
Abstract
The study of death receptor family induced apoptosis has gained momentum in recent years with the knowledge that therapeutic antibodies targeting DR4 and DR5 (death receptor's 4 and 5) have proved efficacious in multiple clinical trials. The therapeutic rationale is based on targeting and amplifying a tumour tissues normal cell death programme (apoptosis). While advances in the targeting of DR4 and DR5 have been successful the search for an agonistic antibody to another family member, the Fas receptor, has proven more elusive. This is partly due to the differing
Keywords
Introduction
Extrinsic apoptosis is mediated through the activation of the TNF/death receptor family. The death receptors such as DR4 and DR5 (death receptor's 4 and 5) are a group of receptors with a conserved cytoplasmic region responsible for induction of the caspase cascade which results in cell death.
1
Molecules that directly activate these receptors, such as agonistic monoclonal antibodies to the Fas or TRAIL receptors and recombinant Fas or TRAIL ligands, represent a new class of therapeutic modalities.
1
There are several agonistic monoclonal antibodies against TRAIL which have recently proved efficacious for cancer treatment in multiple clinical trials.
2
While advances in the targeting of TRAIL have been successful, the search for agonistic antibodies to the Fas receptor has proven more elusive. This is partly due to its expression pattern but also due to the differing
Fas (CD95/Apo-1) is a member of the TNF cell surface receptor family, normally involved in the down regulation of activated lymphocytes by triggering apoptosis (programmed cell death).
3
Binding of Fas Ligand (FasL) or agonistic monoclonal anti-Fas antibodies (anti-Fas mAbs) causes trimerisation of the Fas receptor and leads to the recruitment of adaptor protein FADD (Fas-associated death domain). This in turn recruits procaspase 8 (FADD-like IL-1-converting enzyme, FLICE) to form the death-inducing signalling complex (DISC).3–5 Procaspase 8 molecules become activated at the DISC and in turn activate pro-apoptotic downstream molecules such as caspase 3 and bcl-2 family member BID.
4
The design of therapeutics which target Fas-induced apoptosis is an exciting area of cancer research as deficiency of this cell death programme is a major cause of tumour progression.3,7,8 The search for targeted therapeutics is made more challenging by the fact that individual molecules give contrasting effects
Currently, there are several methods available to assess antibody induced apoptosis
To overcome the shortcomings of current
Materials and Methods
Cell lines and normal culture conditions
Both Jurkat (a human leukaemia T cell line) and HCT116 (a human colon cancer cell line) were obtained from ATCC (American Type Culture Collection, Manassas, VA, USA). Cells were maintained in RPMI Medium 1640 (Sigma, St. Louis, MO, USA) and McCoy's 5A Medium respectively with 10% fetal bovine serum (Invitrogen, Grand Island, New York, USA) and grown in an incubator at 37 °C with 5% CO2. All tissue culture plates and other plasticware were purchased from Sarstedt (Rommelsdorfer Straße, Nümbrecht, Germany).
Reagents
ProSep-G was purchased from Millpore (Billerica, MA, USA). Anti-human Fas antibody (clone CH11) was obtained from Upstate Biotechnology (Lake Placid, NY, USA). Anti-human Fas antibody (clone DX2), human IgG Isotype control and Annexin-V-APC were purchased from BD Biosciences Pharmingen (Franklin Lakes, NJ, USA). Anti-human Fas antibody (clone IB2) was developed in-house. Prodidium Iodide-PE (PI-PE) and Thiazolyl Blue Tetrazolium Bromide (MTT) were purchased from Sigma (St. Louis, MO, USA).
Induction of apoptosis in jurkat/HCT116 cells with anti-Fas antibody
One ml of 1.25 × 105 cells/well was seeded in a 24-well plate the night before the treatment. Cells were treated with CH11/DX2/1B2/Isotype control at a final concentration of 1 μg/ml for 18 hours in the 37 °C with 5% CO2 incubator before the cells were subjected to MTT assay (HCT116 cells only) or Annexin-V and PI staining.
Induction of apoptosis in Jurkat/HCT116 Cells with Prosep-G cross-linked anti-Fas antibody
100 ng of ProSep-G was coated onto a 24-well plate and incubated at 37 °C for 2 hours. CH11/DX2/1B2/ Isotype control at a final concentration of 1μg/ml were added to the ProSep-G coated 24-well plate to allow cross-linking at 37 °C for 2 hours with 100 RPM shaking. The ProSep-G coated 24-well plate with cross-linked anti-Fas antibody was then washed once with 200 μl of PBS, and 1 ml of 2.5 × 105 cells/ well was seeded onto the plates. Plates were incubated for 18 hours at 37 °C with 5% CO2 prior to MTT assay (HCT116 cells only) or Annexin-V and PI staining.
Annexin-v and PI staining
For suspension cells, 1 ml of Jurkat cell suspensions was transferred to a 15 ml tube. For adherent cells, supernatant (medium and floating HCT116 cells) were transferred to 15 ml tubes. The rest of the adherent cells were detached with PBS-EDTA, Dulbecco's phosphate buffered saline (Invitrogen, Grand Island, New York, USA) containing 1mMEDTA.
Both Jurkat and HCT116 cells were pelleted by centrifuged at 1,300 RPM for 3 minutes, and washed two times with 2 ml Flow Cell Wash (BD Biosciences, San Jose, CA, USA). Cell pellets were then re-suspended in l00 μl x 1 Annexin-V Binding buffer (BD Biosciences Pharmingen, Franklin Lakes, NJ, USA) and 4 μl Annexin-V-APC plus 2 μl PI-PE dye mix was added. Samples were incubated in the dark for 15 minutes with 100 RPM shaking at room temperature and analysed within the next half hour by flow cytometry. Tests were done in triplicate, counting a minimum of 5000 total cells each.
MTT Assay
For adherent cells after 18 hours, the diluted MTT was added to the wells as 200ul/well. Two hours later l00 μl of DMSO (Sigma, St. Louis, MO, USA) were added to each well. The ProSep-G coated 24-well plate with cross-linked anti-Fas antibody was then incubated at room temperature with gentle shaking for 5 minutes to dissolve reduced MTT crystals. Absorbance values were determined by ELISA Tecan Sunrise reader (PerkinElmer, Waltham, Massachusetts, USA) at wavelength 570 nm.
Results and Discussion
In this study, we combined a novel ProSep-G coated 24-well plate anti-Fas mAb cross-link technique with traditional Annexin-V and PI staining methods. This combined methodology provides a new platform for high throughput functional screening of anti-Fas mAbs in multiple cell types. The results suggest that this cross-linking technique will provide researchers with a robust way of characterising new anti-Fas mAb candidates that are potentially capable of inducing
In Figure 1 and 2 (results outlined in Table 1), we compare the traditional Annexin-V and PI staining methods with our combined method (cross-linking the anti-Fas mAb) for their ability to identify functional anti-Fas mAbs. No apoptosis was observed in the untreated or Isotype control treated populations (negative control; Fig. 1A). Figure 1B and C show that, in the absence of cross-linking, the anti-human Fas antibodies clone IB2 and clone DX2 were only able to induce low level apoptosis on the Jurkat cells after 18 hours treatment. Only 4.7% Annexin-V positive (early apoptosis) and 9.2% Annexin-V and PI staining positive (apoptotic cell) were observed for DX2; and 2.5% Annexin-V positive (early apoptosis) and 5.1% Annexin-V and PI staining positive (apoptotic cell) were observed for IB2 (Table 1). Analysis of another well characterised anti-human Fas antibody, clone CH11, treatment without the cross-linking (Fig. 1D), gave 7.8% Annexin-V positive staining and 10.3% Annexin-V and PI double positive staining (Table 1). The ability of CH11 to induce apoptosis in the absence of artificial cross linking is likely due to the fact that it is an IgM isotype capable of multiple receptor binding and automatic receptor trimerisation. DX2 and IB2 are both IgG1 isotype and therefore require cross-linking of two or more antibodies to induce receptor trimerisation. 15
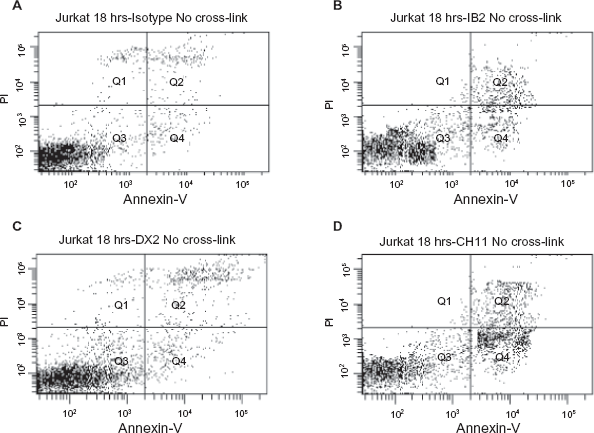
Flow cytometry analysis of Jurkat cells following 18 hour incubation with
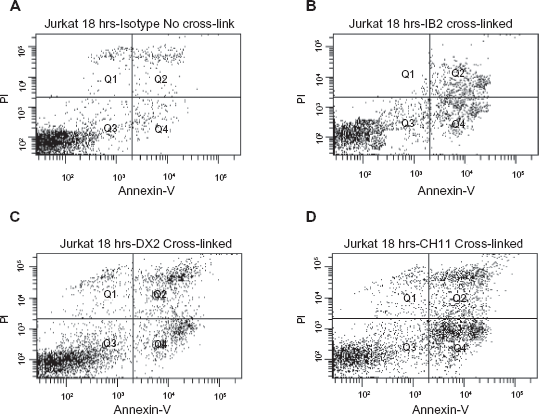
Flow cytometry analysis traditional Annexin-V (X-axis) and PI (Y-axis) staining by using the novel mAb cross-link technique with the method of Jurkat cells following 18 hour incubation with
Comparison the apoptotic effects in Jurkat or HCT116 tumour cells exposed to different anti-Fas mAbs (with and without ProSep-G cross-link technique) or Isotype controls.
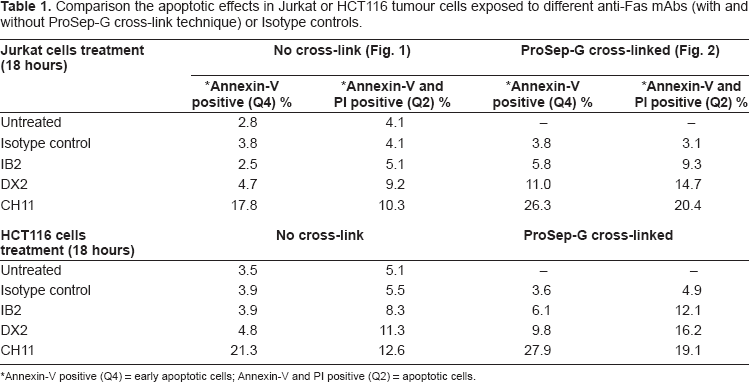
Annexin-V positive (Q4) = early apoptotic cells; Annexin-V and PI positive (Q2) = apoptotic cells.
By using our novel ProSep-G coated anti-Fas mAb cross-linking technique, DX2, IB2 and CH11 were able to induce much higher level of apoptosis in Jurkat and HCT116 cells. For the cross-linked DX2 the level of apoptosis rose from 4.7% to 11% Annexin-V positive staining and from 9.2% to 14.7% for Annexin-V and PI double positive staining in Jurkat cells (Fig. 2C; Table 1). In the same set of experiments, the cross-linked IB2 level of apoptosis increased from 2.5% to 5.8% for Annexin-V positive staining and from 5.1% to 9.3% for Annexin-V and PI double positive staining (Fig. 2B; Table 1). For the CH11, the cross-linking technique also improved its apoptosis level. Cross-linked CH11 apoptosis level increased from 17.8% to 26.3% for Annexin-V positive staining and from 10.3% to 20.4% for Annexin-V and PI double positive staining in Jurkat cells (Fig. 2D; Table 1). In contrast, the level of apoptosis observed in the cross-linked Isotype control remained the same (Fig. 2A; Table 1). We observed a similar pattern of results when we further investigated Isotype control, DX2 and CH11 induced apoptosis at 14- and 24-hour time points (data not shown). As for the adherent colorectal cancer HCT116 cell line, the same pattern of Annexin-V and PI staining results was observed with all antibodies (Table 1).
The novel anti-Fas mAb cross-link technique has a greater ability to differentiate between non-functional and functional antibodies than conventional methodologies. MTT assays were also performed to observe the level of cell kill by each anti-Fas mAb in the presence or absence of cross-linking. HCT116 cells were incubated with anti-Fas antibodies DX2, IB2 and CH11 for 18 hours prior to analysis. All three antibodies gave modest cell killing which was not statistically significant. When the HCT116 cells were incubated with the cross-linked anti-Fas antibodies DX2, CH11 or IB2 for 18 hours, the level of cell death did increase significantly (Fig. 3). The ProSep-G cross-linked anti-Fas antibodies DX2, CH11 and IB2 showed enhanced cell killing by 35%, 50% and 15% respectively compared with those antibodies incubated with the cells alone.
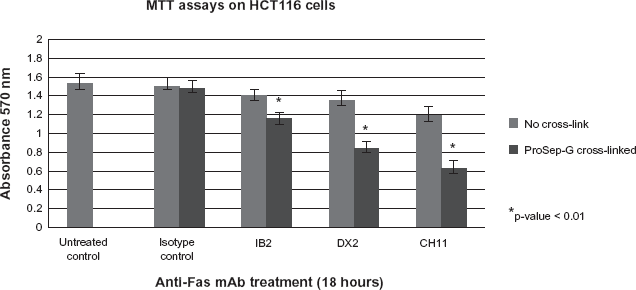
MTT assays showed the ProSep-G cross-linked anti-human Fas antibodies IB2, DX2 and CH11 had significantly enhanced cell killing compared to IB2, DX2 or CH11 incubated with the cells alone.
These results suggest that our novel ProSep-G coated 24-well plate anti-Fas mAb cross-link technique with traditional Annexin-V and PI staining method is well suited to high throughput analysis of anti-Fas mAbs that may be able to induce apoptosis
Conclusion
In conclusion, our novel ProSep-G coated 24-well plate anti-Fas mAb cross-linking technique combined with traditional Annexin-V and PI staining method provides a useful tool for the functional characterisation of anti-FAS mAbs. The use of a technique that artificially mimics
Disclosures
The authors report no conflicts of interest.