Necroscopy and histopathology, performed on three asymptomatic hamsters with no T2W signal hyperintensity (#9, #10, #12; Table 1), showed some PrPSc deposition (but no GFAP nor spongiosis positivity) in only one (hamster # 12, 111 dpi). Regarding asymptomatic hamsters that exhibited T2W MRI hyperintensity in the thalamus, three showed PrPSc deposition (#11, 106 dpi; #13, 111 dpi and #14, 121 dpi), while a weak spongiosis was only detected in one (#13).
Discussion
Aim of this study was to investigate, whether and to which extent changes in MRI signal intensity may precede the onset of clinical signs in laboratory rodents infected with scrapie. An answer to this question might pave the way to further developments of non-invasive imaging approaches for an early diagnosis of the disease and to the possible assessment, under controlled experimental conditions, of the effectiveness of potential therapeutic approaches in retarding disease progression from asymptomatic into the symptomatic-phase and prolonging survival.
MRI experiments performed in our laboratories on GSH i.p. injected with the 263K scrapie strain showed that areas of T2W signal hyperintensity appeared in the thalamus not only in the clinical phase, but also in the absence of clinical signs. The T2W imaging hyperintensity consistently detected in the thalamus of symptomatic hamsters infected by either i.c. or peripheral routes, is in agreement with similar findings reported for i.c. 263K scrapie-infected hamsters examined at the same (4.7 T) or lower magnetic field (1.5 T)13,20 as well as for CD-1 mice i.p. infected with the mouse-adapted 139A scrapie strain, examined by in vivo microimaging at 9.4 T. 14 Differently from these reports, no T2W imaging changes were observed by Dubois et al 21 in symptomatic 263K scrapie infected hamsters.
No T1W changes were observed in any of our animals, also in agreement with previously reported results. 13
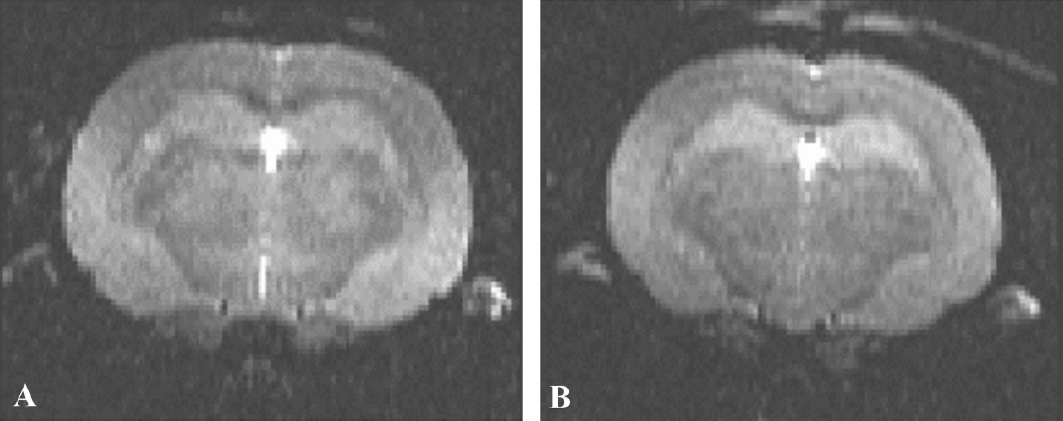
Coronal T2W MR images of golden syrian hamsters orally infected with the 263K scrapie strain and examined in the clinical phase (180 dpi). Signal hyperintensity was detected in the thalamic nuclei of all infected animals (example in panel A), compared with negative controls (example in B).

Coronal T2W MR images of golden syrian hamsters intraperitoneally infected with the 263K scrapie strain and examined in either the clinical phase (panel A; hamster #7, 124 dpi) or before the onset of overt clinical signs (panel B; hamster #13, 111 dpi). Signal hyperintensity was detected in the thalamic nuclei of both infected animals, compared with negative controls (example in C).

Histological and immunohistochemical analysis of ventral thalamus from an i.p. infected hamster with no clinical signs (#13, 111 dpi). Serial sections of the ventral thalamus were analysed by haematoxylin and eosin staining (A) immunohistochemistry for PrPsc (B) and immunohistochemistry for GFAP (C) Bar represents 100 μm.
Regarding the MRI alterations observed in our laboratory in the thalamus of i.p. 263K-infected hamsters during the pre-clinical phase (six/six GSH examined between 106 and 121 dpi), it is worth mentioning that similar changes in the T2W signal intensity were also observed (and reported as unpublished results) by Forloni et al 20 in the thalamus of asymptomatic hamsters intracerebrally infected with the same scrapie strain.
Consistently with these MRI findings, the histopathological analyses performed in our study showed that diffuse and punctuated patterns of PrPSc deposition were already prominent in the thalamic nucleus, as well as in the geniculate nucleus and the adjacent internal capsule, in the late pre-clinical phase. PrPSc deposition in the thalamus, however, was not associated with astrocytosis, and in only one animal there was modest spongiosis. Therefore, on the basis of the clear prevalence of only one of the three typical neuropathological changes, we suggest that T2W hyperintensity in the thalamus of pre-symptomatic animals is likely due to PrPSc deposition.
A proposed interpretation of the existing relationships between MRI signal alterations and neuropathological and behavioural abnormalities suggests that the T2W hyperintensity in MRI images is mainly produced by concurrent effects of PrPSc deposition. The similarity of MRI signal intensity changes observed in our study in the thalamus of GSH seems to be in agreement with the low level of spongiosis observed in our model system at both stages of the disease.
It is also worth noting that in our study areas of T2W-hyperintensity were typically detected in symptomatic animals, irrespective of the route of scrapie infection (i.p., i.c. or per os), supporting the appropriateness of the 263K scrapie model for longitudinally monitoring MRI biomarkers of progressive disease. In this context, the i.p. model is preferable to other infection routes, for avoiding scar artefacts and offering a still acceptable incubation time.
A useful comparison of our results with those reported by other studies similarly focussing on MRI signal alterations in the early clinical phase of scrapie infected rodents (Table 2) requires awareness of the different regional patterns of PrPSc deposition and astrogliosis and also of the different relationships between astrocytosis and amyloidosis, which may occur in the brain of animals infected with different scrapie strains. For instance, while PrPSc deposition was observed in most brain areas of 263K-infected hamsters, such as the thalamus, the cortex and the hippocampus, it was mainly detected in the cerebral cortex and in the hypothalamic paraventricular and supraoptic nuclei in 139H-infected hamsters. 19 Furthermore, PrPSc was also localized with GFAP in reactive astrocytosis in certain areas such as hippocampus in 263K but not in 139H infected hamsters. 19 It is therefore not surprising that other TSE models obtained by using different species of rodents, such as mice infected with different scrapie strains may also result into different patterns of PrPSc deposition, gliosis and neuronal loss and therefore into different patterns of MRI signal changes in different brain regions. Interesting examples of these differences are provided by a recently reported study on C57BL/6J mice intracerebrally infected into the dorsal hippocampus with the ME7 scrapie strain. 15 In this model there was only a decrease in the apparent diffusion coefficient (ADC), but no significant alterations in T1 or T2 relaxation times at the level of any investigated brain area (thalamus, ventral hippocampus or cerebral cortex) at a later stage of the preclinical period (18 weeks post infection).
T2W MRI signal alterations in brain areas of asymptomatic and symptomatic TSE-infected rodents.

N, numbers of infected animals;
in parenthesis, number of examinations.
Beyond these differences, it is interesting to note that the majority of the reported MRI studies were capable to detect T2W alterations in the thalamus of infected rodents in late or early stages of disease. These early changes in MRI signal intensity in the thalamus were also typically detected after the onset of clinical signs, indicating that the combined effects of different pathological mechanisms associated with the progression of scrapie infection (i.e. PrPSc deposition) result in the persistence of the MRI signal changes across the transition from pre-symptomatic to symptomatic stages of the disease.
In conclusion, a substantial body of evidence based upon our results and those of other research groups, point to the possibility of detecting early MRI changes in the thalamus of early symptomatic TSE-infected hamsters. This finding offers a relevant tool to pre-clinical investigations on the effectiveness of possible strategies of treatment of these gloomy and still fatal neurodegenerative disorders.
Footnotes
Acknowledgments
This work was supported by the EU Project N. QLK4-CT-2002-01763 and by the EU Project Neuroprion Network of Excellence (FOOD-CT-2004-506579). F.S. has been a three-year fellow of the EU Project N. QLK4-CT-2002-01763 at the Istituto Superiore di Sanità, Rome. Mr. Massimo Giannini and Mrs. Nadia Palazzini are gratefully acknowledged for excellent technical assistance.
Disclosures
The authors report no conflicts of interest.