
Abstract
In this review, we report on the organization of lipid molecules forming biomembranes. More precisely, the question is how these amphiphiles phase separate under a change of a suitable parameter, like temperature, pressure or membrane environment. The mixture may undergo a lateral or transversal phase separations. This essentially depends on two principal factors, which are the structure (length) of hydrocarbon chains of lipid molecules and the curvature asymmetry of membranes. When the former dominates, a lateral separation is then expected. In contrary, for those biomembranes of high curvature asymmetry, a vertical separation is rather observed. We examine the problem from a static (phase diagrams) and kinetics (relaxation in time) point of view. Finally, the discussion is also extended to the phase separation between lipid molecules and cholesterol in biomembranes (formation of rafts), between phospholipids and grafted polymers on liposomes, or between surfactant and its co-surfactant in bilayer systems.
Keywords
Inroduction
The biological membranes are of crucial importance to life, because they separate the cell from the outside environment. They also separate compartments inside the cell in order to protect important processes and specific events.
The conception that the biological membranes present as a lipid bilayer composed of two adjacent leaflets is now largely recognized by the scientific community. These leaflets are formed by amphiphile molecules possessing hydrophilic polar heads pointing out and hydrophobic fatty acyls chains forming the core.
The majority of lipid molecules are phospholipids [1–3]. These have a polar head group and two non-polar hydrocarbon tails, of which the length is of the order of 5 nm. The polar head is linked by glycerol to two fatty acid tails. One of these chains is a saturated straight fatty acid tail. While the other possesses a kink in the tail due to a cis double bound. Such a kink influences packing and lateral movement of lipid molecules. These lipid molecules then pack together into two leaflets in the membrane.
The temperature of the aqueous medium that surrounds the membrane may also have some effect on the packing of hydrocarbons. It has been observed that, at low temperatures, the bilayer is in a gel state (tightly packed). Indeed, in this case, the lateral diffusion is frozen out, and the solid structure is ensured by the attractive mutual interactions between hydrocarbon chains. At high temperatures, however, the bilayer is in the molten state (sol), and then, the lipid molecules effectuate free lateral movements and rotations. Of course, the sol-gel transition temperature depends on the chemical details of the bilayer.
The membrane incorporates another type of lipid that is cholesterol [1–3]. The fraction of cholesterol may vary with the membrane nature. For instance, plasma membranes essentially contain one cholesterol per phospholipid molecule. But other membranes as those protecting bacteria have no cholesterol. This lipid molecule has a steroid structure, and presents as a polar region linked to a non-polar hydrocarbon tail. Inside the membrane, the cholesterol molecules have the same orientation as phospholipids. The cholesterols have several functions in the membrane. For example, they give rigidity or stability to the cell membrane, and prevent crystallization of hydrocarbons.
The biomembranes also contain glycolipids (sugars). These are lipid molecules that microaggregate in the membrane, and may be protective and insulators. Certain kinds of molecules are bounded by sphingolipids as cholera and tetanus toxins. Sphingolipids and cholesterol favour the aggregation of proteins in microdomains called “rafts”. In fact, these play the role of platforms for the attachment of proteins as the membranes are moved around the cell and also during signal transduction. (More information about the rafts can be found in Refs. [4] and [5]).
The proteins (long macromolecules) are another component of the cell membranes. The transmembrane proteins are amphipathic and formed by hydrophobic and hydrophilic regions having the same orientation as the other lipid molecules. These proteins are also called “integral proteins” [6]. They have as function to transport substances across the membrane like ions and macromolecules. There exist other types of proteins that may be attached to the cytoplasm surface by fatty acyls chains or at the external cell surface by oligosaccharide. These are termed “peripheral membrane proteins”. They have many functions. Particularly, they protect the membrane surface, regulate cell signaling and participate in many other important cellular events [6]. In addition, some peripheral membrane proteins (those having basic residues), tend to bind electrostatically to negatively charged membranes, such as the inner leaflet of the plasma membrane [7].
The proteins possess the following architecture (see, for instance, the monograph by Alexei V. Finkelstein and Oleg B. Ptitsyn [8]) : a primary structure presenting as an amino acid sequence, a regular secondary structure (α-helix and one strand of β-structure), a tertiary structure of a globule folded by one chain, and finally a quaternary structure formed of an oligomeric protein by several chains (dimeric cro repressor).
To have an idea about the composition rate of various components forming the bilayer, we recall that the red cell membranes are composed of 19.5% (w/w) water, 39.6% proteins, 35.1% lipids, and 5.8% carbohydrates. More details concerning the structure, composition and function of the cell membranes can be found in the textbook by Y. Yawata [9].
The lipid molecules effectuate free movements on the two leaflets of the biomembrane. This is the so-called lateral diffusion [10]. The lipid molecules can also jump from a leaflet to the other. This vertical displacement is termed flip-flop diffusion. Moreover, a lipid molecule may rotate around its principal axis (rotational diffusion). The lateral and rotational diffusions give to the membrane its liquid characteristics, but with the constraint that the hydrophobic chains keep their parallelism and remain perpendicular to the surface of the bilayer. In fact, this order is similar to that usually encountered in the liquid-crystals context. As a conclusion, we can say that, from thermodynamics point of view, the stability of the biomembrane is largely determined by the nature of motion of its lipid molecules.
In most cases, the lipid molecules inside the bilayer are of different chemical structure. This means that they experience (relatively small) mutual segregation forces. As a consequence, under some conditions explained below, one assists to the appearance of domains formed by chemically incompatible species. This separation process has been observed within natural systems, such as cells and vesicles. Notice that the mechanism of the formation of domains is similar to that governing the traditional phase separation that takes place by varying temperature, ionic force, lateral pressure, and so on. The integration of a strange molecule in the bilayer may also lead to a rearrangement of lipid molecules into domains alternatively rich in unlike components.
Essentially, there exist two kinds of chemical incompatibilities. The lipid molecules can aggregate on each leaflet and form domains. This is the lateral separation transition (LST). The latter may also occur in the presence of ions (Ca+2, for instance) [11], around an incorporated macromolecule, or by a change of temperature [12]. The other possibility is that, the two leaflets of the bilayer may have different compositions, and then, the lipid molecules undergo a transversal separation transition (TST), where one species jumps from a leaflet towards the preferred one. The TST may also produce by an adsorption of a convenient polymer [12]. In the two cases, the separation mechanism is controlled by different interactions, namely the forces between the hydrophilic polar heads and between the hydrophobic tails. In fact, a variation of the external pH yields a change of the electric charge of the polar head. As a consequence, the attractive forces between two adjacent polar heads are reduced. The introduction of some ions can also lead to a redistribution of lipid molecules, due to the formation of complexes between polar heads and metallic ions. The adsorption of ionic polymers onto the two leaflets may increase the density of lipid molecules, which experience attractive mutual electrostatic interactions. A phase separation can also occur by varying the temperature of the bilayer composed of two lipid molecules of different melting temperatures.
To understand the organization of lipid molecules on the two leaflets, we first focus on a special phase separation that may produce under small variations of the bilayer structure. A typical example is provided by the phosphatidylcholine mixtures, when the acyls chains are of different lengths but of the same chemical nature. Their polar heads are identical but the length difference induces segregation forces between unlike chains. Second, the attention is paid to the influence of cholesterol and grafted polymers on the organization of lipid molecules within biomembranes. Finally, we extend the study to surfactant bilayers.
This paper is organized as follows. We discuss, in Sec. 2, the phase diagrams architecture, in space of relevant parameters, for both LST and TST. We extend, in Sec. 3, discussion to LST between phospholipids and cholesterols within biomembranes. In Sec. 4, we recall the essential of results relatively to LST between phospholipids and grafted polymer chains onto liposomes. Sec. 5 is devoted to kinetics of LST and TST. We draw some concluding remarks in the last section.
LST and TST Thermodynamics
Presentation of the system
To comprehend the thermodynamical mechanism of the phase separations within lipid membranes, we consider, as example, a bilayer composed of acyls chains A and B of different length. We shall denote by T, Φ1 and Φ2, δl, κ and δc
0
, the absolute temperature, the compositions of component A on the two leaflets, the length difference, the bending rigidity constant of the membrane, and the curvature asymmetry gap, respectively. If the mixture is assumed to be incompressible, then, the compositions of component B on the two sides of the bilayer are simply 1 - Φ1 and 1 - Φ2. Therefore, the phase separation between unlike lipid molecules is controlled by four kinds of relevant parameters (T, δl, κ, δc0). Notice that the dependence on temperature can be taken into account through the contact Flory interaction energies (per k
B
T unit) U11, U22 and U12 [13,14]. The former describes the effective interaction energy on leaflet 1, the second on leaflet 2, and the third between leaflets 1 and 2. For both LST and TST, U
ij
≫ 0. These conditions traduce the segregation phenomenon on each side of the bilayer. The temperature dependence of these energies was found to be [15]
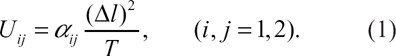
Here, the coefficients α11 ≫ 0, α22 ≫ 0 and α12 ≫ 0 depend on the chemical nature of the problem. More precisely, these quantities are directly proportional to the polarizabilities of the amphiphile molecules A and B. Therefore, the various interaction energies U ij 's vary as the inverse of the absolute temperature T. But if the latter is fixed to some value, the segregation interaction energies scale as the squared length difference. We note that the energies U ij 's can be experimentally accessible by a measurement of the interfacial tension between A and B-rich phases. Indeed, the effective interaction between unlike components is directly proportional to the interfacial tension [16]. Also, the mixing interaction, U, may be estimated by a light-scattering measurement of the structure factor. As a matter of fact, in the high-temperature regime, we have S(q → 0)∝U-1, where q is the wave-vector modulus.
Experimentally, the phase diagrams have been investigated by Knoll et al. [17] and by Silvius [18] by varying progressively the length of acyls chains. The considered mixtures were DLPC (C12)/DPPC (C16) and DMPC (C14)/DSCP (C18). The authors found that the separation transition produces when the difference between the lengths of acyls chains is about four groups CH2.
Theoretically, a new theory [15] was recently introduced, to describe, in a consistent way, the demixing transition within the lipid bilayer composed of acyls chains of different length. This theory is constructed from compositions (Φ1, Φ2) of the A-component on the two leaflets. Locally, these compositions fluctuate around the critical one Φ c = 1/2. With the help of the constructed free energy, which is a functional of compositions (Φ1, Φ2), the critical phase behaviour of the mixture has been studied in details in Ref. [15].
When the LST and TST are second order transitions, an exact equation for the critical line in the (δl, T)-plane has been derived [15], and along which the mixture phase separates. We do not recall here this equation, but we will extract from it all necessary information about the phase behaviour. The discussion will be made in terms of relevant parameters (T, δl, κ, δc0). Before, recall that the quantity δc0 = c A 0 − c B 0 ≫ 0 (asymmetry gap) represents the difference of spontaneous curvatures c A 0 and c B 0 of the membrane caused by the presence of species A and B. The fact that δc0 ≠ 0 reflects the asymmetry property of the bilayer. Return to the bending rigidity constant κ, and notice that it is of the order of few k B T. This quantity may be estimated measuring the membrane fluctuations amplitude. When the temperature T is fixed to some higher value (smaller values correspond to the gel-phase), the only remaining parameters for discussion are the length difference δl and the asymmetry gap δc0. Therefore, the phase separation between unlike acyls chains results from a competition between these two parameters.
As previously shown [15], the LST is entirely controlled by the length difference δl. More precisely, the separation transition occurs as long as the following condition
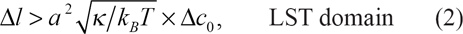
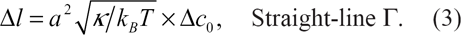

LST and TST domains. The straight-line Γ is drawn choosing κ = 25 x k B T as a particular value of the bending rigidity constant.
On the other hand, the critical line equation derived in Ref. [15] suggests that, when the asymmetry gap δc0 is small enough, the bilayer undergoes a LST for
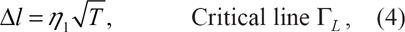
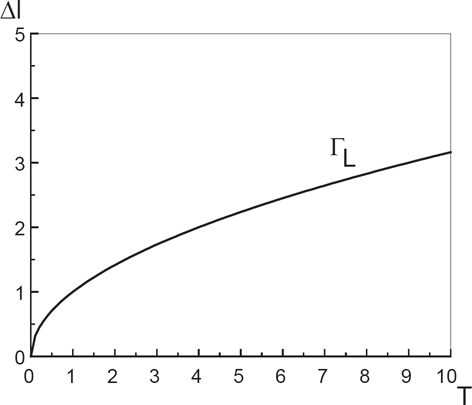
Critical line Γ L for LST, in the (T, δl) -plane.
The TST produces only for higher values of the asymmetry gap [15], that is
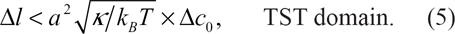
For the above numerical values of the bending rigidity constant and curvature difference, we have the condition that δl < 2a. The TST domain is indicated in Figure 1.
This vertical phase separation is then controlled by the asymmetry gap. From the general equation of the critical line derived in Ref. [15], we extract the information that the TST occurs for
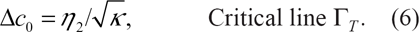
Finally, in Figure 3, we draw the critical line Γ T , which separates high and small asymmetry gap domains. In the upper domain, the mixture is separated, while in the bottom one, it is rather homogeneous.
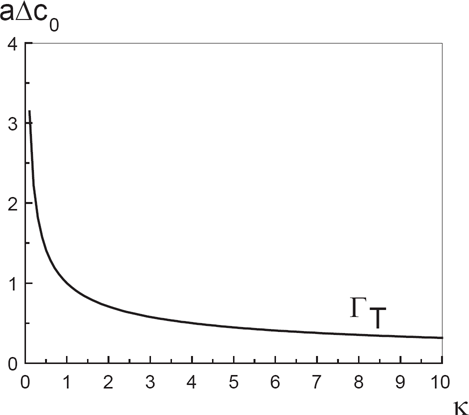
Critical line Γ T for TST, in the (κ, δc0)-plane. For this curve, the bending rigidity constant κ is expressed in k B T unit.
Beside the LST and TST between lipid molecules forming the bilayer, the biomembranes may also undergo another type of phase separation. This latter occurs between phospholipids and cholesterol, and it is accompanied by the appearance of “rafts” [19,20], which are liquid domains rich in cholesterol (mesophase domains). Also, these rafts may be domains composed of saturated lipids (sphingomyelin lipids), or particular proteins [21]. Such a phase separation was observed using fluorescence microscopy [22–28], and NMR and calorimetry [29,30]. We emphasize that this special phase separation has recently shown to have some relevance to signals transduction and membrane traffic [30].
Theoretically, the lipid-cholesterol phase separation has been extensively studied [31,32]. The first approach [31] was based on a one-order parameter model, which takes into account the couplings between the local composition and the internal membrane structure. For this theory, the order parameter is nothing else but the relative bilayer thickness. The purpose was a quantitative investigation of the first-order phase separation in binary mixtures of saturated/unsaturated lipids and lipid/cholesterol. In the same context, two years after, another theory [32] has been proposed. The new theory was constructed considering two order parameters and taking into account the couplings between the leaflets of the membrane. These order parameters are the distance of the cholesterol composition from its critical value and the relative increase of the thickness of the outer leaflet at the critical point [31,33]. Within the framework of this second theory, all phase diagrams related to the lipid-cholesterol LST have been drawn.
Despite the simplicity of the elaborated models [32,33], the theoretical predictions were found to be in agreement with experimental data. But the very recent theory introduced in Ref. [15] seems to be more consistent and coherent, and it can be regarded as a general scope for studying the phase separations within biomembranes. In fact, this theory is different from the models pointed out above by the following points: (1) the considered systems and the definitions of the order parameters are not the same, (2) the coexistence regions are macrodomains (instead of rafts or mesophase domains), (3) the new approach was constructed step by step and the parameters have explicit dependence on the mutual interactions between the unlike species, (4) the study was addressed not only to the lateral phase separation but also to the vertical one, (5) the phase diagrams have been investigated in all space of relevant parameters, and finally, (6) the membrane undulations were taken into account.
LST between Phospholipids and Grafted Polymer Chains
We start by noting that the majority of macromolecules forming the bilayer are simply anchored on the membrane surface, and form a soft branched polymer brush [34].
The extensive studies of grafted polymers were motivated by their potential applications in biological materials, such as liposomes [35–38]. These become now main tools in biology, biochemistry and medicine (as drugs transport agents). The liposomes are artificial vesicles that are synthesized from natural untoxic phospholipids and cholesterol, or from diblock-copolymer solutions.
Consider a membrane with grafted linear polymer chains by one extremity. The latter is a big lipid molecule called “anchor”. It is assumed that the phospholipids (species A) and anchors (species B) are chemically incompatible. This means that one is in the presence of a chemical segregation. It is also supposed that the grafted polymer chains effectuate free movement on the membrane surface. By a change of a convenient parameter like temperature, pressure or membrane environment, a phase separation may produce between the host lipid molecules and anchors.
Such a LST has been first studied in Ref. [34], by considering, for simplicity, that the substrate is a planar surface. The problem was re-examined in a very recent work [39]. The main difference is that, in the new work, the influence of the membrane undulations on the LST is taken into account. In particular, it has been shown that these undulations drastically affect the phase diagram shape of the considered lipid molecule mixture. More precisely, the segregation parameter is increased by a quantity χ
m
, under the effects of the membrane undulations. This extra contribution is the spatial integral of the induced attractive pair-potential between anchors, U(r). It was found [39,40] that the latter decreases with the distance as


This behaviour indicates that the effective attraction phenomenon between species of the same chemical nature is relevant only for those biomembranes of small bending rigidity constant. Also, notice that the fact that χ m is positive definite means that the membrane undulations have as effect to accentuate the phase separation between lipid molecules and grafted polymers. For example, if κ = 25 × k B T, the critical Flory parameter χ m is of the order of 0.2 (per k B T unit).
In the previous work [39], the temperature-composition phase diagram has been drawn, and the effects of the polydispersity of grafted polymer chains on the LST have been studied. As shown in this work, the polydispersity increases the segregation parameter. This can be understood as follows. In the presence of grafted polymer chains with different lengths, the excluded volume interactions are reduced in comparison to the monodisperse system. We recall that the LST is the consequence of a competition between the excluded volume forces between monomers belonging to grafted chains, the chemical segregation between phospholipids and anchors and the membrane undulations.
Consider now a bilayer composed of a surfactant (species A) and its co-surfactant (species B). In particular, the surfactant bilayer can undergo a TST due the strong asymmetry between the two leaflets. The approach pointed out in Sec. 2 can be extended to describe such a phase separation.
The surfactant bilayers have been the subject of extensive investigations in the past. The purpose was the study of their mechanical and geometrical properties due the curvature gap of the two components. For these bilayers, the variation of the free energy due to their deformations around some mean geometry is often described in terms of phenomenological constants that are the bending elastic modulus. In the context of systems made of a surfactant and its co-surfactant, much works have been devoted to the description of the curvature instability in the bilayer surface. The first investigation was achieved in Ref. [40], where the authors studied the consequences of the coupling between the composition asymmetry of the two leaflets and the curvature imposed to the membrane. On the other hand, the effects of electric charges in the case of ionic amphiphiles were also investigated [41–52]. In the same context, it has been proposed [53] a unified theory that takes into account the composition asymmetry and superficial charge density.
For instance, for the description of the elastic properties of the bilayer, a free energy is constructed. The latter incorporates contributions traducing the curvature effects and the composition asymmetry, and their coupling. This free energy is a functional of the mean curvature, c, and the composition of the surfactant, Φ. That of its co-surfactant is then 1 - Φ. This means that the surfactant/co-surfactant mixture is assumed to be incompressible. The minimization of this total free energy, with respect to the mean curvature c, leads to an effective one, c(Φ), which depends, of course, on composition Φ of surfactant. Substituting c by c(Φ) yields an effective free energy that is a pure function of composition Φ. Thanks to this effective free energy, the TST within the bilayer has been investigated, where the surfactant prefers to be located on the monolayer of positive curvature and the co-surfactant on the second one of negative curvature.
Notice that the studies were accomplished using a one-order parameter theory. To our opinion, a more realistic model should be rather a two-order parameter approach like the general one elaborated in Ref. [15]. Indeed, the surfactant may have different compositions on the outer and inner leaflets.
Kinetics of LST and TST
Consider a lipid membrane composed of two unlike lipid molecules A and B that laterally or transversally phase separate. To fix ideas, we assume that the phase separation is controlled by temperature. Hence, the separation transition occurs at some critical temperature T
c
depending on the mutual interactions between the two leaflets of the membrane and on other parameters of the problem (acyls chains length, curvature asymmetry, etc.). At equilibrium, the size of domains alternatively rich in species A and B coincides with the thermal correlation length
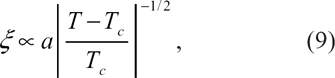
Assume now that the temperature of the system is lowered from an initial value T
i
towards a finale one T
f
very close to the critical point. Between initial and final equilibrium states, the membrane is out of equilibrium. Kinetics may be studied examining the variation of the overall order parameter, Φ (t) = ϕ (t) + Ψ(t), in time t. Here, ϕ (t) and Ψ(t) account for the composition fluctuations relative to the two leaflets. More precisely, the main question is how the quantity Φ relaxes between the two equilibrium states. Since the membrane is formed by unlike species, we expect that this relaxation is controlled by two kinds of relaxation times [54], τ1 and τ2. The former is small and can be interpreted as the time over which the system locally organizes. The second one, however, controls the separation transition, and can be regarded as the necessary time to form final domains of size ξ
f
. A scaling argument shows that the relaxation time τ2 scales as [54]

Within the framework of the linearized theory, the time variation of the order parameter writes [54]
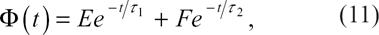
Now, if we take into account the spatial variation of the order parameter, the total composition fluctuation (in reciprocal space),
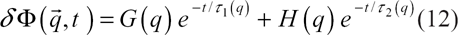
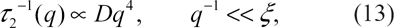
Notice that the above analysis may be extended, in a straightforward way, to kinetics of TST within surfactant bilayers.
Finally, we emphasize that the above kinetic properties may be experimentally accessible by a direct measurement of the dynamic scattering intensity.
In this paper, we reviewed the essential results concerning the organization of amphiphile molecules within lipid membranes. More precisely, we were interested in how these molecules phase separate under a change of a convenient parameter like temperature, pressure or membrane environment. Two phase separations are possible, namely lateral or vertical transitions. The nature of separation crucially depends on the mutual interactions between unlike molecules within the bilayer, and on other factors (acyls chains length, membrane asymmetry, etc.).
The majority of situations require the introduction of two-order parameter models, since the composition is not generally the same on the two leaflets of the membrane. This can be easily seen noting that the areas of interior and exterior membrane surfaces are not identical. In addition, the ionic environment is not the same near the two sides.
We reported on the phase behaviour related to the LST and TST, from static and kinetics point of view. As we pointed out, this phase behaviour is essentially controlled by the structure of amphiphile molecules (through the length of their hydrocarbon chains, for example) or by curvature asymmetry. When the structure dominate, the LST is then favoured. In contrary, a strong curvature asymmetry imposes to the system to transversally phase separate.
Also, we discussed the TST from bilayers formed by a surfactant and its surfactant. In this case, the phase separation is caused by a strong curvature asymmetry, where one species (surfactant or its co-surfactant) prefers to be located on the leaflet of positive curvature. As noted, such a system can be also modelled using a two-order parameter theory.
We examined another lateral separation phenomenon that occurs between phospholipids and cholesterol within biological membranes. This LST is generally accompanied by the appearance of rafts, which are small domains rich in cholesterol. These rafts protect the assembly of proteins on the membrane surface. This transition is largely studied from a theoretical and experimental point of view [55].
The discussion was also extended to the phase separation between phospholipids and grafted polymer chains onto a fluid membrane. It is known that the macromolecules anchored to the membrane phase separate by a variation of few 30% of the surface pressure, and form domains [56]. In very recent experimental investigations [57,58], a separation transition was also observed in lamellar phases of DNA-lipid complexes. More exactly, the authors reported on X-ray diffraction the structural studies of linear DNA chains that are adsorbed on cationic membranes in the lamellar phase of cationic lipid-DNA complexes. We recall that these biological materials are used in nonviral gene therapy clinical applications [59,60].
In this report, we have not discussed the effects of charges on the critical phase behaviour of the bilayer biomembranes. As shown in a very recent work [61], these charges have as effect to renormalize (additively) the elastic constants of the membrane. Therefore, the above conclusions remain unchanged, but the presence of charges simply shifts the critical line along which the system undergoes a phase transition.
Notice that, as it should be, the particular structure of the bilayer biomembranes (chemincal nature of acyls chains and polar heads, inclusions, etc.) affects the value of the critical parameters. But the critical exponents, such as those defining the order parameter, the thermal correlation length and the correlation function remain unchanged. These are independent on the microscopic details of the problem.
To our opinion, the theory presented above can not be applicable to plant cells that often contain only rather small amounts of phospholipids and cholesterol. In fact, the separation transition occurs only for those macroscopic bilayers that incorporate many phospholipids (thermodynamic limit).
Finally, we note that the quality of solvent (selective, good or theta-solvent) where the lipid membrane is trapped has certainly some relevance for physics of the phase separation. In addition, other complexities may also affect this phase transition, such as the polydispersity of the anchored macromolecules, the presence of charges, and so on.
Footnotes
Disclosure
The author reports no conflicts of interest.