
Abstract
Natural killer (NK) cells are a crucial part of the innate immune system and play critical roles in host anti-viral, anti-microbial, and anti-tumor responses. The elucidation of NK cell biology and their therapeutic use are actively being pursued with 200 clinical trials currently underway. In this review, we outline the role of NK cells in cancer immunotherapies and summarize current noninvasive imaging technologies used to track NK cells in vivo to investigate mechanisms of action, develop new therapies, and evaluate efficacy of adoptive transfer.
Introduction
This review focuses on current methods to track natural killer (NK) cells used in cancer therapy: how they can be monitored in vivo to assess tumor response, assay treatment efficacy, and elucidate mechanisms of NK cell-based immunotherapy. NK cell therapy includes activation of endogenous NK cells, and adoptive transfer of activated and genetically modified NK cells. Adoptive transfer of ex vivo-targeted NK cells has been used more recently. Typically, response to adoptive cell therapy is evaluated on the basis of decreases in tumor markers and tumor size and improved survival that are assessed weeks to months after administration of treatment. Localization and function of adoptively transferred immune cells at the tumor site are typically determined through biopsy and ex vivo analysis. Accumulation in the tumor region is one of the requirements for effective adoptive immunotherapy. With the biology of NK cells continually being elucidated, tumor targeting of NK cells can potentially be enhanced using a variety of new methods.
NK cells may be labeled with different markers for in vivo monitoring. 1 Cells can be labeled directly by harvesting them and labeling them ex vivo with fluorophores, radiotracers, or paramagnetic nanoparticles that allow visualization by optical microscopy, positron emission tomography/single-photon emission computed tomography (PET/SPECT), and magnetic resonance imaging (MRI), respectively. Direct labeling procedures may prove to be useful for clinical translation because of the ease of labeling procedures and the potential to use labels that are already approved for clinical use. This method, however, has two disadvantages. First, the level of labeling depends on the capacity of the cell to retain the label, as different cell populations may exhibit different levels of phagocytosis or have different membrane properties. Second, the direct method can be useful for in vivo imaging of only terminally differentiated cells, such as NK cells, dendritic cells, and macrophages, because the label may be lost or diluted as cells proliferate or die. Cells may also be labeled indirectly ex vivo where cells are transduced with a vector carrying a reporter gene. Signal can be generated and tracked in vivo when the reporter gene is expressed and when a transgene-specific probe is administered. Although genetic manipulation makes it possible to track the long-term fate of a cell population (distribution, proliferation, and survival) in vivo, insertion of reporter genes demands stable genetic modification and is currently restricted to preclinical research.
Noninvasive imaging technologies are now able to qualitatively and quantitatively detect the presence of labeled NK cells in target tumors. These imaging signals can potentially be used as real-time biomarkers for tumor response and for differentiating patients who are responders or nonresponders to NK cell therapy. Noninvasive NK cell imaging has the potential to provide immediate evaluation of NK cell therapy in both preclinical and clinical realms.
NK Cells
NK cells are a crucial part of the innate immune system that were originally identified based on their ability to lyse malignant and infected cells without prior sensitization or immunization. 2 NK cells mediate the suppression of infected and tumor cells through several effector mechanisms (e.g. the perforin/granzyme-containing granule, death-receptor and interferon-γ (IFN-γ) mediated pathways, and antibody-dependent cell-mediated cytotoxicity (ADCC)). 3 NK cells produce cytokines that have proinflammatory and immunosuppressive effects (e.g. IFN-γ, tumor necrosis factor-α (TNF-α), or interleukin IL-10) and growth factors, such as granulocyte macrophage colony-stimulating factor (GM-CSF) and granulocyte colony-stimulating factor (G-CSF)). They also produce many kinds of chemokines that are crucial in NK cell trafficking to lymph nodes and areas of inflammation, as well as their colocalization with dendritic and other hematopoietic cells.4,5 NK cell-mediated cytotoxicity and cytokine production provide regulatory roles of NK cells that impact members of the adaptive immune system, such as dendritic cells, macrophages, neutrophils, and T and B cells. 4
Human NK cells are broadly defined as CD3– CD56+ cells and can be further divided into two subsets based on the level of their expression of CD56. 6 CD56bright NK cells are predominantly involved in immunoregulation by producing cytokines. CD56dim NK cells are mainly cytolytic. CD56 NK cells also express high levels of low-affinity Fc receptor for IgG (FcγIIIR–-CD16), which mediates ADCC. NK cells are widely distributed throughout lymphoid and nonlymphoid tissues, including bone marrow, lymph nodes, blood, liver, spleen, and lung. 5 About 90% of NK cells in the peripheral blood and spleen are CD56dim CD16+ NK cells. Most NK cells in the lymph nodes are CD56brightCD16dim/-. 7
NK cells are equipped with a variety of receptors that stimulate or inhibit activity. NK cell activation is dependent on the balance between inhibitory and activating receptors. Activating receptors interact with soluble ligands like cytokines and cell surface antigens. Human activating receptors include the natural cytotoxicity receptors (NCR–NKp46, NKp30, and NKp44), C-type lectin receptors (CD94/NKG2C, NKG2D, NKG2E/H, and NKG2F), and killer cell immunoglobulin-like receptors (KIRs) (KIR-2DS and KIR-3DS). Inhibitory receptors include C-type lectin receptors (CD94/NKG2A/B) and KIRs (KIR-2DL and KIR-3DL). 8 The NK cell receptor repertoire provides three ways by which NK cells take part in tumor immune surveillance. First is the “missing self-recognition” mechanism. At steady state, normal cells express major histocompatibility complex (MHC) class I “self” molecules that bind to inhibitory receptors and inhibit NK cell killing. Under stress conditions, MHC class I is down-regulated in target cells resulting in loss of inhibitory signals and activation of NK cells. Second is the “non-self recognition” process. NK cells possess activating receptors that recognize molecules that are pathogen-encoded, only expressed by infected cells, and are not expressed by the host. For example, in mice, the cytomegalovirus-encoded m157 ligand is recognized by the activating receptor Ly49H in NK cells. Human NK cell activating receptor NKp46 can recognize hemagglutinins on virus-infected cells and initiate NK cell-mediated lysis.9–13 Third is the “stress-induced self” process, wherein NK cells recognize proteins up-regulated only by transformed or infected cells. Furthermore, IFNs and cytokines (IL-2, IL-12, IL-15, IL-18, and IL-21) can potentiate these surveillance roles by enhancing activation, increasing cytolytic activity and promoting maturation of NK cells.4,8
NK Cells in Tumor Immunotherapy
NK cells play crucial roles in the innate and adaptive immunities against infections and tumors. Their therapeutic use is currently being pursued in cancer immunotherapy.8,14 Briefly, there are five methods in which NK cell therapy is being employed pre-clinically and clinically.
Autologous NK cells can be activated and potentiated through systemic administration of cytokines (IL-2, IL-12, IL-15, IL-18, IL-21, and type I IFNs).4,15–18 However, poor clinical outcome has been observed with severe side effect in cancer patients, including vascular leak. Ex vivo activation and expansion of autologous NK cell using cytokines and eventual adoptive transfer back to the patients have shown greater success.
8
Use of allogeneic NK cells in adoptive cell transfer has shown KIR mismatch and enhanced tumor-killing ability. KIR mismatch contributes to alloreactivity of NK cells through the “missing self-recognition” process, i.e. the inhibitory KIRs on allogeneic NK cells do not detect self MHC class I ligands.19,20 Following transfer of haploidentical allogeneic NK cells in acute myeloid leukemia (AML) patients, 26% had complete remission and most patients experienced NK cell expansion following IL-2 therapy.
19
To take advantage of the alloreactivity of NK cells, they are expanded and activated in vitro prior to adoptive transfer using various cytokines (IL-2, IL-15, or IL-21) and growth factors.21,22 NK cell function can also be enhanced by blocking inhibitory KIR with monoclonal antibodies.
23
ADCC: NK cells expressing the activating receptor type IIIA Fc receptor (CD16) recognize antibodies on target cells and trigger NK cell-mediated ADCC. The use of monoclonal antibodies with or without immunocytokine stimulation of endogenous or adoptive NK cells could potentiate ADCC and improve immunotherapy. Antibody treatment of non-Hodgkin lymphoma (with anti-CD20–-Rituximab) and metastatic breast cancer (with anti-transtuzumab/Herceptin) shows strong evidence that ADCC is NK cell mediated.24–26 NK cell lines: Ex vivo-activated NK cells have increased cytolytic activity compared to parental NK cells. This has led to the use of cytotoxic NK cells lines that can be continuously expanded, maintained using good manufacturing practice (GMP), and kept readily available without a donor. There are seven established NK cells lines: NK-92, YT, NKL, HANK-1, KHYG-1, NK-YS, and NKG.8,27 The human NK-92 cell line originated from a patient with large granular lymphoma and is the only cell line that has been shown safe in adoptive transfer in humans.28,29 NK-92 cells can also be expanded in recombinant IL-2. More recently, Tonn et al showed that infusions of ex vivo IL-2-activated NK-92 cells were well tolerated in patients with advanced cancer and remain in circulation up to 48 hours.
30
Genetic modification: In addition to mediating cytotoxicity of NK cells, it is also important to optimize accumulation and retention of the modified NK cells at the tumor site by altering the targeting receptor milieu. Small interfering RNA (siRNA)-based technologies can be used to down-regulate the expression of inhibitory NK cell receptors or overexpress activating receptors.31–33 Gene transfer of chimeric tumor-antigen-specific receptors in NK cells enhance their tumor targeting and killing abilities. Studies in vitro and in vivo have shown that genetic modification of NK cells via transfer of chimeric receptors targeted against HER2/neu, carcinoembryonic antigen (CEA), CD33+ leukemia, and epithelial cell adhesion molecules (EpCAM)-positive prostate cancer provided specific cytotoxicity.34–39 Genetically modifying NK-92 cells with CD20-specific chimeric antigen potentiated NK cell cytotoxicity toward a CD20 expressing lymphoma that was previously resistant to NK cells.
40
Multimodal immunotherapy promises to optimize NK cell efficacy and tumor targeting. 15 NK cell therapy can be combined with cytokines, hematopoietic growth factors, humanized monoclonal antibodies, immunocytokines, and/or immunomodulatory drugs.4,15–18,41,42 Other types of cancer treatments may provide some tumor priming that could also enhance direct or indirect NK cell-mediated tumor recognition and killing. Ionizing radiation and chemotherapy may lead to increased expression of NKG2D ligands on tumor cells and increased NK cell targeting and binding of NKG2D activating NK cell receptors. 43 Drugs acting on histone deacetylase inhibitors or proteasome inhibitors may up-regulate expression of TNF-related apoptosis-inducing ligands on malignant cells and render them more susceptible for TRAIL-mediated NK cell-induced death.44–47 Focused ultrasound treatment of human breast cancer increases tumor-infiltrating lymphocytes that include: CD3, CD4, CD8, CD4/CD8 T cells, B cells, and NK cells. Levels of FasL+, granzyme+, and perforin+ lymphocytes also significantly increase after focused ultrasound treatment. 48 Focused ultrasound has also been used to disrupt the blood-brain barrier to deliver targeted NK-92 cells expressing a chimeric HER2 antigen receptor to HER2-expressing human breast tumor cells. 49
Current NK Cell Tracking Methods
Quantitation of the distribution and timing of NK cell trafficking is necessary to clinically assess the efficacy of their adoptive transfer. Using various imaging techniques, this evaluation can be done quickly and noninvasively in patients receiving adoptive NK cell therapy. There are currently three methods to track NK cells in vivo 50 : (1) optical imaging (OI) using fluorescence (FLI) or bioluminescent imaging (BLI), (2) radionuclide-based imaging using PET or SPECT, and (3) MRI.
Direct or indirect cell labeling can be used to track NK cells using OI. Cells can be directly labeled with exogenous fluorescent tracers for FLI or indirectly via transfection of a reporter gene that generates a protein product for FLI (e.g. green fluorescent protein, GFP) or BLI (e.g. luciferase). Fluorescent dyes can be used to permanently label NK cells and other lymphocytes through covalent and noncovalent binding to DNA, RNA, and protein sites; cell membrane insertion; and reactions in the cytosol. 51 Optical markers exhibit high sensitivity and high specificity, but provide limited resolution (2–3 mm), poor anatomical discrimination, and limited tissue penetration (1 mm fluorescence; 3 mm bioluminescence). OI is well suited for small animal models due to small tissue depths as compared to humans. There are also no readily available clinical instruments.
Using OI, NK-92scFv(MOC31)-zeta cells targeted to EpCAM antigen in human prostate cancer xenografts have been tracked in vivo in athymic rats using a near-infrared dye DiD (1,1‘-diotadecyl-3,3,3’,3’-tetramethylindodicarbocyanine). Tumor fluorescence increased significantly after injection of targeted NK-92 cells vs. nontargeted parental NK-92 cells (Fig. 1). 39 NK-92MI cells have also been labeled with anti-CD56 antibody conjugated with quantum dots (QD705) without affecting viability and IFG-γ production and cytolytic activity. QD705-labeled NK cells have been tracked up to 12 days post intratumoral injection in human malignant melanoma (MeWo) xenografts in mice. 52 Conjugation of fluorescence organic dye (Cy5.5) with magnetic nanoparticles (Fe3O4/SiO2) has also been used to visualize the migration of NK-92MI cells to the tumor site controlled by an external magnetic field. 53

Adapted from Tavri et al, 39 with permission. Optical (fluorescence) imaging scans of rats with EpCAM-positive DU145 prostate cancer xenografts (arrow) before and after injection of targeted (NK-92-scFv(MOC31)-zeta) (subject #4) and nontargeted NK-92 cells (subject #7) labeled with DiD. Scans show increases in tumor fluorescence signal at 1.5–24 hours post injection (p.i.) of NK cells. The subject that received nontargeted NK-92 cells shows nonspecific signal in the liver/lung region 1.5–24 hours p.i. The subject that received the targeted NK-92 cells showed the nonspecific liver/lung fluorescence only at 8 hours p.i.
PET and SPECT provide high sensitivity for detecting labeled NK cells (in the nanomolar concentration range), high specificity (target to background contrast), quantification of labeled cells, and has immediate clinical translation through use of FDA-approved labeling agents. However, limitations include poor spatial resolution (~1–5 mm), limited anatomic information, ionizing radiation, and, depending on the radionuclide, limited duration and number of scanning sessions. 50 Radionuclide tracer decay limits follow-up studies within hours ( 18 FDG 2–4 hours) to days (111In 4–7 days). Integration of computed tomography (CT) can improve the resolution of PET and SPECT through attenuation correction and allows registration of tracer signals with anatomical structures. 54
NK cells have been labeled with positron emitting radionuclides, including 18 FDG (half life = 109 min) and 11 C-methyl iodide (half life = 20 min) for PET,22,35,55 as well as with gamma emitting radioisotope 111In (half life = 2.8 days) for SPECT.56,57 PET tracers have been used to experimentally investigate the optimization of NK cell targeting to tumors. Melder et al, using 11 C-methyl iodide and PET, demonstrated that 4–30% of injected activated NK cells localized (1 hour post injection) to the tumor site of a murine FSaII fibrosarcoma model compared to 3–4% of nonactivated splenic lymphocytes. 22 Using an FDA-approved PET label, 18 FDG, Meier et al tracked genetically modified NK-92 cells, NK-92-scFv(FRP5)-zeta cells. These cells express a chimeric antigen receptor targeting tumor-associated Her2/Neu antigens, and were shown to accumulate more in the tumor site compared to parental NK-92 cells in mice (Fig. 2). 35 Additionally, SPECT agents have been used to explore optimal administration routes for adoptively transferred NK cells clinically. Matera and colleagues used 111In-oxide-labeled recombinant IL-2-activated NK cells, evaluated by SPECT, to show that intra-arterial injection vs. intra-venous results in increased accumulation of adoptively transferred NK cells in the liver tumor site (colon carcinoma metastasis). 56

Reprinted with permission from Meier et al. 35 (Left) Representative digital autoradiographs of tumors and organ tissues from mice with HER2/ neu-positive sarcoma that received 18 FDG-labeled targeted NK-92 (NK-92-scFv(FRP5)-zeta) cells (A) and nontargeted parental NK-92 cells (B). The signal intensity bar shows count density of photostimulated luminescence per unit area: PSL/mm2 (in log scale). (Right) Uptake of radioactivity in selected organs subtracted from the background noise (rectangular box on autoradiograph image).
For MRI, NK cells have been labeled with nanoparticles, such as superparamagnetic iron oxide (SPIO) and ultra-small superparamagnetic iron oxide (USPIO) nanoparticles.34,36,58,59 Depending on how long the adoptively transferred NK cells survives; NK cells are able to retain their iron oxide label up to 4 days.
58
Iron oxide-based agents produce strong hypointensity in
Meier et al demonstrated significant decreases in

Reproduced with permission from Meier et al.
36
Axial and coronal
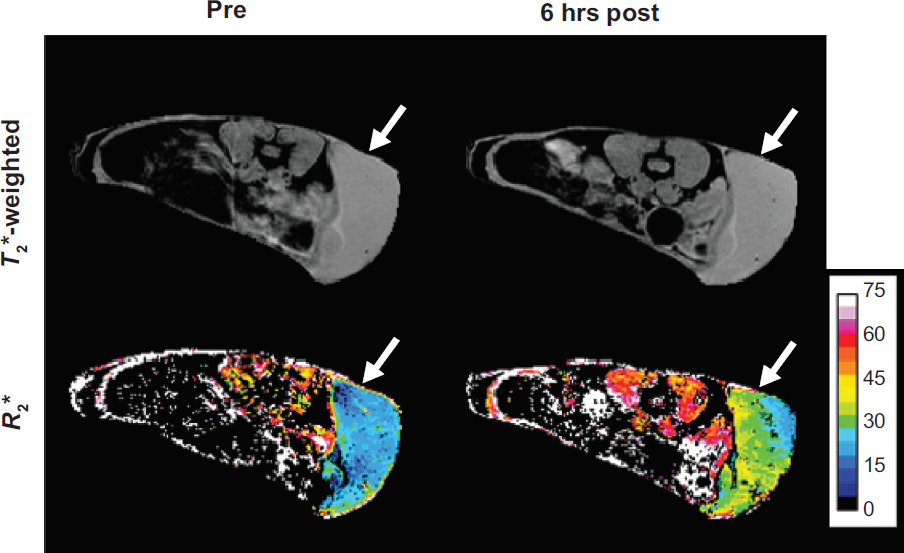
Axial
MRI is readily available clinically, has relatively high resolution (better than 0.1 mm), well-defined soft-tissue contrast, and has no ionizing radiation. Disadvantages of MRI compared to OI or radiotracer-based imaging are limited sensitivity for detecting small cell populations, relatively high cost, long scan times, and low specificity. Quantitation of cells in vivo is theoretically possible using relaxation rate maps, but this type of quantification is indirect as the MR signal detected is related to the perturbation in tissue proton magnetization rather than directly to cell concentration.
Conclusion
Autologous NK cell or NK cell lines are being used clinically. Multiplexing adoptive transfer with patient and tumor priming (via adjuvant and/or immunocytokine treatments, genetic modification of NK cells, and/or combinatorial therapies) are being explored to optimize targeting to the tumor and enhance cytotoxicity of NK cells. In vivo assessment of the spatiotemporal distribution and the concentration of transferred NK cells within tumor sites are necessary to evaluate the efficacy of NK cell-based therapies in patients. Existing imaging techniques with optimized probes can be used noninvasively and results can be visualized in real time. For future clinical applications, multimodal imaging (MRI and PET) offers the most desirable characteristics for measuring responses to NK cell adoptive therapy, to qualitatively and quantitatively track cells, and to measure cellular viability through visualization of PET and MRI signal colocalization.
Footnotes
Acknowledgments
The authors acknowledge the contributions of Sharon Lin for invaluable assistance with animal- and cell-based work. Drs. Andrew Raubitschek and David Colcher (City of Hope, Duarte CA) provided cogent comments as well as animals and cells for our NK cell studies.