
Abstract
In contrast to radiolabelled erythrocytes and platelets, radiolabelled neutrophils leave the circulating blood in an exponential manner, indicating random rather than age-dependent removal. Neutrophils transit the spleen with a range of residence times that are log normally distributed. We hypothesized that neutrophils are conditioned to undergo apoptosis to an extent that depends on their intrasplenic residence time and that this provides an explanation for the random removal of these cells from blood. Splenic venous and peripheral arterial blood was sampled simultaneously during abdominal surgery in four patients and age-dependent apoptosis assessed in whole blood using annexin V/PI staining. Apoptosis increased after 4 and 20 h ex-vivo incubation and was invariably higher in splenic venous vs arterial neutrophils. Transit through the spleen appears to promote neutrophil apoptosis, with subsequent high efficiency clearance by the liver. This may explain the mechanism underlying the random removal of neutrophils from the blood.
Introduction
In-111-labelling studies have shown that human neutrophils are destroyed in the liver, spleen and bone marrow in approximately equal numbers. 1 Given that the spleen has a blood flow of about 200 ml/min, 2 and the mean neutrophil life span in the circulation is about 10 h,3,4 there must be a permanent arterio-venous neutrophil concentration gradient across the spleen of approximately 3%. 5
Data from mice suggest that circulating neutrophils begin to re-express CXCR4 in an age-dependent manner, facilitating their return to the bone marrow through interaction with CXCL12. 6 This would be consistent with age-related destruction of neutrophils in the bone marrow. Labelled neutrophils however, leave the circulation exponentially with a half-time of about 7 h, irrespective of labelling technique,1,3,4 suggesting that, in addition to age-related removal, there is also a major element of random destruction.
Neutrophils undergo pooling in the spleen with a mean transit time of ∼10 min. 7 The kinetics of neutrophil trafficking through the spleen are consistent with a log-normal distribution of individual intra-splenic neutrophil transit times, 7 hence some neutrophils transit the spleen within seconds whereas others take 20 min or longer. We hypothesized that the longer a neutrophil resides in the spleen, the more likely it is to be permanently retained in this organ and destroyed by apoptosis. We also hypothesized that neutrophils leaving the spleen (via splenic venous blood) will, having been exposed to intra-splenic pro-apoptotic stimuli, undergo apoptosis more rapidly than neutrophils with a history of shorter or less recent exposure to the splenic micro-environment.
The aim of this proof-of-concept study was to compare the rates of apoptosis of neutrophils harvested from splenic venous blood in comparison with neutrophils sampled simultaneously from arterial blood.
Methods
Patients
Seven patients undergoing Whipple's procedure for carcinoma of the pancreas consented to intra-operative sampling of splenic venous and radial arterial blood. Of these, four went on to be operable (see Table 1 for clinical details). The study was approved by the local research ethical committee and each patient gave written informed consent.
Clinical and laboratory details of the patients studied.
Measurement of Neutrophil Apoptosis in Whole blood
Paired samples of blood (5 ml) were taken from the radial arterial line and the splenic vein at the time of surgery using 1 ml citrate used as anticoagulant. These samples were handled in an identical manner throughout. The blood was divided into 100 μl samples and incubated in multi-well plates containing penicillin and streptomycin (final concentration 50 U/ml) in a 5% CO2 incubator at 37°C. Either immediately or following a 4 or 24 h period of incubation, the cells were centrifuged (150
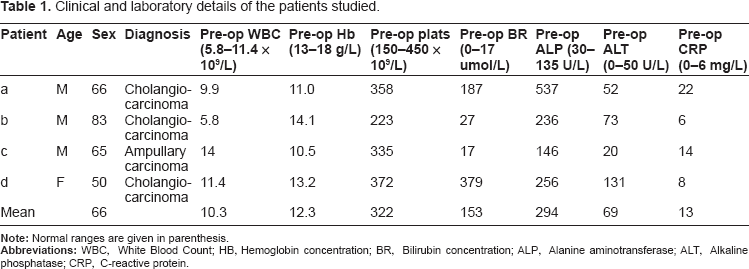
Assessment of neutrophil apoptosis in whole blood samples. Representative flow cytometric histograms (upper panels) and dot plot images (lower panels) of whole blood samples anti-coagulated with citrate and then incubated in vitro for 4 h (A, C) or 20 h (B, D) prior to annexin V/PI staining, fixation, red cell lysis and flow cytometric analysis. Analysis of the gated neutrophil population is shown only and demonstrates the clear separation of apoptotic and non-apoptotic cells, with minimal cell necrosis (PI positivity) and the time-dependent nature of constitutive apoptosis. Data are derived from a single experiment representative of >100 replicates.
Results
There was minimal annexin V or PI positivity at time zero in either splenic venous or arterial blood neutrophils however apoptosis was very apparent at 4 and 20 hours. In all patients, the rate of apoptosis was greater in neutrophils in the splenic venous blood samples compared with peripheral blood neutrophils (Fig. 2). This was most marked in patients a, c and d at 4 h and patient b at 20 h.
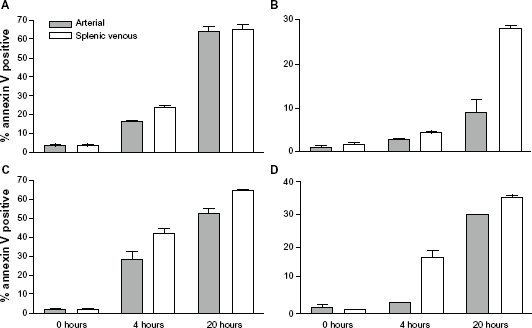
Comparison of rates of apoptosis in neutrophils in splenic venous and arterial blood samples. Whole blood samples were taken from splenic venous and peripheral arterial blood in four patients (shown independently A-D), anti-coagulated with citrate and then incubated for 0, 4 or 20 h prior to annexin V and PI staining, red cell lysis and flow cytometric gating and analysis. The percentage of Annexin V high (apoptotic) cells is displayed as a function of ex vivo incubation time in the 4 patients studied. Neutrophils from splenic venous blood showed greater Annexin V binding than neutrophils from peripheral arterial blood. Each column represents the mean (SD) of triplicate measurements.
While the rate of neutrophil apoptosis observed at 20 hours in patients a and c would be in keeping with many previous in vitro studies undertaken using purified human neutrophils, 8 patients b and d showed much lower rate of apoptosis. While we suspect that this may relate to different exposures to circulating survival factors and/or differences in the priming status of cells between patients, such data were not collected. However, there was no difference in the extent of basal shape-change of neutrophils in the arterial and splenic venous samples (data not shown); ie, no evidence that the spleen independently affected the priming status of circulating neutrophils.
Discussion
An attraction of the technique described here for quantifying neutrophil apoptosis in whole blood is that it lends itself to ex vivo work and near-patient testing. It was evaluated extensively prior to use in this patient group and demands only very small blood samples, an advantage for patients undergoing major surgery. Likewise because many samples can be analyzed immediately and without lengthy purification steps, they can be taken from multiple sites and processed simultaneously for direct comparative studies.
This study has shown reproducible differences in the biological properties of neutrophils isolated from blood entering and leaving the spleen and suggests that the splenic microenvironment may alter the apoptotic threshold of neutrophils. Individual neutrophil residence time in the spleen is log-normally (ie, exponentially) distributed, 7 so if the likelihood of apoptosis being triggered depends on residence time, our results would be consistent with the hypothesis that the spleen converts the age-dependent cell death that is observed in vitro 10 to the random destruction seen in vivo. The rarity with which apoptotic neutrophils are seen in peripheral blood could then be explained by the strategic location of the liver, in relation to the spleen, for entrapment and engulfment of apoptotic or apoptotic-prone neutrophils escaping from the spleen. Indeed the liver is well recognized as the predominant site of destruction of damaged neutrophils, and liver disease is one of the few pathological states where apoptotic neutrophils can be seen in the circulation. 11
All circulating blood cells are delayed as they transit the spleen. Red cells have the fastest transit time, an essential requirement as the spleen would otherwise become grossly congested. In spite of their markedly disparate sizes, granulocytes and platelets have similar mean spleen transit times of ∼10 min.2,7,12 It is likely that lymphocytes and monocytes have a similar transit time. 12 Why these blood cells have such long transit times is unknown, as are the mechanisms controlling them. A role in initiating neutrophil apoptosis would be the first function attributable to a long mean intrasplenic transit time. If the spleen's microenvironment is conducive to apoptosis, this may also explain why cancer metastases are so rare in this organ, in contrast to organs in which neutrophil transit is delayed, such as the liver 7 and bone marrow, 1 or modestly delayed, as in the lungs. 13
Peripheral blood neutrophils from patients with cirrhosis also undergo apoptosis in vitro more rapidly than neutrophils from age-matched healthy subjects. 14 Our study suggests three possible inter-related mechanisms to explain this: firstly, failure of a diseased liver to extract apoptotic neutrophils arriving directly from the spleen; secondly, splenic venous blood by-passing the liver via porto-systemic shunts; and thirdly, increased spleen-induced apoptosis as a result of more frequent re-circulations of neutrophils through an enlarged spleen. All these support the concept of the spleen promoting neutrophil apoptosis. Moreover, in a study of ‘hepato-splenic’ neutropenia in patients with schistosomiasis, an accelerated rate of in vitro neutrophil apoptosis was observed in patients with a spleen but not in patients who had undergone splenectomy. 15
In conclusion, these preliminary data suggest a novel role for the spleen in the control of circulating neutrophil lifespan and suggest that further experiments in this field will be productive.
Disclosure
This manuscript has been read and approved by all authors. This paper is unique and is not under consideration by any other publication and has not been published elsewhere. The authors and peer reviewers of this paper report no conflicts of interest. The authors confirm that they have permission to reproduce any copyrighted material.
Footnotes
Acknowledgements
This work was supported by the MRC, Wellcome Trust, Papworth Hospital NHS Foundation Trust R&D Department and NIHR Cambridge Biomedical Research Centre. JFW was funded by a MRC Clinical Training Fellowship and CS by a Wellcome Trust Clinical Training Fellowship.