
Abstract
Fungal exopolysaccharides (EPSs) have been recognized as high value biomacromolecules for the last two decades. These products, including pullulan, scleroglucan, and botryosphaeran, have several applications in industries, pharmaceuticals, medicine, foods etc. Although fungal EPSs are highly relevant, to date information concerning fungal biosynthesis is scarce and an extensive search for new fugal species that can produce novel EPSs is still needed. In most cases, the molecular weight variations and sugar compositions of fungal EPSs are dependent to culture medium composition and different physical conditions provided during fermentation. An inclusive and illustrative review on fungal EPS is presented here. The general outline of the present work includes fungal EPS production, their compositions and applications. An emphasis is also given to listing out different fungal strains that can produce EPSs.
Keywords
Introduction
Carbohydrates are naturally occurring and well distributed, and are the most important building blocks of the biosphere. These evolutionary and biologically important organic compounds are present on Earth in different forms. Traditionally, on the basis of the number of sugar units, carbohydrates are classified into three groups: monosaccharides, oligosaccharides, and polysaccharides. The natural macromolecules composed of several monosaccharide units (more than ten) are known as polysaccharides and are synthesized at different stages of life cycle of every living organisms for different purposes. The monosaccharide units of polysaccharides are joined to each other by an acetal linkage. These acetal linkages are formed by the reaction of a hemiacetal hydroxyl group of one unit with an alcohol group of another unit which liberates water to give a glycosidic bond. Polysaccharides not only have different sequences of monomeric units but also have different sequences of glycosidic linkages and different types of branching. They may be amorphous or even insoluble in water. All these factors together give polysaccharides a great diversity of structure, property, and functions. Polysaccharides used at the industrial level are almost all of plant and seaweed origin. 1 These long-chain and high molecular weight polymers, such as starch, alginate, Arabic gum, carrageenan, agar, and gaur gum, are widely employed in the food, pharmaceutical, and cosmetic industries.2–4
Microorganisms, however, are known for their ability to synthesize polysaccharides with different structural complexities.5,6 These polysaccharides either remain attached to the cell surface or are found in the extracellular medium. 7 Two hundred years ago, Henri Braconnot discovered chitin, the first carbohydrate polymer which is found in edible mushrooms.8,9 The extracellular polysaccharide production by microbes was first reported in 1861 as a “viscous fermentation” by Pasteur. The organism that produces this polysaccharide was a bacterium identified as Leuconostoc mesenteroides by Van Tieghem. 10 Among the three major classes of microbial polysaccharides, exopolysaccharides (EPSs) had several advantages over intracellular and cell wall polysaccharides including huge production in short time, easy isolation, and purification. EPSs of microbial origin might represent a valid alternative to plant and algal products considering that their properties are almost identical to those currently used gums. 1 In other cases, the microbial products have unusual molecular structures and peculiar conformations, thus conferring unique and potentially interesting properties with potential industrial uses.1,3,5,11 Together with the knowledge of biological properties, the structures and fermentation process of microbial EPSs produced are also very important for understanding their physiological activities and industrial applicability.
Among the microbial EPS producers, bacteria and fungi are most common. Bacterial EPSs have been studied extensively by researchers. 6 EPS production from fungi has been studied adequately over the last two decades. Different EPS production by fungi including Ganoderma lucidum, Agaricus blazi, Cordyceps sp., Lentinus edodes, and Grifola frondosa through submerged cultures had been reported, all of which have different and interesting biological activities. 12
In the present review, an attempt was taken to recapitulate all the existing literature concerning fungal EPS production, composition, and biological applications in a single frame to allow us to realize the advances and perspectives in the knowledge and applications of fungal EPSs.
Production of Exopolysaccharides from Fungi
EPS production from fungi mainly depends on the type of fungal strain used, physical conditions maintained during fermentation, and type of medium components applied for the production.
Fungal strains producing exopolysaccharides
At present, a considerable number of fungi including higher basidiomycetes, lower filamentous fungi, and yeasts from different ecological niches were known for their ability to synthesize EPSs in laboratory culture systems. However, many still remain uninvestigated or under explored. For better observations at a glance, list of reported fungal EPS producers are represented in Table 1.
List of some fungi, are reported to produce exopolysaccharides in laboratory culture techniques.

Parameters affecting exopolysaccharide production
EPSs are generally synthesized intracellularly and secreted to the surroundings. Very little information is available regarding the biosynthesis of EPSs from fungi.1,13 Only a few fungal EPS biosynthesis pathways have been studied, such as EPS biosynthesis by G. lucidum. 14 Most researchers have studied the optimization of culture medium and conditions for EPS production from different fungal strains. In this context, Figure 1 recapitulates the main factors upon which fungal EPS production is dependent. The majority of the published papers followed liquid submerged culture techniques for EPS production and some researchers used statistical methods including response surface methodology, orthogonal matrix method using Box-Behnken design, Plackett-Burman design, central composite design, or fractional factorial design for optimization of EPS production in liquid submerged fermentation.15–18 Sandford suggested that most of the EPS producing fungi are aerobic or facultative anaerobic, and oxygen limitation did not support EPS production. 19 Like Sandford, published articles have also showed that EPS production by fungi normally reached its optimum level in shaking conditions when oxygen is available in the medium. 19 Ruperez and Leal evaluated the EPS production by Aspergillus parasiticus in agitated and static submerged fermentation. 20 In agitated culture, this organism produced more EPSs (0.41 gm/L) than static culture (0.18 gm/L). Agitated culture technique for EPS production by this Aspergillus species was also favorable considering the cultivation time. In agitated culture, maximum EPS production was found after 5 days of fermentation whereas in static culture 6 days of cultivation are needed.
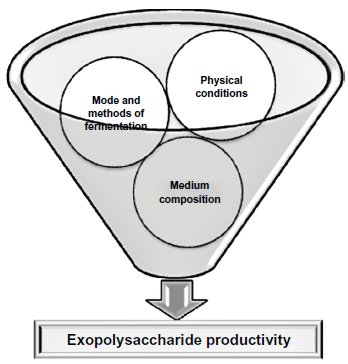
A schematic illustration: main factors on which fungal exopolysaccharide production depends (Physical conditions: temperature, pH, oxygen level, incubation time etc.; Medium composition: carbon source, nitrogen source, different salts, special additives like vegetative oils, vitamin etc.; Mode and methods of fermentation: agitated culture, static culture, submerged culture using general optimization process or statistical methods like response surface methodology, orthogonal matrix method using Box-Behnken design, Plackett-Burman design etc.).
Roukas and Liakopoulou-Kyriakides examined the production of pullulan by Aureobasidium pullulans P56 and reported that EPS production was higher (23 gm/L) at an aeration rate of 1 vvm, in comparison to cultures with low aeration of 0.5 vvm (14 gm/L) and culture without oxygen supply (12 gm/L). 21 Kim et al reported with high aeration rate (3.5 vvm), the increase in the dissolved oxygen is associated with higher EPS production by Paecilomyces sinclairii. 22 Xu and Yun examined the influence of aeration on EPS production by Paecilomyces tenuipes C240 in a stirred-tank fermenter and found that at a high aeration rate (3.5 vvm), maximum EPS (2.36 gm/L) was produced. 23 Among the three different extracellular proteoglycans produced by this organism, two of them showed variation in their composition when produced in various aeration conditions. Gibbs and Seviour reported that pullulan production from A. pullulans ATTC 9348 was optimally supported up to a certain level of dissolved oxygen in bioreactor, over which EPS production decreased. 24 These findings suggest that EPS production by fungi is oxygen dependent but there is a specific limit beyond which production might be reduced.
The pH of culture medium is another reflective factor that persuades the fungal EPS production. 25 Generally, fungi favored low pH for EPS production with a range between pH 3.0 to 6.5.15,18,26–32 A few fungi preferred neutral or alkaline pH for maximum EPS production.33–35 In 2004, Shu and Lung examined the effects of pH on EPS production by Antrodia camphorate and reported that variation in medium pH induces A. camphorate to produce EPS with different molecular weight (Mw). 36 They noticed that relatively high Mw EPSs in low amount was produced at lower medium pH while low Mw EPS with high yield was recorded at higher medium pH. In 2012, Abdel-Aziz et al reported that acid pH shock induced the EPS production by Mucor rouxii. 37 Authors reported that M. rouxii showed normal growth at pH 5.0 to 7.5 but EPS production increased gradually with increase in pH values up to a pH of 10.0. They also noticed that when the initial medium pH adjusted at pH 3.5, it showed maximum flocculating activity. 37
Most of the fungal strains produced maximum EPSs within a temperature range 22 °C to 30 °C (Table 2). Only a few reports suggested that 20 °C was most effective for EPS production by fungi.26,34
Optimum culture conditions for maximum exopolysaccharide production by different fungal strains.
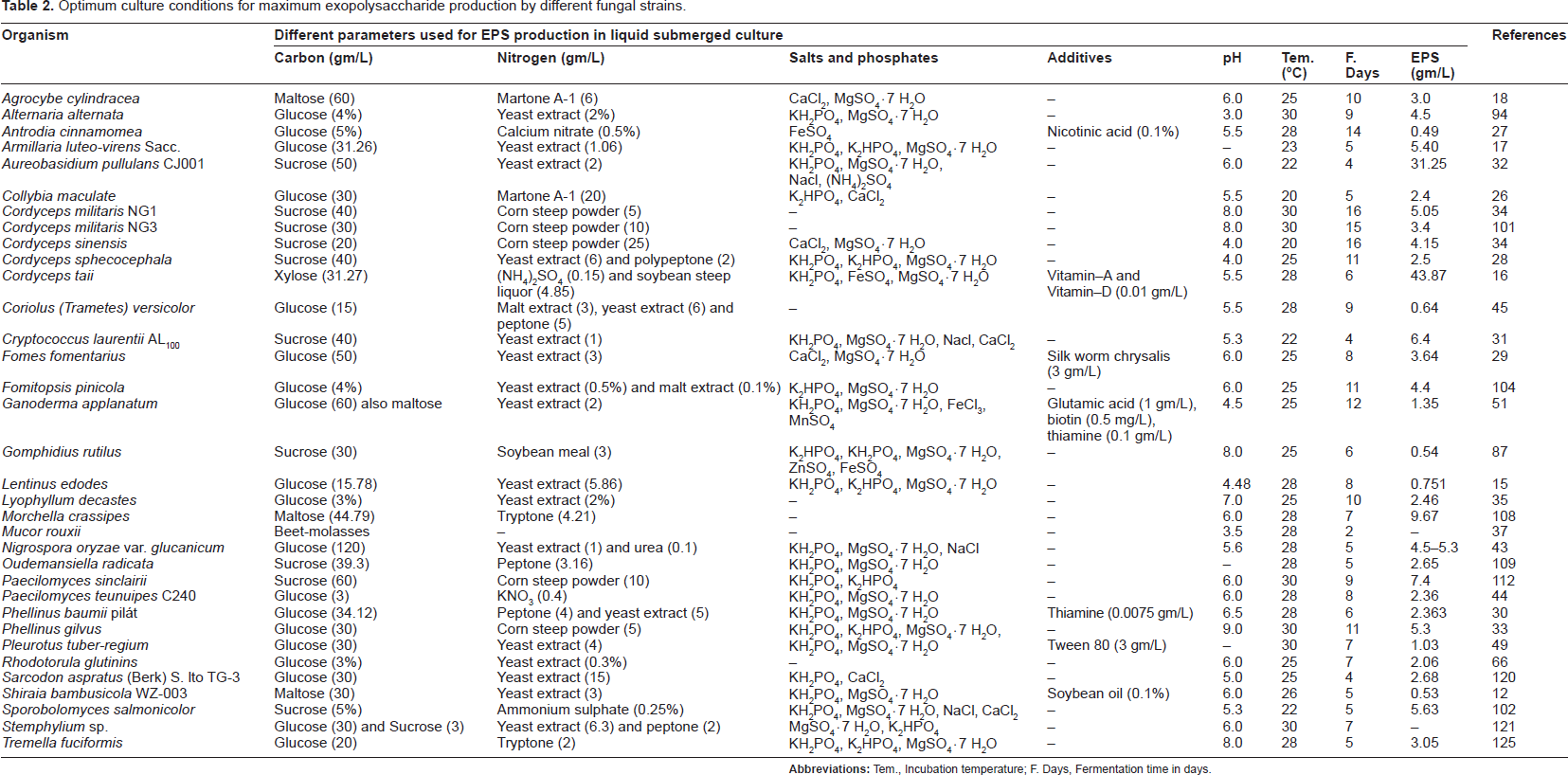
EPS production by fungi at different time intervals was studied by many researchers. Their findings suggested that EPS production could be maximized either at its late exponential stage or its early stationary stages of growth. Unlike bacterial EPS production, fungi needed long incubation time for maximum EPS production. Generally 4 to 15 days of fermentation needed by different fungal strains for optimum EPS production (Table 2). Shu and Lung studied the effect of incubation time on EPS produced by A. camphorate. 36 They found that high molecular weight EPS reduced with increase in fermentation time and, that after completion of fermentation, only low molecular weight EPSs were present in the fermented broth. Similar results were also reported by Lee et al when they were studied the pullulan production from A. pullulans. 38
Although in most cases composition of EPS is independent to the type of carbohydrate used for the production of that EPS, the production intensity is very much dependant on the carbon source used and its concentration. In general, glucose, sucrose, maltose, lactose, fructose, galactose, xylose, cellobiose, sorbitol, xylitol, mannitol, and different types of agricultural byproducts are used as carbon source in the culture medium. In most of the cases glucose, sucrose, and maltose have been selected as the most influential carbon sources for the production of fungal EPSs (Table 2). These observations indicate that there may be some effects of catabolic repression of different sugars in various EPS synthesis, that different fungal strains have different sugar uptake fascinations, or that these sugars may be easily metabolized by fungi. Actual reasons, however, are not clearly understood as in most cases fungal growth and EPS production are not directly proportional. Elisashvili et al studied eight basidiomycetes for EPS production. 39 They reported that G. lucidum, Inonotus levis, and Phellinus robustus produced maximum EPSs in media containing glucose as a carbon source. Cerrena maxima, Phellinus igniarius, Trametes versicolor, on the other hand, favored maltose while Agaricus nevoi favored mannitol as the most supportive carbon source for EPS production. Bae et al evaluated the effect of carbon sources on EPS production by Paecilomyces japonica and reported that maltose is the preferred carbon source and results in higher EPS production (30 gm/L) than sucrose (25 gm/L). 40 In an another report, Elisashvili et al reported an interesting observation that L. edodes and Pleurotus spp. strains produced maximum EPSs in culture media containing sodium gluconate as carbon source but the reasons were not clearly understood. 41 The concentration of selected carbon source in the culture media is another critical factor for EPS production. In most of the findings, between 30 and 60 gm/L carbon was suggested to best support EPS production from fungi (Table 2), although a few exceptions were also reported by Fariña et al, 42 Sudhakaran and Shewale, 43 Xu et al, 44 Tavares et al. 45 Fariña et al studied Sclerotium rolfsii ATCC201126, which produced approximately four times more scleroglucan when grown in 150 gm/L sucrose than 20 gm/L from the same carbon source. 42 Sometimes combined carbon sources induced EPS production by fungi, demonstrated in the report of Zhang et al where they observed that 2 gm/L glucose with 30 gm/L starch influenced Sorangium cellulosum for maximum EPS (17.5 gm/L) production. 46
Nitrogen supplementation is another variable that is reported to induce EPS production. Both inorganic and organic nitrogen sources were tested by several researchers to find the appropriate one. Among the organic sources peptone, yeast extract poly peptone, Martone A-1, Soybean meal, and corn steep powder were tested mostly. From numerous research findings, it was noticed that yeast extract and corn steep powder are good nitrogen supplements that induce EPS production from different fungal strains (Table 2). Among the various inorganic sources ammonium chloride, ammonium sulfate, sodium nitrate, potassium nitrate, urea, and diammonium oxalate mono-hydrate are commonly studied by researchers. Many observations suggested that in the presence of inorganic nitrogen sources, fungi produce less EPSs in comparison to organic nitrogen supplements. Among the inorganic nitrogen sources, ammonium salts are frequently more efficient than other inorganic salts. 5 In very few studies, other inorganic salts have been found to best provide nitrogen sources for EPS production from fungi. Sodium nitrate had been found most suitable for the production of epiglucan by Epicoccum nigrum and for the production of scleroglucan by S. rolfsii ATCC201126.42,47 Urea was found most effective for EPS production from Nigrospora oryzae var. glucanicum 43 Sutherland reported that EPS production generally occurred in nitrogen limiting conditions. 1 Researchers examined and reported that for different fungal strains, different concentrations of selected nitrogen favored maximum EPS production (Table 2). Except for a few reports, scientists found that in comparison to carbon sources very little nitrogen is required by fungi for EPS production and concentrations between 1–10 gm/L is are sufficient (Table 2). In exceptional cases, where more than 10 gm/L nitrogen sources were reported as best EPS production supportive, an interesting observation was recorded by almost all researchers: the concentration of carbon sources is always ≥ 30 gm/L which indicates lower consumption of carbon comparing others.
Many findings indicate that a phosphate source is an important addition needed by fungi for EPS production. Potassium dihydrogen phosphate (KH2PO4) and dipotassium monohydrogen phosphate (K2HPO4) have been reported as the most efficient phosphate supplements (Table 2). Among the different additional ionic salts, many describe magnesium sulphate (MgSO4 · 7H2O) as most suitable for EPS production by different fugal strains while in some cases calcium chloride (CaCl2), sodium chloride (NaCl) are also needed by the fungi for optimal production (Table 2). Tang et al practiced the one-variable-at-a-time approach and response surface methodology to evaluate the effect of metal ions on EPS (Tuber polysaccharide) production by Tuber sinense 48 From their research they concluded that 30 mM Mg2+ and 5 mM K+ maximized the EPS production (5.86 gm/L) in submerged culture, which was 130.7% higher compared to EPS produced in basal medium without metal ions.
Effects of some other additives including vegetable oils, fatty acids, surfactants, and vitamins were also studied and reported.12, 16,49,50 These reports showed that sometimes these additives resulted in maximum EPS production. Lee et al used glutamic acid, biotin, and thiamine in culture medium for maximum EPS production. 51 Yang and He reported that addition of 0.1% soybean oil in culture medium influenced EPS production by Shiraia bambusicola WZ-003. 12 Yang et al evaluated the effect of additional fatty acids on mycelia growth and EPS production by suspended and immobilized G. lucidum. 50 They had reported that in suspension culture, palmetic acid and oleic acid at less than 0.25 gm/L showed incensement in EPS production whereas linoleic acid had a strong reductive effect on EPS production by this organism. In immobilized cultures only palmitic acid showed influential effect on EPS production. Addition of 3 gm/L Tween 80 induced maximum EPS production by Pleurotus tuber-regium. 49 Conversely, Hsieh et al reported that surfactants like Tween 80 or Span 80, although induce the cell growth of G. frondosa, significantly reduce the EPS production by the organism. 52 The authors also evaluated the effect of plant oils on EPS production and reported that addition of 0.5% olive oil at the stationary phage of fungal growth induced maximum EPS (2.248 gm/L) production while same oil addition in early growth phage induced mycelia growth and reduced EPS production. Xiao et al used vitamin A and D in the culture medium as growth-stimulating factor for maximum EPS production by Oudemansiella radicata. 16 Lin and Chen reported that 0.1% (v/v) thiamine induced EPS production in complex medium by Antrodia cinnamomea. 53 Use of nicotinic acid (0.1% v/v) in the EPS production medium was reported by Lin and Sung when they evaluated the optimization process for EPS production by A. cinnamomea. 27
Compositions of EPS Produced by Different Fungi
The monomeric composition and structure of fungal EPSs were usually evaluated by different experimental analysis of intact EPSs, hydrolyzed or partially hydrolyzed EPSs, or their derivatives. In general, these studies were analyzed through paper chromatography, HPLC, gas-liquid chromatography (GLC), gas-liquid chromatography-mass spectrometry (GLC-MS), and 1D and 2D NMR spectroscopy. The composition of fungal EPSs vary from pure sugars to sugars combined with a second unit such as protein, phosphate, sulfate, or amine. Different types of sugar unites were found in fungal EPSs such as glucose, mannose, galactose, xylose, fucose, and rhamnose. It was also noticed that EPSs composed of the same monosaccharide units that were synthesized by different fungi had different molecular weight. This is caused by differing chain length or branching patterns. To review the composition of EPSs and their corresponding molecular weight, see Table 3.
Compositions and molecular weight (Mw) distributions of some fungal exopolysaccharide.
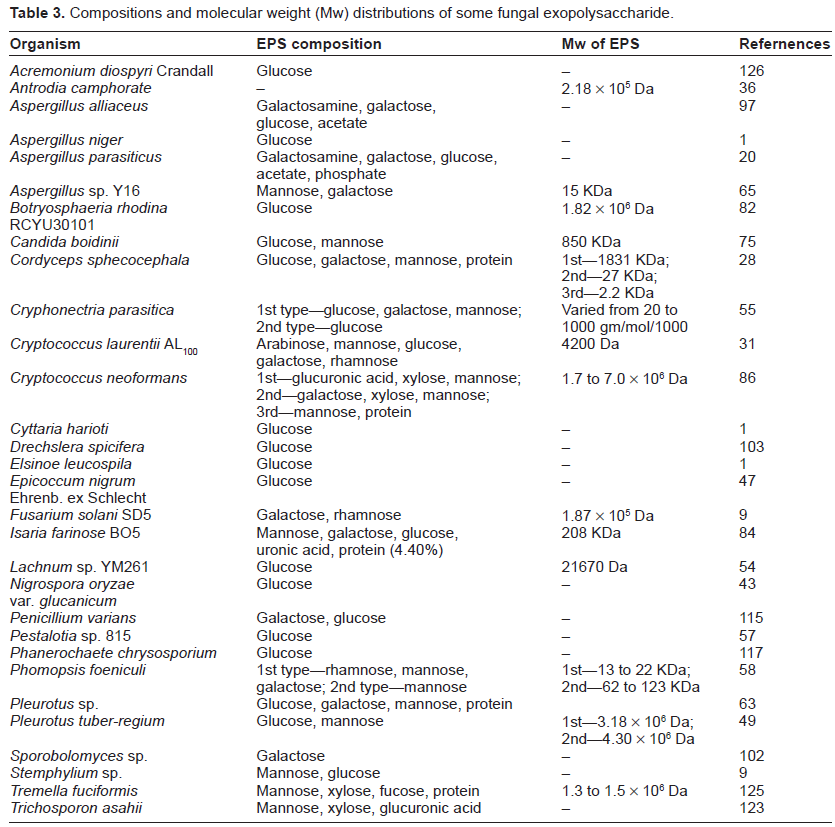
The linkage patterns and form of the monosaccharide units of fungal EPSs are also very diverse. Lachnum sp. YM261 produces an extracellular glucan with β-(1, 3)-D-pyran glycosidic bonding. 54 Ruperez and Leal reported an EPS of A. parasiticus having monosaccharides with α-configuration and also reported that it contained predominantly 1, 6 linkages. 20 Forabosco et al reported a novel type of pullulan production by Cryphonectria parasitica. 55 Unlike other pullulans which contain α-(1, 6) maltotriose, this one contains high levels of α-(1, 6) maltotetrose subunits. Xu et al reported that in two types of bioreactor EPSs having different molecular weight (total five) were isolated. 56 From a stirred-tank reactor, three types of EPSs were isolated among which two had glucose as the major monosaccharide component while the other types were composed majorly of mannose. In an airlift reactor, this organism produced two different EPSs. One showed glucose as its main building block, while the other showed arabinose. Pestolotan, an EPS produced by Pestalotia sp. 815, has three β-(1, 6) branched glucose residues for every five β-(1, 3) residues. 57 Corsaro et al reported on two types of EPS production of Phomopsis foeniculi. 58 One EPS produced by this organism is a galactan with structure of (-6)-β-D-Galacto furanose (1, 5)-β-D-Galacto furanose (1, 5)-β-D-Galacto furanose (1-)n. The other is a mannan with a backbone of α-(1, 6)-linked mannopyranose units. Leung et al reported an EPS having β-D-glucan backbone produced by Cordyceps sinensis Cs-HK1. 59 Wang et al reported another type of EPS production from the same Cs-HK1 strain, where researchers found that the isolated EPS has a chemical composition of α-D glucopyranose, α-D-pyrano glucuronic acid in molar ratio of 8:1 and trace amount of mannose. 60 Ichikawa et al reported an extracellular glucuronoxylomannan production from Trichosporon asahii. 61 This EPS has a chemical backbone of α-(1, 3)-D-mannan like hexasaccharide substituted with one β-(1, 2)-glucopyranosyluronic acid and six β-D-xylopyranosyl units. Bernabe et al reported an EPS production from Lecanicillium muscarium CBS 413.70C. 62 The structural composition study suggests the presence of α-(1, 4)-glucopyranosyl units and α-(1, 6)-glucopyranosyl units. Gutierrez et al reported that six species of Pleurotus genus produced EPSs with the main building block of β-(1, 3)-D-glucose and that 25% of total units are C-6 branched. 63 Kim et al reported on the production of an EPS by G. resinaceum where under different culture conditions in 5-l stirred tank bioreactor, composition of EPSs were varied and that the main monomeric units of these EPSs were fucose, xylose, mannose, glucose, galactose as well as some protein moities. 64 EPS of P. tuber-regium has a main building block of 1,6 linked mannopyranosyl residues almost all of which are branched at O-2 with a side chain containing two 2,6 linked mannopyranosyl residues and a terminal glucopyranosyl residue. 49 The back bone of the EPS produced by endophytic Aspergillus sp. Y16 contains mainly 1, 2 linked α-D-mannopyranose units, substituted at C-6 by 1, 6 linked α-D-mannopyranose, 1 linked β-D-galactofuranose and 1 linked β-D-mannopyranose units. 65 Ibrahim et al evaluated the compositions of EPS produced by Rhodotorula glutinins and reported that its EPS is made up of mannose, glucose and arabinose in a molar ratio of 3.2:1.0:0.8. 66 S. cellulosum produced an EPS containing 38.3% proteins and 58.5% carbohydrates, of which glucose, mannose and glucuronic acid were present at 51.3%, 39.2% and 10.5%, respectively 46 Mahapatra and Banerjee reported a rhamnogalactan production from endophytic fungus Fusarium solani SD5. Structural elucidation of this EPS indicated the presence of terminal α-L-rhamnopyranosyl, (1, 2)-α-L-rhamnopyranosyl, (1, 4)-β-D-galactopyranosyl, (1, 4, 6)-β-D-galactopyranosyl moieties in a molar ratio of 1:1:3:1. 9
Applications of Different Fungal EPSs
Microbial EPSs, including fungal EPSs, have gained importance from the last few decades as several studies showed different applications that not only indicate the alternative source of marketed plant or seaweed polysaccharides but also have some new and interesting bioapplicability. Furthermore, upstream and downstream processing of these EPSs is easier and one can produce a much larger amount in a shorter time when compared to plant or algal polysaccharide production. Fungal EPSs have several applications in the food and pharmaceutical industries among others. An overview on different fields where fungal EPSs can be applied is presented in Figure 2. Among the different fungal EPSs pullulan, Scleroglucan, and Botryosphaeran are well known for their applications in different fields. Here, some reported applications of different fungal EPSs are listed out for understanding the wide functional scope of these biomacromolecules.
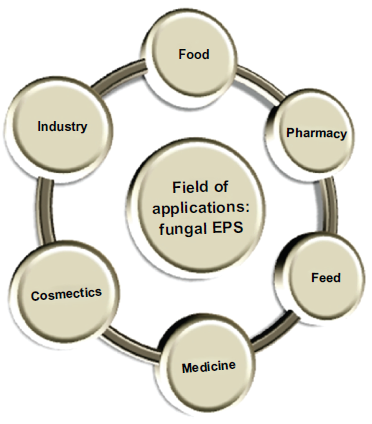
An overview: field of applications of fungal exopolysaccharides.
Pullulan, an EPS of A. pullulans, can be used as a thickener, a viscosity stabilizer in the food industry, and also for the preparation of nontoxic, biodegradable, edible plastic materials. 67
Fungal EPS Scleroglucan is used industrially for the Enhanced Oil Recovery 68 Scleroglucan is sometimes called schizophyllan under the industrial name BIOVIS (produced by Degussa society) with bacterial polysaccharide, xanthan gum applied is used for preparation of a drilling fluids (water based muds) with low mud toxicity. 69 Farwick et al reported the high water binding capacity of scleroglucan on epithelial cells. 70 Other industrial applications of this EPS were in preparation of adhesives, water colors, printing inks, and animal feed composition. 71 It is also used in the manufacturing of cosmetics and in various skin care products, creams and protective lotions.72,73
EPS of Byssochlamy nivea showed Kaolin-flocculating behavior. Gomoiu and Catley suggested that this EPS can be used for sedimentation of fibers in downstream removal of paper fibers from white water effluent in paper industries and thus possesses pollution reducing efficacy in paper industries as well as rivers where industrial effluents were discarded. 74
EPSs from five yeast strains and one yeast-like fungus showed drag-reducing activity. 75
Exopolymer (glycoprotein) produced by G. lucidum can increase the swimming endurance capacity of mice by about 10 minutes and reduced the muscle and liver glycogen exhaustion by 18.5% and 67.2% respectively. 76
Fungal β-glucans (both extracellular and intracellular) are effective in promoting health, protection from mutations and treatment of diseases like cancer, microbial infections, hypercholesterolaemia and diabetes.77,78
Pestan, a fungal EPS produced by Pestalotiopsis sp. KCTC 8637, has applications in wastewater treatment as a biosorbent of lead and zinc. Each gram of pestan absorbed 120 mg lead and 60 mg zinc. 79 EPS of Aspergillus fumigatus also showed sorption efficiency of two heavy metal ions, copper and lead. 80
Botryosphaeran, a EPS of Botryosphaeria rhodina, was shown to significantly decrease the clastogenic effect of cyclophosphamide-induced micronucleus formation in polychromatic erythrocytes of bone marrow and reticulocytes in peripheral blood in mice. 81 Botryosphaeran is also toxicologically accepted and a potent immunomodulator. 82
EPSs produced by Akanthomyces pistillariiformis BCC2694, Cordyceps dipterigena BCC2073, P. tenuipes BCC2656, and Phytocordyceps sp. BCC2744 was shown to be biocompatible and have potentiality as a wound dressing material through testing induction in interleukine-8 production in normal human dermal fibroblasts cells. 83
EPS of Phellinus baumii Pilát showed direct immune-stimulating activity on splenocyte proliferative response and acid phosphatase activity in peritoneal macrophages of mice. 30 Wang et al reported the immunomodulatory activities of EPS produced by C. sinensis Cs-HK1. The activities were evaluated in Raw 264.7 macrophage cell cultures. 60
Water soluble extracellular polysaccharides of Isaria farinose BO5 showed antitumor and antioxidant activity in Kunming mice. 82 EPS of Hypsizigus marmoreus and Fomes formentarius showed antitumor activity examined on human gastric cancer cells SGC-7901.29,85
Extracellular glucuronoxylomannan of Cryptococcus neoformans inhibited the entry of HIV virus in TZM-bl cell lines. 86
An extracellular glucan of Lachnum sp. YM261 showed strong anti-ageing activity tested in D-gal model mice. 54
C. sinensis Cs-HK1 produced an EPS which has moderate antioxidant activities. 57 EPSs of Ganoderma resinaceum, Gomphidius rutilus and endophytic Aspergillus sp. Y16 showed in vitro anti oxidant activity.64,65,87
Endophytic Fusarium oxysporium DzF17 has been reported as an EPS producer. Its EPS showed elicitor activities on growth and diosgenin production in cell suspension culture of Dioscorea zinbiberensis. 88
Endophytic F. solani SD5 was reported to produce an extracellular rhamnogalactan that showed anti-inflammatory and anti-allergic activity in vitro. EPS (1000 µg/mL) protects 55% of erythrocytes from hypotonic solution induced membrane lysis. Compound 48/80 induced mast cell degranulation was also protected by 56% with 100 µg/mL EPS. 9
The EPS of G. frondosa HB0071 showed inhibitory effect on matrix metallo proteinase-1 expression in UV-irradiated human dermal fibroblasts and thus may contribute to inhibitory action in photo-aging skin by reducing matrix metallo proteinase-1 related matrix degradation system. 89
P. baumii and Stropharia rugosoannulata produce EPS which show hypoglycemic activity in streptozotocin induced diabetic rats. 90
Preliminary examinations with EPS produced by R. glutinins are found to have anti-oxidant, antiviral, and antitumor activities. 66
Conclusion
In 2001, Hurtley et al despondently stated that the chemistry and biology of carbohydrate research is like a “Cinderella field”, though promising but to date does not get as much attention as genomes and proteins. 91 This realization is tinged with realism, although fungi have been extensively applied in industry since the 1940s for diverse bioactive metabolite production. In the case of EPS production, however, the efforts were not satisfactory to quench the thirst for knowledge in this field up until recently. Considerable increases in investigation of fungal EPS production or their physio-chemical characterizations by researchers were noticed only over the last two decades. Nevertheless, the findings are impressive from both the scientific angle and applicability. The hope is that this trend will continue to increase and will enrich both scientific knowledge base and provide better assistance for life in the future.
Funding
Author(s) disclose no funding sources.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication author(s) have provided to the publisher signed confirmation of compliance with legal and ethical obligations including but not limited to the following: authorship and contributorship, conflicts of interest, privacy and confidentiality and (where applicable) protection of human and animal research subjects. The authors have read and confirmed their agreement with the ICMJE authorship and conflict of interest criteria. The authors have also confirmed that this article is unique and not under consideration or published in any other publication, and that they have permission from rights holders to reproduce any copyrighted material. Any disclosures are made in this section. The external blind peer reviewers report no conflicts of interest. Provenance: the authors were invited to submit this paper.