
Abstract
Background
Because of the potential anti-inflammatory effects, linagliptin, a therapeutic dipeptidyl peptidase-4 inhibitor, is used as an effective drug for diabetic patients for whom inflammation is a prognosis-related factor. We investigated the anti-inflammatory mechanism of linagliptin using seven markers.
Methods
We pretreated human umbilical vein endothelial cells (HUVECs), with linagliptin and lipopolysaccharide (LPS). The cytosolic fractions were evaluated for protein kinase A (PKA), protein kinase B (PKB), protein kinase C (PKC), ratio of reactive oxygen species (ROS) and Cu/Zn superoxide dismutase (SOD), activator protein 1 (AP-1), and adenosine 3′,5′-cyclic monophosphate (cAMP).
Results
Linagliptin increased the PKA and PKC activities and the cAMP levels in LPS-treated cells. However, it inhibited LPS-induced PKB phosphorylation, ratio of ROS and Cu/Zn SOD, and LPS-stimulated AP-1 nuclear translocation.
Conclusion
We reaffirmed the anti-inflammatory and antioxidant effects of linagliptin. These effects might be related to the three protein kinases. Our findings suggest that linagliptin has a wide range of anti-inflammatory effects.
Keywords
Introduction
The number of patients with diabetes is increasing world-wide. 1 Dipeptidyl peptidase-4 (DPP-4) inhibitors reduce blood glucose levels by preventing the DPP-4 enzymes. 2 Although nine DPP-4 inhibitors have been known, their antidiabetic effects are alike. Therefore, it is significant to recognize the fundamental pharmacological mechanism of each inhibitor in order to be used in suitable patients. Linagliptin is the bile-excreted, oral antidiabetic DPP-4 inhibitor. Linagliptin therapy has been shown to reduce the risk of cardiovascular and cerebrovascular disorders.3,4 Moreover, we have reported its anti-inflammatory effects both in vivo and in vitro.5,6 Some studies have also examined the anti-inflammatory effects of linagliptin on endothelial cells in mice. 7 Therefore, linagliptin may have specific effects other than decreasing blood glucose levels.
In our previous in vitro research, linagliptin inhibited lipopolysaccharide (LPS)-stimulated interleukin (IL)-6 production, intranuclear p65 [a nuclear factor kappa-B] expression, and p38 mitogen-activated protein kinase (MAPK) activation in human umbilical vein endothelial cells (HUVECs). 6 These three markers have been found to be important in LPS-and protein kinase A (PKA)-mediated inflammatory signaling pathways. Moreover, these three markers are the most downstream target proteins of inflammatory signaling pathways. However, due to certain limitations in our research, the detailed cascade reactions and upstream or downstream of the markers were not investigated. Further extensive research on the anti-inflammatory mechanisms of linagliptin might clarify the specificity of linagliptin, which the other nine DPP-4 inhibitors do not have. Protein kinases and oxidative stress are the key elements in inflammatory signaling pathways. Therefore, studying the anti-inflammatory effects of linagliptin on protein kinases and oxidative stress might help elucidate its underlying mechanism.
This study is an extension and a more thorough follow-up of our previous research. In this study, we investigated the cascade reactions upstream and downstream to intranuclear p65 expression or p38 MAPK activation. The following seven markers were analyzed: PKA, whose activation may prevent MAPK phosphorylation; 8 protein kinase B (PKB), which is also known as Akt and is a serine/threonine-specific protein kinase that plays a key role in multiple cellular processes; 9 protein kinase C (PKC), which regulates various cellular responses including inflammation, by maturation, catalytic activation, and targeting; 10 reactive oxygen species (ROS), which are used as markers of oxidative stress; Cu/Zn superoxide dismutase (SOD), which is one of the major antioxidant enzymes; activator protein 1 (AP-1), which is a transcription factor that regulates gene expression in response to a variety of stimuli including bacterial infections; 11 and adenosine 3′,5′-cyclic monophosphate (cAMP), which activates PKA. 12
Methods
Materials and Cell Culture
Linagliptin compounds were supplied by Boehringer Ingelheim Pharmaceuticals, Inc. Sitagliptin was provided by Santa Cruz Biotechnology, Inc. LPS was obtained from Sigma-Aldrich. All purest compounds used in this research were available commercially. The HUVEC, EGM™ BulletKit™ medium, and EGM™ SingleQuots™ Kit were obtained from Lonza. HUVECs were cultured in basal medium supplemented with 10 mL fetal bovine serum (FBS), 2 mL bovine brain extracts, 0.5 mL human epidermal growth factor, 0.5 mL hydrocortisone, 50 mg/mL gentamicin, 50 µg/mL amphotericin B, and 0.5 mL ascorbic acid (EGM™ BulletKit™ medium and EGM™ SingleQuots™ Kit). The cells were cultured at 37°C in an atmosphere of 5% CO2 and 95% air.
Drug treatment
All operations were performed under sterile conditions in medium without FBS, bovine brain extracts, and hydrocortisone in order not to elevate the spectrophotometric absorbance through binding with coated antibodies.
HUVECs were cultured for 24 hours at a concentration of 1–2 x 105 cells/mL, and were pretreated with 50 nM linagliptin, 150 nM sitagliptin for 1 hour, or left untreated. After this, the culture media were changed, and the cells were treated with 1 µg/mL LPS together with 50 nM linagliptin, 150 nM sitagliptin, or left untreated. The concentration of LPS was 1 µg/mL, which was sufficient to induce inflammation in HUVECs. For all experiments with LPS, the above procedure was performed using 50 nM linagliptin or 150 nM sitagliptin. The same number of cells (N = 1–2 x 105 cells/mL) was used in all treatment groups for standardization. Each experiment was repeated three times.
Measurement of PKA and PKC activities
All samples from the cytosolic fractions were measured using ELISA kits 30 minutes after treatment with LPS and linagliptin. We examined the PKA and PKC activities using the PKA and PKC activity kits, respectively (Enzo Life Sciences, Inc.). In this assay, microplates precoated with PKA and PKC substrates were used. The assayed samples were added to the suitable wells, followed by the annexing of adenosine triphosphate to start the reaction. Then, a phosphospecific substrate antibody was annexed to the wells, which binds to the phosphorylated peptide substrate. The peroxidase-conjugated secondary antibody was then bonded to the phosphospecific antibody. The tetramethylbenzidine (TMB) substrate developed the assay and the intensity of the color obtained was proportional to the PKA and PKC phosphotransferase activities. The acid stop solution stopped the color development reaction, and the color intensity was measured with a microplate reader at 450 nm (SpectraMax).
Measurement of PKB phosphorylation
All samples from the cytosolic fractions were measured using ELISA kits 30 minutes after treatment with LPS and linagliptin. We measured PKB phosphorylation using the RayBio Phosphorylation Akt (Ser473) and pan-Akt ELISA kit (RayBiotech, Inc.). This sandwich ELISA kit is an in vitro enzyme-linked immunosorbent assay for the measurement of phospho-Akt (Ser473) and pan-Akt. A pan-Akt antibody was coated onto a microplate. Samples were pipetted into the wells and Akt present in the sample was bound to the wells by the immobilized antibody. The wells were washed, and anti-phospho-Akt (Ser473) or anti-pan-Akt were used to detect phosphorylated or pan-Akt, respectively. After washing away the unbound antibody, a horseradish peroxidase (HRP)-conjugated anti-rabbit IgG was pipetted into the wells. The wells were washed again, a TMB substrate solution was added to the wells, following which a color developed in proportion to the amount of Akt (Ser473) or pan-Akt bound. The stop solution changed the color from blue to yellow, and the color intensity was measured at 450 nm (SpectraMax).
Evaluation of ROS
The 5-(and-6)-chloromethyl-2′,7′-dichlorodihydrofluorescein diacetate (CM-H2DCF-DA; Molecular Probes) was used to evaluate intracellular ROS formation in HUVECs. The samples from the cytosolic fractions were measured 10 minutes after treatment with LPS and linagliptin. After cellular uptake, CM-H2DCF-DA was first modified to 2′,7′-dichlorofluorescein by intracellular esterases and then to oxidized fluorescent 2′,7′-dichlorofluorescein by intracellular ROS. The CM-H2DCF-DA was dissolved in dimethyl sulfoxide, and the cells were incubated with 7 µM CM-H2DCF-DA for 15 minutes at 37°C and 5% CO2. After treatment, the cells were washed with heated phosphate-buffered saline. Fluorescence intensity was measured on a plate reader (Berthold Technologies) at an excitation wavelength of 488 nm and a measuring wavelength of 525 nm.
Measurement of Cu/Zn SOD levels
All samples from the cytosolic fractions were measured using ELISA kits 10 minutes after treatment with LPS and linagliptin. We measured Cu/Zn SOD levels using Human Cu/Zn SOD Platinum ELISA BMS222/BMS222TEN (eBioscience, Inc.). This ELISA system shows no cross-reactivity with other SOD isozymes. An anti-human Cu/Zn SOD was adsorbed onto microwells. Cu/Zn SOD in the sample or standard was bound to antibodies adsorbed to the microwells. An HRP-conjugated anti-Cu/Zn SOD antibody was added, which binds to Cu/Zn SOD captured by the primary antibody. Following incubation, the unbound HRP-conjugated anti-Cu/Zn SOD antibody was removed by a washing step, and a substrate solution with HRP was annexed to the wells. The color intensity was proportional to the amount of Cu/Zn SOD present in the sample or standard. The acid stop solution stopped the color development reaction, and the color intensity was measured with a microplate reader at 450 nm (SpectraMax).
Measurement of AP-1 nuclear translocation
A P-1 nuclear translocation was evaluated by determining the ratio of intranuclear and intracellular AP-1 levels. Samples were measured using ELISA kits 30 minutes after treatment with LPS and linagliptin. We measured AP-1 levels using the AP-1 ELISA kit (Affymetrix). An oligonucleotide containing an AP-1 consensus-binding site was immobilized on the microplate. Activated AP-1 from nuclear or cytosolic fractions specifically binds to this oligonucleotide. The complex bound to the oligonucleotide was detected by an antibody directed against the AP-1 subunit. An additional secondary HRP-conjugated antibody provided sensitive colorimetric readout, easily quantified by spectrophotometry. The absorbance was measured at 450 nm (SpectraMax).
Measurement of cAMP
All samples from the cytosolic fractions were measured using ELISA kits 10 minutes after simultaneous treatment with LPS and linagliptin. We examined the cAMP levels by using a cAMP EIA kit (Cayman Chemical). This assay is based on the competition between free cAMP and a cAMP-acetylcholinesterase (AChE) conjugate (cAMP Tracer) for a limited number of cAMP-specific rabbit antibody binding sites. Because the concentration of the cAMP Tracer is held constant while the concentration of cAMP varies, the amount of cAMP Tracer that is able to bind to the rabbit antibody is inversely proportional to the concentration of cAMP in the well. This rabbit antibody-cAMP (either free or tracer) complex binds to the mouse monoclonal anti-rabbit IgG that was previously attached to the well. The plate was washed to remove any unbound reagents, following which Ellman's Reagent (which contains the substrate to AChE) was added to the well. The product of this enzymatic reaction had a distinct yellow color, and the color intensity was measured with a microplate reader at 412 nm (SpectraMax).
Statistical analysis
The results are expressed as mean ± standard error. The results of various treatments were compared with the control by using one-way analysis of variance and Bonferroni post hoc analysis. A one-sided P-value of less than 0.05 was considered statistically significant.
Results
PKA activity
Linagliptin alone significantly increased PKA activity compared with the control treatment. Further, there was no difference in PKA activity between the LPS-treated cells and the untreated control cells. However, linagliptin significantly increased PKA activity in the LPS-treated cells (Fig. 1).
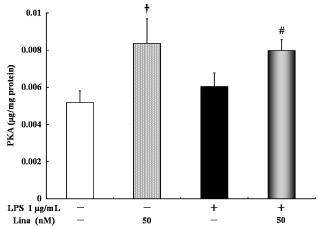
Effects of linagliptin on protein kinase A (PKA). HUVECs were treated with LPS and/or linagliptin. Linagliptin increased the PKA activity in both LPS-treated and untreated cells. PKA in the cytosolic fractions was determined by ELISA after 30 minutes of treatment. Values are expressed as mean ± standard error. †P < 0.05 vs. control; #P < 0.05 vs. LPS 1 µg/ml.
Phosphorylation of PKB
With respect to PKB phosphorylation, no significant difference was detected between the linagliptin-treated and untreated control cells, and between the LPS-treated and untreated cells. However, linagliptin significantly inhibited the LPS-stimulated PKB phosphorylation (Fig. 2).
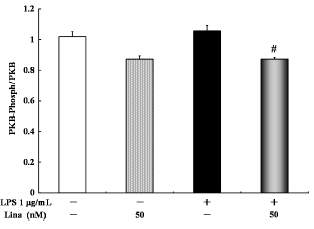
Effects of linagliptin on protein kinase B (PKB). HUVECs were treated with LPS and/or linagliptin. Linagliptin inhibited the LPS-stimulated PKB phosphorylation. PKB in the cytosolic fractions was determined by ELISA after 30 minutes of treatment. Values are expressed as mean ± standard error. #P < 0.05 vs. LPS 1 µg/mL.
PKC activity
Linagliptin alone significantly increased PKC activity compared with the control treatment. Further, there was no significant difference in PKC activity between the LPS-treated cells and the untreated control cells. However, linagliptin significantly increased PKC activity in the LPS-treated cells (Fig. 3).
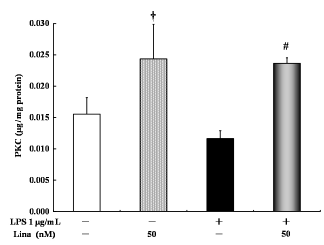
Effects of linagliptin on protein kinase C (PKC). HUVECs were treated with LPS and/or linagliptin. Linagliptin increased the PKC activity in both LPS-treated and untreated cells. PKC in the cytosolic fractions was determined by ELISA after 30 minutes of treatment. Values are expressed as mean ± standard error. †P < 0.05 vs. control; #P < 0.05 vs. LPS 1 µg/mL.
Effects of linagliptin on oxidative stress state
Oxidative stress is controlled by a balance between the ROS-generating system and the antioxidant system. Cu/Zn SOD is one of the typical cytosol markers in the antioxidant system. Therefore, we evaluated oxidative stress by determining the ratio of ROS and Cu/Zn SOD.
With respect to the ratio of ROS and Cu/Zn SOD, no significant difference was detected between the linagliptin-treated and untreated cells. However, LPS only significantly increased the ratio of ROS and Cu/Zn SOD in comparison to the control treatment. Linagliptin significantly decreased the LPS-stimulated ratio of ROS and Cu/Zn SOD (Fig. 4).
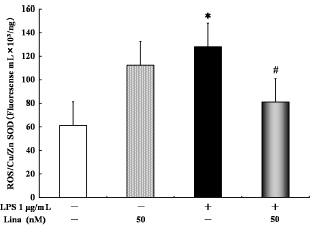
Effects of linagliptin on oxidative stress state. HUVECs were treated with LPS and/or linagliptin. LPS alone significantly increased the ratio of reactive oxygen species (ROS) and Cu/Zn superoxide dismutase (SOD) compared with the control treatment. Treatment with linagliptin significantly decreased the LPS-stimulated ratio of ROS and Cu/Zn SOD. ROS and Cu/Zn SOD in the cytosolic fractions were determined by ELISA after 10 minutes of treatment. Values are expressed as mean ± standard error. *P < 0.05 vs. control; #P < 0.05 vs. LPS 1 µg/ml.
AP-1 nuclear translocation
With respect to AP-1 nuclear translocation, no significant difference was detected between the linagliptin-treated and untreated control cells, and between the LPS-treated and untreated cells. However, linagliptin significantly inhibited the LPS-stimulated AP-1 nuclear translocation (Fig. 5).
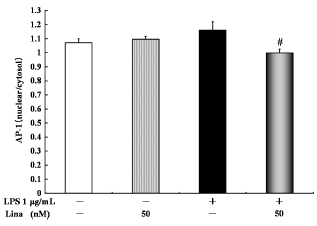
Effects of linagliptin on activator protein 1 (AP-1) nuclear translocation. HUVECs were treated with LPS and/or linagliptin. Linagliptin inhibited the LPS-stimulated ratio of intranuclear and intracellular AP-1 levels. intranuclear and intracellular AP-1 in the cytosolic fractions were determined by ELISA after 30 minutes of treatment. Values are expressed as mean ± standard error. #P < 0.05 vs. LPS 1 µg/mL.
cAMP levels
With respect to the cAMP levels, no significant difference was detected between the linagliptin-treated and untreated cells. LPS alone significantly decreased cAMP levels in comparison to the control treatment. However, linagliptin significantly increased cAMP levels in the LPS-treated cells (Fig. 6).
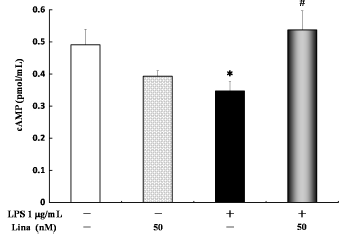
Effects of linagliptin on adenosine 3′,5′-cyclic monophosphate (cAMP). HUVECs were treated with LPS and/or linagliptin. LPS significantly decreased the cAMP levels compared with the control treatment. Treatment with linagliptin significantly increased cAMP levels in the LPS-treated cells. cAMP in the cytosolic fractions was determined by ELISA after 10 minutes of treatment. Values are expressed as mean + standard error. *P < 0.05 vs. control; #P < 0.05 vs. LPS 1 µg/mL.
PKA activity with linagliptin and sitagliptin treatment
Linagliptin alone significantly increased PKA activity compared with the control treatment. However, sitagliptin alone did not significantly increase PKA activity compared with the control treatment. Further, linagliptin significantly increased PKA activity in the LPS-treated cells. However, sitagliptin did not significantly increase PKA activity in the LPS-treated cells (Fig. 7).
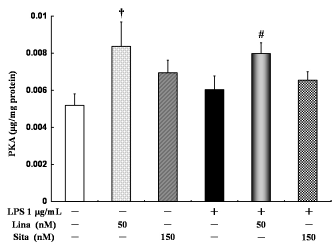
Effects of linagliptin and sitagliptin on protein kinase A (PKA). HUVECs were treated with LPS and/or linagliptin and/or sitagliptin. Linagliptin increased the PKA activity in both LPS-treated and untreated cells. Sinagliptin did not increase the PKA activity in both LPS-treated and untreated cells. PKA in the cytosolic fractions was determined by ELISA after 30 minutes of treatment. Values are expressed as mean ± standard error. †P < 0.05 vs. control; #P < 0.05 vs. LPS 1 µg/mL.
Discussion
In this study, linagliptin increased the PKA and PKC activities in both LPS-treated and untreated cells and increased cAMP levels in the LPS-treated cells. On the other hand, linagliptin inhibited the LPS-induced PKB phosphorylation, oxidative stress, and AP-1 nuclear translocation. To our knowledge, this study is the first to examine the effectiveness of linagliptin on three types of protein kinases in vitro using HUVECs. Moreover, by examining the AP-1 nuclear translocation and oxidative stress, we reaffirmed the anti-inflammatory and antioxidant effects of linagliptin.
We found that pretreatment with linagliptin significantly increased the PKA activity by 1.6-fold in comparison to the control. Among all DPP-4 inhibitors, linagliptin is the only DPP-4 inhibitor that has a xanthine-related skeletal system. This system, which is also exhibited by caffeine, is related to anti-inflammation. 13 These drugs promote cAMP levels via inhibiting phosphodiesterase, which in turn induces PKA activation.8,12 In this study, linagliptin significantly increased cAMP levels in the LPS-treated cells. Therefore, there is a possibility that linagliptin controls MAPK phosphorylation through PKA activation by increased cAMP levels. In fact, we previously reported the opposite effects of H89 (PK A inhibitor) on linagliptin-induced anti-inflammatory effects. H89 significantly increased linagliptin-inhibited IL-6 levels. 6 However, the pharmacological mechanisms on the anti-inflammation mediated by xanthine-related skeleton system remain unclear, necessitating further study.
PKB, PKC, and AP-1 are the key proteins, which mediate the LPS-induced inflammatory signaling pathway. PKB, a serine/threonine kinase, is recruited to the membrane by the activation of phosphatidylinositol 3-kinase and phosphatidylinositol (3,4,5)-trisphosphate. Activated PKB mediates inflammatory responses. 9 AP-1 is a transcription factor, which regulates gene expression in response to a variety of stimuli, including bacterial infections. 11 In this study, linagliptin inhibited LPS-induced PKB phosphorylation and AP-1 nuclear translocation. Therefore, we could reaffirm the anti-inflammatory effects of linagliptin. PKC is also one of the serine/threonine kinases, which phosphorylates the substrate and plays an important role in inflammatory signaling pathways. 10 Dai et al 14 reported that two DPP-4 inhibitors, sitagliptin and vildagliptin, inhibited the signals of inflammation via inhibiting phosphorylated PKC, 24 hours after administration of medicine. Therefore, we expected linagliptin to inhibit the LPS-induced PKC activity. However, on the contrary, linagliptin promoted the PKC activity in both LPS-treated and untreated cells. These data suggested that PKC might not be involved in the anti-inflammatory mechanism of linagliptin or LPS of proinflammatory agent might be affected. It is necessary to investigate the induction of inflammation by other agents. Further investigations on the inhibitory effects of linagliptin are ongoing.
Some reports have examined the antioxidative effects of linagliptin using ROS generation. 15 ROS, which is strongly involved in inflammation, induces P38 phosphorylation via the autophosphorylated apoptosis signal-regulating kinase 1. 16 On the other hand, Cu/Zn SOD is one of the typical cytosol proteins in the antioxidant system. Therefore, we thought that the ratio of ROS and Cu/Zn SOD represented the oxidative stress state. Because linagliptin significantly decreased the LPS-stimulated ratio of ROS and Cu/Zn SOD, linagliptin was thought to prevent oxidative stress. Oxidative stress is an important atherosclerosis-related factor. Linagliptin therapy also reduces the risk of cardiovascular and cerebrovascular diseases. 3 Therefore, the decreased ratio of ROS and Cu/Zn SOD might reflect an anti-atherosclerotic effect of linagliptin treatment.4,7
This study had some limitations. First, the anti-inflammatory effects of linagliptin in cascade reactions are still unclear. We assume the direct various anti-inflammatory points by linagliptin: membrane receptors, DPP-4 (CD26), intracellular inflammation signal proteins. However, the results of this study suggested that linagliptin may have wide-ranging anti-inflammatory effects. Second, we did not investigate the morphological changes of cells. However, the obvious morphological changes of the HUVECs were not observed during the subculture, it is certainly important to reveal morphological changes of cells using cell death staining such as TUNEL assay. Third, linagliptin activated PKA and inhibited cAMP in the absence of LPS. LPS of proinflammatory agent might be affected. Moreover, it is unclear that these effects are specific in HUVECs. It is important to investigate the anti-inflammatory effects of linagliptin in other cell lines. Further extensive study on the anti-inflammation of DPP-4 inhibitors may be able to use nine DPP-4 inhibitors according to patient requirements.
Conclusion
We reaffirmed the anti-inflammation and antioxidant effects by linagliptin in vitro by using HUVECs pretreated with LPS. Moreover, in this study, we investigated the effects of linagliptin on the three protein kinases. Our findings suggested that linagliptin has wide-ranging anti-inflammatory effects.
Author Contributions
Conceived and designed the experiments: YN and MI. Analyzed the data: YN. Wrote the first draft of the manuscript: YN. Contributed to the writing of the manuscript: MI. Agreed with manuscript results and conclusions: YN, MI, MT, HH, IO, HG, and YG. Jointly developed the structure and arguments for the paper: TG, KH, AY, and TK. Made critical revisions and approved the final version: YK. All the authors reviewed and conceded the final version of the manuscript.