
Abstract
The use of antibiotics is unavoidable in trying to treat acute infections and in the prevention and control of chronic infections. Over the years, an ever increasing number of infections has escalated the use of antibiotics, which has necessitated action against an emerging bacterial resistance. There seems to be a continuous acquisition of new resistance mechanisms among bacteria that switch niches between human, animals, and the environment. An antibiotic resistant strain emerges when it acquires the DNA that confers the added capacity needed to survive in an unusual niche. Once acquired, a new resistance mechanism evolves according to the dynamics of the microenvironment; there is then a high probability that it is transferred to other species or to an avirulent strain of the same species. A well understood model for studying emerging antibiotic resistance and its impact is Pseudomonas aeruginosa, an opportunistic pathogen which is able to cause acute and chronic infections in nosocomial settings. This bacterium has a huge genetic repertoire consisting of genes that encode both innate and acquired antibiotic resistance traits. Besides acute infections, chronic colonization of P. aeruginosa in the lungs of cystic fibrosis (CF) patients plays a significant role in morbidity and mortality. Antibiotics used in the treatment of such infections has increased the longevity of patients over the last several decades. However, emerging multidrug resistant strains and the eventual increase in the dosage of antibiotic(s) is of major concern. Though there are various infections that are treated by single/combined antibiotics, the particular case of P. aeruginosa infection in CF patients serves as a reference for understanding the impact of overuse of antibiotics and emerging antibiotic resistant strains. This mini review presents the need for judicious use of antibiotics to treat various types of infections, protecting patients and the environment, as well as achieving a better treatment outcome.
Keywords
Introduction
Evolutionary events are, in general, dependent on spatial and temporal factors that initiate or drive changes in a biological entity. Bacteria are well studied for their response to extracellular factors in the human body forcing them to choose between acute and chronic mode of infections. Highly virulent strains of bacteria are able to cause acute infections with marked symptoms. In immune compromised individuals, even less virulent bacteria are capable of causing infections. These opportunistic pathogens manifest chronic infections through prolonged colonization of the human body. Given interventions from the environment, new pathogens emerge and less pathogenic bacteria become infectious by acquiring virulent traits through horizontal gene transfer. 1 Infectious diseases are responsible for more than a quarter of deaths worldwide. In other words, treatments that require the use of antibiotics are on the rise, as is the emerging resistance to antimicrobials among bacteria. In industrialized nations such as the US, more than half of antibiotics used in livestock and animal agriculture are for therapeutic purposes, and about 60%–80% of total antibiotics are used for prophylactic or nontherapeutic purposes to promote growth and to improve feed efficiency (Fig. 1).2,3 In this scenario, gut microbiota and other microorganisms continually exposed to sub-inhibitory concentrations of antibiotics pave the way for the development of resistance following adaptation to unusual niches. Bacteria that can thrive in diverse niches have a serious impact on therapeutic choices available for treating infections. This review highlights the combined effects of human genetic disease, opportunistic bacterial infections, and drug treatments that lead to emerging antimicrobial resistance.
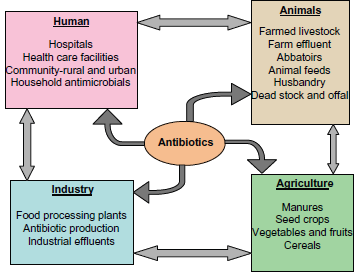
Usage and distribution of antibiotics.
Cystic Fibrosis and P. aeruginosa
Cystic fibrosis (CF) is one of the most common autosomal recessive genetic disorders among Caucasians. It is caused by mutation in the cAMP-regulated chloride channel, the CF transmembrane conductance regulator gene (CFTR), which leads to generalized exocrinopathy. As a consequence, dysregulated airway surface liquid in the lungs, accumulation of mucus, and thickened secretions contribute to the defective mucociliary clearance of microbes. In addition, continuous buildup of mucus creates a gradient of oxygen concentration in the lung environment and contributes significantly to the pathogenicity of lungs affected by CF. Mucus plugs create a suitable environment for bacteria to colonize and persist in the unusual niche where mucus also serves as a nutrient. 4 Poor prognosis of CF and the resulting mortality are attributed to microbial infections and persistent inflammatory response against pathogens in the lung. 5 Though the CF lung environment harbors different species of bacteria, only a few are found to be predominant, including Pseudomonas aeruginosa, Staphylococcus aureus, Burkholderia cepacia complex, Haemophilus influenzae, Stenotrophomonas maltophilia, and Achoromobacter xylosoxidans.6–8 Among bacteria responsible for deteriorating lung function, the role of P. aeruginosa has been explored widely in the context of global population structure, emergence of antibiotic resistance, and chronic colonization of CF lung through the formation of biofilms.5,9,10 Infection rates increase steadily with age, reaching up to 80% of adults 25 years or older colonized with P. aeruginosa.14,15
A huge genetic repertoire and mosaic genome structure makes P. aeruginosa a versatile opportunistic pathogen in nosocomial settings, particularly conditions involving burns and wounds, meningitis, endocarditis, and microbial keratitis. In the CF lung, P. aeruginosa displays a common phenotype irrespective of the genetic content, including mucoidy, lipopolysaccharide modifications, lack of flagella and pili, and upregulated antibiotic efflux after prolonged colonization.11–13 Controlled expression of genes in the genome backbone, DNA insertions, and in the extra-chromosomal elements equip P. aeruginosa to colonize CF lung and gradually outcompete other bacteria in the microenvironment. Besides intrinsic resistance mechanisms P. aeruginosa develops resistance to antibiotics during the course of accumulation of mutations in genes that encode antibiotic targets. Resistance is also conferred in this bacterium through the acquired gene cassettes from elsewhere.
Resistance to Antibiotics
Antibiotic resistance mechanisms in bacteria include enzymatic inactivation, target modification, overexpression or by-pass, sequestration, intracellular localization, increased efflux and decreased influx, and physical blockade through biofilm formation (Fig. 2). In addition, resistance to antibiotics arises because of spontaneous or induced mutations in genes associated with the resistance traits. In a community setting such as the lung of CF patients, microbes evolve by various means. To date investigations have revealed de novo incorporation of new genes and small sequence changes that lead to changes in the function of existing genes. Gene duplication and divergence of one or both of the copies, as well as shuffling and swapping of large domains of DNA, have also been observed.
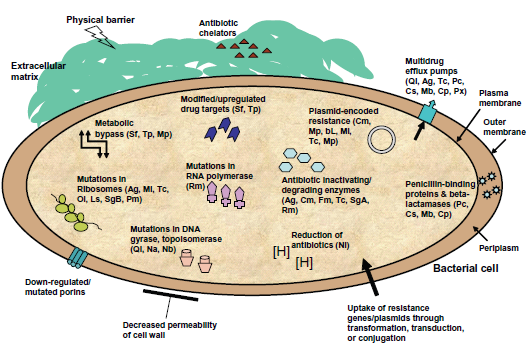
Innate and acquired antibiotic-resistance mechanisms in bacteria.
Bacteria in the form of biofilms show increased resistance to several antibiotics when compared to planktonic or free living counterparts. The minimal inhibitory concentration of tobramycin for P. aeruginosa microcolonies/biofilm, grown in artificial sputum medium, increased at least 50 times compared to planktonic forms. 4 Biofilm bacteria sometimes display up to a thousand-fold increase in antibiotic resistance, when compared to planktonic bacteria. 16 This characteristic is because of reduced diffusion or entrapment of antibiotics by exopolysaccharide matrix, slow growth rates, development of dormant persister cells, and the expression of antibiotic efflux or modifying proteins.10,17–19 The exopolysaccharide alginate produced by P. aeruginosa has been shown to retard the movement of cationic antimicrobial peptides, quaternary ammonium compounds, and aminoglycoside antibiotics through the matrix, thereby reducing the effective concentration of antibiotics. 20 In addition to the mentioned factors, sub-MIC levels of antibiotics promote adaptation of P. aeruginosa to the CF lung milieu through the accumulation of mutations in antibiotic target genes, eventually aiding survival in the hostile environment.21–23 Subinhibitory concentrations of aminoglycosides enhanced biofilm formation by P. aeruginosa through aminoglycoside response regulator (arr), by modulating intracellular levels of a secondary messenger molecule, cyclic-di-GMP. 40 Periplasmic glucans produced by clinical isolates of P. aerguinosa bind aminoglycosides, thereby reducing its effective concentration.36,37 Bacteriophages are reservoirs of antimicrobial resistance genes and play an important role in restructuring bacterial genomes.41,42
A study on CF sputum microbiology in the US from 1995 to 2008 involving different CF centers showed P. aeruginosa resistance to tobramycin and amikacin became increasingly prevalent and antibiotic resistance increased among other bacteria in the CF lung. 27 The emergence of antibiotic resistant isolates is mostly connected to mutations in the DNA mismatch repair system. 28 In addition, bacteria in the CF lung that are often exposed to the oxygen radicals generated by polymorphonuclear leukocytes (as a result of chronic inflammation) often damage DNA. Hyper-mutators arise due to mutations in mismatch repair genes mutS, mutL, and in DNA oxidative repair system genes mutT and mutY. They have developed resistance to antibiotics through increased production of beta-lactamases and MexCD-OprJ efflux pump.22,23,29 Hypoxic conditions altered stoichiometry of multidrug efflux pump linker protein expression towards a dominant MexEF-OprN, which conferred resistance to antibiotics.33,34 Similarly, upregulation of the MexXY efflux system is necessary for adaptation of P. aeruginosa to the CF lung environment. 19
Spontaneous or induced mutations–-single or multiple–-in antibiotic target genes confer resistance. Mutations in the domain V of 23S rRNA conferred azithromycin resistance in P. aeruginosa. 35 Extensively drug resistant, high risk clones possess mutations in multiple genes, including oprD, gyrA, parC, and mexZ, and overproduction of AmpC and AadB, all of which are involved in antibiotic resistance. 39 Mutation in cbrA (a sensor kinase of the cbrAB two-component regulatory system) enhanced biofilm formation and conferred resistance to polymyxin B, ciprofloxacin, and tobramycin. 38 These genetic variations in a P. aeruginosa population offer the advantage of an insurance effect. The ‘insurance hypothesis’ states that diversity within a population provides protection for the community as a whole against a wide range of adverse or changing conditions. 30 A shift in the bacterial metabolism to suit the nutrient status of a microenvironment also leads to the development of antibiotic resistance. In a study, the prototype strain of worldwide spread P. aeruginosa clone C displayed slow growth rate and a unique proteome pattern in 2D-gels. Identification of protein spots revealed down-regulation of proteins involved in the metabolism of amino acids and fatty acids, as well as of proteins involved in antibiotic influx. 31
P. aeruginosa modulates production of virulence factors in a cell-density dependent fashion through quorum-sensing (QS) circuitry. QS consists of a las and rhl N-acyl homoserine lactone system that produces 3-oxo-C12- and C4-homoserine lactones, respectively. 24 The third QS 2-alkyl-4-quinolone system produces 2-heptyl-4-quinolone and Pseudomonas quinolone signal. 25 These QS systems not only engage in intra-species communication within biofilms, but also affect other species and the host. 26 P. aeruginosa lasR mutants exhibited metabolic shift, decreased oxygen consumption, and increased nitrate utilization, which are predicted to confer resistance to frequently used antibiotics tobramycin and ciprofloxacin, in response to the conditions in the CF lung. 32 Therefore, targeting QS in P. aeruginosa may attenuate virulence and increase susceptibility of bacteria in biofilms to antibiotics.
Antibiotic use and Economic Considerations in CF
Bacterial infections contribute significantly to the economic burden and costs associated with treatment and care for individuals. In the case of chronic infections such as in CF, they constitute a lifetime commitment to covering healthcare costs, particularly for antibiotics. In the United States, the average basic yearly medical cost per CF patient is $48,098; almost half of this cost is disbursed towards anti-Pseudomonal antibiotic tobramycin.15,43,44 This estimation does not include additional medical expenses incurred for transplantations, malnutrition, CF-associated diabetes, and other related complications. 43 Moreover, there has been a steady increase in the costs of prescription drugs, outpatient visits, durable medical equipment, and diagnostic procedures, with the highest cost incurred for treating and caring of CF patients older than 30 years of age. 44 Antibiotics must be administered not only during exacerbations but also for stable suppressive therapies so as to maintain low bacterial levels in the airways and to improve lung function. 45 An extrapolation of data from a study in a single Chicago hospital showed that total extra costs in all United States hospitals for treating resistant infections could rise to US$ 25–35 billion. 46 In the United Kingdom, 40% of P. aeruginosa isolates associated with CF infection were resistant to 2 or more antibiotics. 47 Under such circumstances, treatment strategy includes administering a combination of antibiotics with different modes of action, further increasing the cost.
The treatment for CF-related infections starts with oral and inhaled therapies in an outpatient setting, and the use of intravenous route for patients with severe exacerbations. The antibiotic course in general consists of at least two agents, with differing mechanisms of action against bacteria, chosen based on drug susceptibility testing. Sometimes, treatment involves multiple antibiotics to have a synergistic effect depending on the sensitivity pattern, patient tolerance, drug availability, and preference. 48 The fluoroquinolones, ciprofloxacin or levofloxacin, are the most commonly used oral agents to treat exacerbations caused by P. aeruginosa infection. Oral macrolide (azithromycin) treatment has been shown to improve lung function and other clinical markers through its anti-inflammatory effects, enhanced chloride efflux from airway epithelial cells, altered biofilm formation, and its direct antioxidant property.49–52 These agents are often combined with an inhaled course of tobramycin, aztreonam or colistin. Intravenous regimens include anti-Pseudomonal beta-lactams (pipercillin or ticarcillin), third-generation cephalosporins (ceftazidime), fourth-generation cephalosporins (cefepime), carbapenems (meropenem or imipenem), or monobactams (aztreonam), combined with an aminoglycoside (amikacin, gentamicin or tobramycin).53,54 During pulmonary exacerbations, high doses of gentamicin and tobramycin, usually at 10–15 mg/kg/day, are needed. Though the life expectancy of CF patients has improved with the use of antibiotics, risks remain in the form of nephrotoxicity, vestibular dysfunction, and loss of hearing from ototoxicity, especially with the use of intravenous aminoglycosides. 55 During pregnancy in CF patients, administration of tobramycin crosses placenta and accumulates in the amniotic fluid, fetal plasma, and in the kidneys. Though there is no evidence of congenital defects, damage to the VIII cranial nerve and nephrotoxicity can be expected.
The type and combination of antibiotics, dosage, and treatment schedules vary between countries, hospitals and centers, as well as between CF patients. For example, the 20 year old Copenhagen Model followed by the Copenhagen CF Center targeted early eradication of P. aeruginosa infections. The model consisted of inhaled colistin and intravenous ciprofloxacin for 3 months. In this regimen, 80% of CF patients were free of chronic P. aeruginosa infection for up to 15 years and the recovered bacterial isolates showed less resistance to those antibiotics.14,56–58 In-house standardized long term and low dose treatment regimens also proved effective against S. aureus, S. pneumoniae, and H. influenzae in Danish CF patients. 59 Such treatments are effective only in the case of early infection or colonization irrespective of the age of patients. It is nearly impossible to eradicate chronic colonization of P. aeruginosa in the form of biofilms. 56 However, antibiotics can keep the secondary infection status under control and can improve lung function, which is important for the day to day wellbeing of CF patients.
Co-Infections in CF
Besides P. aeruignosa, bacteria such as S. aureus, B. cepacia complex, and H. influenzae often infect CF patients. It is common in newborns in the early stages of CF and in older patients. CF airways also attract several atypical microbes (for example, Inquilinus limosus, Gluconobacter spp., Agrobacterium spp., Ochrobactrum spp.), which are of environmental origin.60–62 Though asymptomatic, atypical bacteria in the airways may cause inflammatory response in the absence of typical CF pathogens. 63 In the case of multispecies infections, it is often difficult to point signs and symptoms to a specific bacterium. Therefore, CF patients are required to receive an additional set of single or combination antibiotics, depending on the susceptibility information. Antibiotics used for S. aureus infection–-cephalexin, amoxicillin/clavulanic acid, dicloxacillin, clindamycin, doxycycline, minocycline, trimethoprim-sulfamethoxazole, linezolid, vancomycin, nafcillin, tigecycline, quinupristin-dalfopristin; for infections with B. cepacia complex–-ceftazidime, meropenem, trimethoprim-sulfamethoxazole, doxycycline, minocycline; for S. maltophilia infection–-fluoroquinolones, ceftazidime, ticarcillin-clavulanate, minocycline, trimethoprim-sulfamethoxazole; for Acinetobacter xylosoxidans – carbapenems, minocycline, pipercillintazobactam, ciprofloxacin; for O. anthropi and Brevundimonas diminuta infection–-imipenem and tobramycin; for infections with Nontuberculous Mycobacteria (Mycobacterium avium complex and M. abscessus)–-macrolides (azithromycin or clarithromycin), rifampin, ethambutol, amikacin, imipenem, tigecycline, linezolid, moxifloxacin.
The inventory of atypical species in the CF airways expanded with the increasing number of metagenomic studies, systematic sequencing of 16S ribosomal RNA, and other molecular signatures.41,42,60,64,65 Though the number of bacterial species identified from the CF airway samples increased, the interaction between members of CF microbiota and the subsequent pathophysiological outcome needs elucidation. Adding to existing complications, CF disease per se evolved into a multi-organ and multi-infection disease.
Conclusion
New forms of resistance in bacteria spread rapidly via intra-species and inter-species gene transfers in the environment, where community biofilms are common. The emergence and persistence of multidrug-resistant bacteria poses significant challenges to public health. Higher resistance rates correlate with a nation's higher per capita antibiotic consumption. According to the Center for Disease Control and Prevention, the USA has estimated that every year, around one-third of the prescriptions are unnecessary. 66 So far, there is no consensus on the best use of a single or combination antibiotic strategy to treat CF-related infections worldwide. It is also impractical to have a generalized therapeutic option to treat infections associated with CF, which is a highly heterogeneous disease condition. However, early eradication of bacterial infection/colonization may be a suitable option for efficient use of antibiotics in CF patients, achieving control over the development of antibiotic resistance in the future. On the humanitarian side, it would be appropriate to implement public health awareness measures against antibiotic resistant microbes. This could be achieved by continuously emphasizing prevention and fighting against the spread of emerging resistance through rational use of antibiotics and suitable infection control measures. Educating CF patients and their family members to maintain proper hygiene and protection may be an initial step towards avoiding unnecessary exposure to pathogens.
Author Contributions
Conceived and designed the concept/content: DS. Analyzed the data: DS. Wrote the first draft of the manuscript: DS. Contributed to the writing of the manuscript: DS. Agree with manuscript results and conclusions: DS. Jointly developed the structure and arguments for the paper: DS. Made critical revisions and approved final version: DS. All authors reviewed and approved of the final manuscript.
Funding
Author(s) disclose no funding sources.
Competing Interests
Author(s) disclose no potential conflicts of interest.
Disclosures and Ethics
As a requirement of publication the authors have provided signed confirmation of their compliance with ethical and legal obligations including but not limited to compliance with ICMJE authorship and competing interests guidelines, that the article is neither under consideration for publication nor published elsewhere, of their compliance with legal and ethical guidelines concerning human and animal research participants (if applicable), and that permission has been obtained for reproduction of any copyrighted material. This article was subject to blind, independent, expert peer review. The reviewers reported no competing interests. Provenance: the authors were invited to submit this paper.